
Abstract
Pollen microspores from a hybrid between Lolium multiflorum (2n=4x=28) and Festuca arundinacea (2n=6x=42) were cultured and over 200 androgenic green plants established. In the initial hybrid one chromosome from each of the five homoeologous groups was labelled by a distinct PGI/2 homoeoallele. Segregation of PGI/2 alleles in the male gametes and in the androgenic plants indicated chromosome pairing preferences in the hybrid at meiosis. The pentaploid Festulolium hybrid genome comprised two homologous sets of Lolium (Lm) chromosomes and the polyhaploid set of F. arundinacea, namely one genome of F. pratensis (Fp) and two of F. glaucescens(Fg and Fg1). There was a high degree of preferential chromosome pairing between the two homologous sets of L. multiflorum chromosomes, and each androgenic plant contained a Lolium PGI/2 allele. Results supported earlier work indicating that gamete viability depended on the presence of a complete Lolium genome. Genomic in situ hybridization (GISH) was carried out on mitotic chromosome preparations of two androgenic plants using the total genomic DNA of L. multiflorum as a probe. Recombinants were observed between chromosomes of all three genomes in the pentaploid hybrid, demonstrating the efficacy of the hybrid as a starting point in introgression breeding programmes.
Similar content being viewed by others

Introduction
The genera Lolium and Festuca include most of the major temperate grasses, and together include a range of complementary, agronomically desirable traits (Thomas & Humphreys, 1991). Our ultimate aim is to create homozygous lines from Festulolium hybrids which combine the high nutritive value of Lolium with the stress tolerance of Festuca.
Androgenesis, or anther culture, allows us to produce monoploid plants from which we can eventually develop homozygous plants by chromosome doubling (Mayer et al., 1995). It can reveal novel characters governed by gene combinations, including recessive alleles, that are otherwise only rarely or never observed. This enables us to identify and select for such hidden traits. We describe here how androgenesis of a very amenable ×Festulolium pentaploid hybrid genotype which gave rise to a population of aneupolyhaploid plants enabled us to reveal much of the potential genotypic and phenotypic variation that can be generated through meiosis.
The Festulolium (5x) hybrid was constructed with one chromosome in each of the five homoeologous groups labelled by a distinct phosphoglucoisomerase (PGI/2) homoeoallele. We describe the segregation of PGI/2 homoeoalleles in the androgenic population and their use as indicators of chromosome behaviour at meiosis. In addition two androgenic plants, which combine particularly high drought and freezing tolerance with desirable agronomic characters, are characterized cytologically using genomic in situ hybridization (GISH).
Materials and methods
The pentaploid hybrid between L. multiflorum (Lm) (2n=4x=28)×F. arundinacea (Fa) (2n=6x=42) used for anther culture was produced as described by Humphreys (1989), and has a genomic constitution of LmLmFpFgFg1 (Fp=F. pratensis, FgFg1=F. glaucescens). The two homologous chromosomes of Lm are labelled at the PGI/2 locus by two alleles a and b, whereas the three homoeologous chromosomes of Fa are labelled by PGI/2 homoeoalleles a+, c and e. The c allele labelled a chromosome in the F. pratensis (Fp) genome, and the a+ and e PGI/2 alleles labelled chromosomes in each of the two F. glaucescens (Fg and Fg1) genomes of F. arundinacea (Humphreys, 1995). Because the hybrid is very responsive to cell culture (Humphreys & Dalton, 1992) and produces many inflorescences which are male fertile, it is ideally suited for anther culture. The hybrid was used previously in a backcrossing programme with L. multiflorum (2x) (Humphreys & Ghesquière, 1994).
The androgenic plants were produced following the procedures described by Ward (1993). Great care was taken to isolate and record the origin of different embryoids as they were formed so as to eliminate any possible risk that two or more embryoids might derive from the same microspore and have the same genotype. The PGI/2 phenotype of all androgenic plants was determined as described by Humphreys (1995).
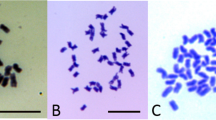
Two androgenic plants (genotypes 193 and 219) with proven high drought and freezing resistance (Zare, 1996) were selected for detailed cytological examination using genomic in situ hybridization (Thomas et al., 1994). Briefly, total genomic DNA of Lm labelled with rhodamine-4-dUTP was used as a probe on mitotic chromosome preparations of the two plants. As blocking DNA, ×40 probe concentration of Fa DNA was used with the Lm probe. Following GISH, chromosome preparations were counterstained with 4§,6-diamidino-2-phenylindole (DAPI). Chromosomes containing alien recombinant segments were identified as Lm, Fp, or Fg/Fg1 by the presence of the appropriate GISH labelling on both sides of the centromere.
Results
Phenotypic traits
A total of 221 green androgenic plants, derived from different microspores and thus having different genotypes, were successfully established in soil. The plants had very diverse expression of leaf size, tiller number, growth habit, plant height, root: shoot ratio, growth rate, tillering capacity, and tolerance to cold and drought (Zare, 1996). In particular, 6 per cent of the androgenic plants tested under simulated conditions were found to be more drought or cold tolerant than F. arundinacea, the more resistant parent (Humphreys et al., 1996).
Transmission of isozyme homoeoalleles
The frequencies of the five PGI/2 homoeoalleles in the androgenic population differed significantly (Table 1). This was caused primarily by the high frequency of the Lolium a allele (27 per cent) which was recovered at greater frequency than any other PGI/2 allele (16–19 per cent). The three Festuca PGI/2 alleles were recovered at similar frequency.
