
Abstract
Aim:
To establish a system for monitoring the activation of G-protein-coupled receptors (GPCRs) using β-lactamase enzyme fragment complementation (EFC) technology.
Methods:
Two inactive β-lactamase deletion fragments, bla(a) and bla(b), were fused to β-arrestin and GPCR, respectively. A stable cell line named HEK/293-β2a2, which expressed two fusion proteins, GPCR/bla(b) and β-arrestin2/bla(a), was generated under antibiotic selection. A natural compound library of high performance liquid chromatography (HPLC)-fractionated samples from the ethanol extracts of Chinese medicinal herbs was used for high-throughput screening (HTS) of β2-adrenoceptor (β2AR) agonists against the cell line HEK/293-β2a2. The interested hits were validated by the measurement of second-messenger cyclic adenosine monophosphate (cAMP) production.
Results:
The stable cell line HEK/293-β2a2 responded to β2AR agonist/antagonist in a dose-dependent manner. The EC50 value obtained for isoproterenol was 15.5 nmol/L, and the IC50 value obtained for propranolol was 51.9 nmol/L. Furthermore, HTS was performed to identify β2AR agonists from the natural compound library we established. The Z′ factor value was determined to be 0.68. Three hits were identified from primary screening and found to be as potent as isoproterenol in a cAMP assay.
Conclusion:
A cell-based high-throughput functional assay was established to directly monitor the activation of GPCRs based on the interaction between agonist-activated GPCR and β-arrestin using β-lactamase EFC technology, which can be used to search for leads in the natural compound library.
Similar content being viewed by others


Introduction
G-protein-coupled receptors (GPCRs) are the largest known family of transmembrane receptors. They bind to and transduce signals for a huge variety of endogenous ligands, and they are involved in most physiological functions, such as metabolism, neuronal transmission, embryonic development, hormonal homeostasis and cancer invasion1, 2. Of all currently marketed drugs, over 30% target GPCRs3, and the development of new technologies for GPCR-based drug discovery promises to keep this field active for years to come.
Recently, several cell-based drug discovery strategies were developed using β-arrestin as a surrogate marker for GPCR function. β-arrestins are ubiquitously expressed in all cells and tissues. They bind with high affinity to the agonist-activated GPCR carboxyl-terminal tail. Assays developed based on GPCR-β-arrestin interaction, such as transfluor assay4, 5, bioluminescence resonance energy transfer assay6, PathHunter™ β-arrestin Assays7 and Tango assay,8 were widely used in the investigation of many GPCRs9, 10, 11, 12, 13, 14, 15. These achievements have demonstrated that monitoring GPCR-β-arrestin interaction can offer a direct indication of GPCR activation.
Here we describe a cell-based protein-protein interaction assay based on β-lactamase complementation technology that uses GPCR and β-arrestin as protein partners to monitor GPCR activation. The β-lactamase-based enzyme fragment complementation (EFC) assay was pioneered in 200216, 17. The properties of β-lactamase provide significant advantages in mammalian cells. β-lactamase is monomeric and relatively small18, and it can be expressed in many different cell types and diverse cellular compartments19. In addition, there is no endogenous β-lactamase activity. A highly sensitive cell-permeable fluorescent substrate, CCF4/AM, also offers great potential for the quantification of results in intact cells20. In our research, we successfully applied this system to monitor GPCRs activation. Two inactive β-lactamase deletion fragments, bla(a) and bla(b), were fused to β-arrestin and GPCR, respectively. The two β-lactamase fragments complement each other and recover activity by agonist-promoted interaction between GPCR and β-arrestin. The recovery of β-lactamase activity was then determined with the cell-permeable fluorescent substrate, CCF4/AM, to demonstrate GPCR activation.
In this study, we chose β2-adrenoceptor (β2AR) as a model. β2AR is a typical GPCR that has been thoroughly characterized. The β2-adrenoceptor agonists are the most widely used agents in the treatment of asthma owing to their bronchodilator actions21. Chinese medicinal herbs have been used to treat many diseases, and they have demonstrated effectiveness for thousands of years. Our lab established a natural compound library containing high performance liquid chromatography (HPLC)-fractionated samples from the ethanol extracts of Chinese medicinal herbs. This library was successfully used to find inhibitors for Severe Acute Respiratory Syndrome-associated coronavirus (SARS-CoV) and agonists for dopamine receptors22, 23. In the following, we describe in detail the application of β-lactamase-based EFC assay to monitor GPCR activation and to screen for agonists of the β2-adrenoceptor from the natural compound library we constructed.
Materials and methods
Materials
Isoproterenol and propranolol were purchased from Sigma (St Louis, USA). CCF4/AM was obtained from Invitrogen (Carlsbad, CA, USA).
Plasmid construction
β-lactamase gene fragments bla(a) and bla(b) were amplified and ligased to pEGFP-N1 vector (Clontech, Mountain View, CA, USA). The primers used were the forward primers [5′-CGG GGTACCGGAGGAGGAGGAAGTGGAGGAGGAGGAAGTGGAGGAGGAGGAAGTCACCCAGAAACGCTGGTG-3′ for bla(a) and 5′-CCCAAGCTTGGTGGTGGTGGTAGTGGTGGTGGTGGTAGTGGTGGTGGTGGTAGTCTACTTACTCTAGCTTCC-3′ for bla(b)] and the reverse primers [5′-ATTTGCGGCCGC TCAGCCAGTTAATAGTTTG-3′ for bla(a) and 5′-ATTTGCGGCCGCTTACCAATGCTTAATCAGT-3′ for bla(b)]. A flexible (G4S)3 linker was inserted in front of each β-lactamase fragment. Human β-arrestin2 and β2AR cDNA without a stop codon were cloned in front of the β-lactamase fragments bla(a) and bla(b), respectively, to generate pβarr2-bla(a) and pβ2AR-bla(b), which are two expression vectors. The primers used were the forward primers (5′-CTAGCTAGCACCATGGGGGAGAAACCCGGGAC-3′ for β-arrestin2 and 5′-CTAGCTAGCACCATGGGGCAACCCGGGAAC-3′ for β2AR) and the reverse primers (5′-CGGGGTACCCTAGCAGAGTTGATCATC-3′ for β-arrestin2 and 5′-CCCAAGCTTCAGCAGTGAGTCATTTGTAC-3′ for β2AR).
Cell culture and stable cell line selection
HEK-293 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum and 2 mmol/L L-glutamine. The stable cell line HEK/293-β2a2 was generated by co-transfection of two expression constructs, pβarr2-bla(a) and pβ2AR-bla(b), into HEK-293 cells. Cells were selected by the addition of 0.8 g/L G418 into the culture medium for 15 d. The isolation of HEK/293-β2a2 cell clones was performed by the stimulation of cellular clones with 1 μmol/L isoproterenol. The clones, which turned from green to blue under fluorescence microscopy detection, were individually isolated.
CCF4/AM-based β-lactamase assay
To perform the CCF4/AM assay, cells were loaded with 1 μmol/L CCF4/AM according to the dye provider's protocol and then incubated at room temperature for 30 min. A Flexstation II (Molecular Devices, Sunnyvale, CA, USA) was used to determine the blue/green ratio by setting the excitation at 409 nm and the emission at 460 nm (for blue fluorescence) or 530 nm (for green fluorescence). The blue/green ratio was calculated following the manufacturer's instructions.
Induction/inhibition of β2AR receptors
HEK/293-β2a2 cells (4×104 cells/well) were seeded in a 96-well black matrigel-coated microplate with a clear-bottom and cultured for 24 h. For agonist dose response, cells were then incubated with isoproterenol in serum-free DMEM for 5 min at 37 °C. For antagonist dose response, cells were first pre-incubated for 10 min in serum-free DMEM containing different concentrations of propranolol at room temperature. Then 1 μmol/L of isoproterenol was added and incubated for another 5 min at 37 °C. CCF4/AM was then loaded to perform the β-lactamase assay. The blue/green ratio was determined by a Flexstation II. Images were captured by fluorescence microscopy (Olympus, Tokyo, Japan).
Preparation of the natural compound library
Herbal plants with therapeutic indications were collected from an herb market in Anguo, Hebei, China. Briefly, the dry Chinese herbs were dissolved in 90% ethanol for 12 h and sonicated for 0.5 h. The remains were then soaked with 50% ethanol for 12 h and extracted for 0.5 h. The ethanol extract was filtered and lyophilized. Products after lyophilization were re-dissolved in 90% ethanol for high performance liquid chromatography (HPLC) isolation. A total of 16 gradients were obtained by HPLC isolation for each Chinese herb extract. All gradients were lyophilized, dissolved in dimethyl sulfoxide (DMSO) and kept in a -80 °C freezer.
High-throughput screening
Stably transfected HEK/293-β2a2 cells were seeded at 4×104 cells per well in a 96-well black matrigel-coated microplate with a clear-bottom. The cells were cultured for 24 h before treatment. Culture media were replaced with serum-free DMEM containing 0.01 g/L diluted testing compounds. Positive and negative controls were designed in each plate. After incubation at 37 °C for 5 min, β-lactamase activity was quantitated with CCF4/AM.
cAMP assay
HEK/293-β2a2 cells were plated (4×104 cells/well) in a 96-well plate for 24 h. Compounds were diluted with phosphate-buffered saline and incubated with the cells for 15 min prior to measuring the cAMP level with cAMP-Glo™ Assay (Promega, Madison, WI, USA), according to product instructions. Luminescence was determined using an Envision 2100 (PerkinElmer, Waltham, MA, USA).
Data analysis and calculations
Data were presented as mean±SD. Statistical significance was evaluated with Student's t-test, and the difference was considered statistically significant at P<0.05. The Z' factor was calculated as described previously by Zhang et al24. For concentration-response determinations, a curve was fitted to calculate EC50 and IC50 using GraphPad Prism 5 software.
Results
Recombinant cell line generation
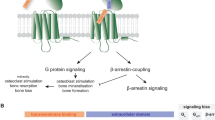
To study GPCR activation with a β-lactamase EFC assay, stable cell line HEK/293-β2a2 was established by the co-transfection of plasmids encoding human β-arrestin2/bla(a) and β2AR/bla(b), two fusion constructs (Figure 1A), into HEK-293. They were selected as described in materials and methods. The cell clone with the best signal to background ratio was chosen and named HEK/293-β2a2. In this clone, β2AR/bla(b) and β-arrestin2/bla(a), two fusion constructs, were highly expressed. The two β-lactamase deletion mutants, bla(a) and bla(b), are individually inactive and have a low affinity for each other. Ligand stimulation caused the conformation change of β2AR and recruited β-arrestins to the cell membrane. The interaction between β2AR and β-arrestin brought the two inactive β-lactamase fragments together to reconstitute a functional enzyme. The recovery of β-lactamase activity, as determined with the CCF4/AM assay, was a direct indicator of agonist-induced GPCR activation via the β-arrestin recruitment (Figure 1B).

β-lactamase EFC based receptor/β-arrestin interaction assay. (A) Vector construction. β-lactamase fragment was fused to target protein with a flexible (G4S)3 linker. (B) Illustration of β-lactamase EFC based receptor/β-arrestin interaction assay. Agonist (A) activated GPCR recruits β-arrestin to the cell membrane, which brings two β-lactamase fragments proximity to reconstitute and recover its activity. The recovery of β-lactamase activity was then determined with CCF4/AM assay.
Characteristics of the stable cell line HEK/293-β2a2
Agonist stimulation in cells co-expressing β2AR/bla(b) and β-arrestin2/bla(a) caused the complementation of two β-lactamase fragments. The reconstituted β-lactamase hydrolyzed the substrate CCF4/AM and emitted blue fluorescence when excited at 409 nm. The generation of blue fluorescent cells could be attributed to the activation of G-protein-coupled receptors (Figure 2A, top). The agonist effect could be inhibited by the β2AR antagonist, propranolol. When pre-incubated with propranolol, the percentage of cells that emitted blue fluorescence after agonist stimulation was similar to untreated cells (Figure 2A, middle and bottom). Dose response curves for isoproterenol and propranolol were also calculated using GraphPad Prism 5 software. As shown in Figure 2B, HEK/293-β2a2 cells responded to the known β2AR agonist, isoproterenol, with a dose-dependent increase in β-lactamase activity. The EC50 value of isoproterenol was found to be 15.5 nmol/L, which is not only similar to previously reported data for HEK-293 cells (11 nmol/L) using an isoproterenol-stimulated β2AR sequestration assay25 but also data obtained in C2C12 cells (27 nmol/L) using a β-galactosidase based EFC assay7. Dose-dependent inhibition was demonstrated under the treatment of the β2AR antagonist, propranolol, in the presence of 1 μmol/L isoproterenol (Figure 2C). The IC50 of 51.9 nmol/L obtained for propanolol using β-lactamase EFC assay was also consistent with reported data26.

Characteristics of stable cell line HEK/293-β2a2. (A) Immunofluorescence assay of β-lactamase activity. Stable cell line HEK/293-β2a2 treated with 1 μmol/L isoproterenol (top); pre-incubated with 10 μmol/L propranolol and then treated with 1 μmol/L isoproterenol (middle); untreated (bottom) shown clear difference in blue cells percentage imaged by fluorescence microscope (magnification×100). (B) and (C) Dose response curves for isoproterenol and propranolol. Data represent means±SD for triplicate samples.
Screening of β2AR agonists from natural compound library
To see if this cell-based assay was suitable for high-throughput screening, the Z' factor value was calculated as described previously by Zhang et al24. Half of the plate was treated with 1 μmol/L isoproterenol, and half of the plate was untreated. Four wells (A1, B1, G12, and H12) were designed for reagent control to calculate the blue/green ratio for the demonstration of β-lactamase activity. The Z' factor was 0.68 in our experiment (Figure 3A). To further assess the value of this cell model for searching agonists targeting β2-adrenoceptor, we screened 352 HPLC-fractionated samples from the ethanol extracts of Chinese medicinal herbs in a 96-well format. Isoproterenol was used as the positive control, and 1% DMSO was used as the negative control. Cells were incubated with fractionated samples for 5 min in a serum-free medium. All wells were treated identically, and all contained 1% DMSO, a concentration which has been determined to have no adverse effect on the cells or the assay (data not shown). The fraction activity was expressed as the percentage of maximal β-lactamase induction by 1 μmol/L isoproterenol. As shown in Figure 3B, three fractions each exhibited a significant effect on the induction of β-lactamase activity (>40%). The effect of the other fractions on the induction of β-lactamase complementation is also shown in Figure 3. These fractions, which had activities below three times of standard deviation, were considered insignificant.

Z′ factor determination and high-throughput screening. (A) Z′ factor determination. This experiment was representative of three experiments. (B) High-throughput screening from natural compound library. Each triangle represents the reading from one well. The induction of β-lactamase activity was expressed as a percentage of isoproterenol induction (performed in the same plate).
Hits validation
To confirm the activity of the primary screen hits, we repeated the receptor/β-arrestin interaction assay in triplicate (Figure 4A). Fractions 10-B11 and 10-C11 were identified to have >50% of maximum induction while fraction 10-D11 showed >40% of maximum induction. Three positive hits were further validated with the melanocortin receptor 4 (MC4R) cell line, which was also generated based on the β-lactamase complementation assay. As expected, they all had no agonist effect on the MC4R cell line (Figure S1). β2-adrenoceptor is a classic G-protein-coupled receptor used for investigation. Once the ligand binds to a β2-adrenoceptor, the receptor will change the conformation and couple to a heterotrimeric G-protein, resulting in an elevation of cAMP levels. Thus, hits obtained from the primary screening were also validated with another functional assay to measure the induction of cAMP expression. As shown in Figure 4B, three fractions had nearly the same intensity of cAMP induction as the positive control, isoproterenol. These two experiments confirm the effect of the three fractions on β2-adrenoceptor activation. Three positive fractions were consecutively obtained from Fructus Aurantii Immaturus. HPLC chromatograms of fractions 10-B11, 10-C11, and 10-D11 are shown in Figure 5. In addition, the methyl thiazolyl tetrazolium (MTT) test showed that fraction treatment at testing condition did not decrease cell viability (data not shown). All these results suggest that a cell-based functional assay system based on β-lactamase complementation technology has high selectivity for agonist identification. It is a useful tool for high-throughput screening and monitoring GPCR activation.

Positive hits validation. (A) Repeat of the receptor/β-arrestin interaction assay. Data represent means±SD for triplicate samples. (B) Positive hits validation by cAMP quantitation. Three hits have similar potential compared with positive control isoproterenol in elevating cAMP expression. Data represent means±SD for triplicate samples. cP<0.01 vs Mock.

HPLC chromatograms of three positive fractions. (A) Fraction 10-B11 and 10-C11 (Detector A-215 nm). (B) Fraction 10-D11 (Detector A-269 nm). The arrows indicated the positions of active fractions.
Discussion
We established a cell-based high-throughput screening assay system for monitoring G-protein-coupled receptor activation using β-lactamase complementation technology. Detection of β-lactamase activity recovery with CCF4/AM could be observed and imaged in intact cells by a fluorescence microscope. This system is also adapted for sorting or analyzing by flow cytometry. The two fragments of β-lactamase are only about 10 kDa and 19 kDa. They are relatively smaller than many other tag proteins, and their size decreases the possibility of altering the function of chimeric fusion proteins. The small size, monomeric characteristic, and availability of a cell-permeable fluorescent substrate suggest that a system based on the β-lactamase enzyme complementation has wide potential in biological studies and drug discovery.
GPCR activation leads to signal transduction through two mediators: G-proteins and β-arrestins. Agonists bind to GPCR and activate heterotrimeric G-proteins. This activation promotes second-messenger signaling and subsequently regulates different physiological functions. Another pathway for GPCR signal transduction is mediated by β-arrestins. The activated receptors can be phosphorylated by G-protein-coupled receptor kinases and recruit β-arrestins, which terminate signal transduction through G-proteins. Receptors move into clathrin-coated pits with β-arrestins from cell membranes, which cause receptor internalization and desensitization. Meanwhile, β-arrestins also serve as a scaffold for numerous signaling molecules that are stimulated by receptor agonists, which initiate a second parallel set of signals27. Traditionally, it was thought that ligand binding stimulated or inhibited all receptor functions to an equal extent, but in recent years, researchers have found “biased ligands” in many GPCRs28, 29, 30. Perfect biased ligands stimulate one receptor activity without stimulating other known receptor activity. The development of β-arrestin-biased ligands might provide opportunities for novel therapies31. Our cell-based model provided a tool for the measurement of agonist-promoted interactions between GPCRs and β-arrestins. Along with assays that measure second-messenger production (for example, Ca2+ or cAMP), our model remains promising in the search for a biased ligand specific to the GPCR signal pathway that can activate only G-protein- or β-arrestins-mediated signal transduction.
Traditional Chinese medicinal herbs are effective in the treatment of many diseases. In this study, we screened a traditional Chinese medicinal herb library for hits that could activate β2-adrenoceptor, recruit β-arrestin and cause the complementation of a β-lactamase fragment. Three hits were found from 352 fraction samples. Validated with the measurement of cAMP production, the three herbal extracts are each possible agonists of β2-adrenoceptor.
In summary, our achievement demonstrated that a cell-based assay using β-lactamase complementation technology was suitable for ligand screening. This assay was also validated on two other GPCRs: melanocortin receptor 4 and dopamine receptor 5 (data not shown). Our approach may provide an alternative strategy for scientists interested in this area.
Author contribution
Chuan-ke ZHAO designed and performed the research, analyzed the data and wrote the paper; Qi YIN discussed and revised the paper; and Shi-you LI revised the paper.
References
Pierce KL, Premont RT, Lefkowitz RJ . Seven-transmembrane receptors. Nat Rev Mol Cell Biol 2002; 3: 639–50.
Wise A, Jupe SC, Rees S . The identification of ligands at orphan G-protein coupled receptors. Annu Rev Pharmacol Toxicol 2004; 44: 43–66.
Wise A, Gearing K, Rees S . Target validation of G-protein coupled receptors. Drug Discov Today 2002; 7: 235–46.
Barak LS, Ferguson SS, Zhang J, Caron MG . A beta-arrestin/green fluorescent protein biosensor for detecting G protein-coupled receptor activation. J Biol Chem 1997; 272: 27497–500.
Zhang J, Barak LS, Anborgh PH, Laporte SA, Caron MG, Ferguson SS . Cellular trafficking of G protein-coupled receptor/beta-arrestin endocytic complexes. J Biol Chem 1999; 274: 10999–1006.
Angers S, Salahpour A, Joly E, Hilairet S, Chelsky D, Dennis M, et al. Detection of beta 2-adrenergic receptor dimerization in living cells using bioluminescence resonance energy transfer (BRET). Proc Natl Acad Sci USA 2000; 97: 3684–9.
Yan YX, Boldt-Houle DM, Tillotson BP, Gee MA, D'Eon BJ, Chang XJ, et al. Cell-based high-throughput screening assay system for monitoring G protein-coupled receptor activation using beta-galactosidase enzyme complementation technology. J Biomol Screen 2002; 7: 451–9.
Barnea G, Strapps W, Herrada G, Berman Y, Ong J, Kloss B, et al. The genetic design of signaling cascades to record receptor activation. Proc Natl Acad Sci USA 2008; 105: 64–9.
van Der Lee MM, Bras M, van Koppen CJ, Zaman GJ . Beta-arrestin recruitment assay for the identification of agonists of the sphingosine 1-phosphate receptor EDG1. J Biomol Screen 2008; 13: 986–98.
Zhao X, Jones A, Olson KR, Peng K, Wehrman T, Park A, et al. A homogeneous enzyme fragment complementation-based beta-arrestin translocation assay for high-throughput screening of G-protein-coupled receptors. J Biomol Screen 2008; 13: 737–47.
Doucette C, Vedvik K, Koepnick E, Bergsma A, Thomson B, Turek-Etienne TC . Kappa opioid receptor screen with the Tango beta-arrestin recruitment technology and characterization of hits with second-messenger assays. J Biomol Screen 2009; 14: 381–94.
Ghosh RN, DeBiasio R, Hudson CC, Ramer ER, Cowan CL, Oakley RH . Quantitative cell-based high-content screening for vasopressin receptor agonists using transfluor technology. J Biomol Screen 2005; 10: 476–84.
Garippa RJ, Hoffman AF, Gradl G, Kirsch A . High-throughput confocal microscopy for beta-arrestin-green fluorescent protein translocation G protein-coupled receptor assays using the Evotec Opera. Methods Enzymol 2006; 414: 99–120.
Bertrand L, Parent S, Caron M, Legault M, Joly E, Angers S, et al. The BRET2/arrestin assay in stable recombinant cells: a platform to screen for compounds that interact with G protein-coupled receptors (GPCRS). J Recept Signal Transduct Res 2002; 22: 533–41.
Vrecl M, Jorgensen R, Pogacnik A, Heding A . Development of a BRET2 screening assay using beta-arrestin 2 mutants. J Biomol Screen 2004; 9: 322–33.
Galarneau A, Primeau M, Trudeau LE, Michnick SW . Beta-lactamase protein fragment complementation assays as in vivo and in vitro sensors of protein protein interactions. Nat Biotechnol 2002; 20: 619–22.
Wehrman T, Kleaveland B, Her JH, Balint RF, Blau HM . Protein-protein interactions monitored in mammalian cells via complementation of beta-lactamase enzyme fragments. Proc Natl Acad Sci USA 2002; 99: 3469–74.
Philippon A, Dusart J, Joris B, Frere JM . The diversity, structure and regulation of beta-lactamases. Cell Mol Life Sci 1998; 54: 341–6.
Moore JT, Davis ST, Dev IK . The development of beta-lactamase as a highly versatile genetic reporter for eukaryotic cells. Anal Biochem 1997; 247: 203–9.
Zlokarnik G, Negulescu PA, Knapp TE, Mere L, Burres N, Feng L, et al. Quantitation of transcription and clonal selection of single living cells with beta-lactamase as reporter. Science 1998; 279: 84–8.
Milic M, Bao X, Rizos D, Liu F, Ziegler MG . Literature review and pilot studies of the effect of QT correction formulas on reported beta2-agonist-induced QTc prolongation. Clin Ther 2006; 28: 582–90.
Li SY, Chen C, Zhang HQ, Guo HY, Wang H, Wang L, et al. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antiviral Res 2005; 67: 18–23.
Zhang YL, Zhang HQ, Liu XY, Hua SN, Zhou LB, Yu J, et al. Identification of human dopamine receptors agonists from Chinese herbs. Acta Pharmacol Sin 2007; 28: 132–9.
Zhang JH, Chung TD, Oldenburg KR . A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J Biomol Screen 1999; 4: 67–73.
Ferguson SS, Menard L, Barak LS, Koch WJ, Colapietro AM, Caron MG . Role of phosphorylation in agonist-promoted beta 2-adrenergic receptor sequestration. Rescue of a sequestration-defective mutant receptor by beta ARK1. J Biol Chem 1995; 270: 24782–9.
Allen M, Hall D, Collins B, Moore K . A homogeneous high throughput nonradioactive method for measurement of functional activity of Gs-coupled receptors in membranes. J Biomol Screen 2002; 7: 35–44.
Lefkowitz RJ, Shenoy SK . Transduction of receptor signals by beta-arrestins. Science 2005; 308: 512–7.
Bisello A, Chorev M, Rosenblatt M, Monticelli L, Mierke DF, Ferrari SL . Selective ligand-induced stabilization of active and desensitized parathyroid hormone type 1 receptor conformations. J Biol Chem 2002; 277: 38524–30.
Holloway AC, Qian H, Pipolo L, Ziogas J, Miura S, Karnik S, et al. Side-chain substitutions within angiotensin II reveal different requirements for signaling, internalization, and phosphorylation of type 1A angiotensin receptors. Mol Pharmacol 2002; 61: 768–77.
Groer CE, Tidgewell K, Moyer RA, Harding WW, Rothman RB, Prisinzano TE, et al. An opioid agonist that does not induce micro-opioid receptor — arrestin interactions or receptor internalization. Mol Pharmacol 2007; 71: 549–57.
Violin JD, Lefkowitz RJ . Beta-arrestin-biased ligands at seven-transmembrane receptors. Trends Pharmacol Sci 2007; 28: 416–22.
Acknowledgements
This research was supported by the National Natural Science Foundation of China (grant No 30873103). We thank Xiao-yu LIU of HD Biosciences for assistance in the preparation of Chinese medicinal herb extracts.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhao, Ck., Yin, Q. & Li, Sy. A high-throughput screening system for G-protein-coupled receptors using β-lactamase enzyme complementation technology. Acta Pharmacol Sin 31, 1618–1624 (2010). https://doi.org/10.1038/aps.2010.154
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2010.154