Almost all plants (98 per cent) carried at least one Lolium allele, and 10 per cent carried both (Table 2). The recovery of the Festuca alleles (a+, c and e) was less consistent: 30 per cent carried no Fg/Fg1 (a+ or e) allele, and 14 per cent carried both Fg and Fg1 alleles. The Fp c allele was recovered in 44 per cent of the androgenic plants (Table 2).
Further information on transmission of the five alleles is provided (Table 1) by comparing their frequency in the androgenic population with that in a BC1 population (produced by pollination diploid L. multiflorum with the pentaploid ×Festulolium hybrid, as reported by Humphreys & Ghesquière, 1994). There was no significant difference between the two populations in the frequency of PGI/2 homoeoalleles. Therefore, there was little or no selection in the BC1 against gametes with any Lm, Fp, Fg or Fg1 PGI/2 alleles, and hence it is likely that chromosomes from the homoeologous set were transmitted in accordance with their frequency in the male gamete.
Genome structure and recombination revealed by GISH
When the two androgenic plants which combined drought and freezing tolerance were studied using GISH (Fig. 1), the Lm DNA probe labelled with rhodamine-4dUTP hybridized preferentially with Lolium chromosomes (coloured pink) and with introgressed Lolium chromosome segments on Festuca chromosomes. The Lm DNA probe partially hybridized to certain Festuca chromosomes (coloured mauve) but failed to hybridize with other Festuca chromosomes (coloured blue). The Lm genome is known to be structurally closer to Fp than to Fg/Fg1 (Humphreys & Ghesquière, 1994) and the Festuca chromosomes which partially hybridized with the Lm probe were therefore ascribed to the Fp genome. In consequence, the chromosomes which showed no evidence of hybridization with the Lm probe belong to the Fg or Fg1 genomes. Chromosomes with partial hybridization to the Lm probe and carrying segments with no hybridization to the Lm probe are likely to be Fp chromosomes carrying Fg/Fg1 introgressions (for example Fig. 1b(4)).

Genomic in situ hybridization using Lolium multiflorum total genomic DNA as probe labelled with rhodamine 4dUTP on mitotic preparations of androgenic genotypes (a) 193, (b) 219, both 2n=21. Chromosomes which hybridize with the Lm probe are Lm (coloured pink); chromosomes which partially hybridize with the Lm probe are Fp (coloured mauve); and chromosomes with no hybridization to the Lm probe are Fg (coloured blue). Examples of interspecific recombinants are arrowed: (1) Fg or Fg1 chromosome with Lm recombinant, (2) Lm chromosome with Fg or Fg1 recombinant, (3) Lm chromosome 2 with Fp recombinant and (4) Fp chromosome with Fg or Fg1 recombinant. Lm, Lolium multiflorum; Fp, Festuca pratensis; Fg/Fg1, F. glaucescens. Bar=10 μm.
Genotype 193 had 21 chromosomes of which eight belonged to the Lm genome, seven to Fp and six to Fg/Fg1 (Fig. 1a). Genotype 219 also had 21 chromosomes: seven Lm, six Fp and eight Fg/Fg1 (Fig. 1b). Recombinants involving all three genomes were observed (an example of each combination is illustrated in Fig. 1) and this was confirmed with GISH using total genomic DNA probes of Fp and Fg/Fg1 in addition to Lm (not shown). One of the recombinants in 219 was derived from Fp, and replaced most of the long arm of a Lm chromosome with a large satellite identified as chromosome 2 (Fig. 1b(3)).
Discussion
Despite high levels of homoeologous chromosome pairing in pentaploid Festulolium hybrids (Humphreys, 1989), certain chromosome pairing preferences might be anticipated. For example, the two homologous Lm genomes would be expected to pair preferentially, and earlier meiotic chromosome studies support this conclusion (for example Morgan et al., 1988; Humphreys, 1989).
The analysis of segregation of PGI/2 alleles in the androgenic population described here provides clear indications as to whether these predictions for chromosome behaviour at meiosis are accurate. The presence of a Lm PGI/2 allele in 98 per cent of androgenic plants is strong evidence for preferential chromosome pairing and near-regular disjunction between the two Lm genomes of the pentaploid hybrid. This adds to evidence from backcross breeding programmes (Morgan et al., 1988; Humphreys & Ghesquière, 1994) that each gamete in the pentaploid hybrid contains a complete Lm genome. The low frequency (2 per cent) of plants with no Lm PGI/2 alleles shows that a complete Lm genome is nearly always required to provide gamete viability. On the other hand, the existence of a large proportion of androgenic plants with either no Fp (56 per cent) or no Fg or Fg1 (30 per cent) PGI/2-labelled chromosomes indicates that a complete Festuca genome is not essential for gamete viability.
A lower than expected recovery of a PGI/2 allele in the BC1 would have been evidence of gametophytic selection through pollen competition or zygotic abortion, which can occur in backcross breeding programmes and preclude recovery of certain gene combinations (Humphreys & Thorogood, 1993). However, no difference was observed between the segregation and recovery of the five PGI/2 alleles in androgenic plants with their transmission into the BC1 from Lm (2x)×Festulolium (5x) hybrids (see Humphreys & Ghesquière, 1994). Therefore, it is likely that there is no serious barrier to any gene combination in ×Festulolium hybrids.
The GISH chromosome study of the two androgenic plants demonstrates that chromosomes from Lm, Fp and Fg/Fg1 genomes are present and can be distinguished by their degree of hybridization with the Lm-DNA probe (Fig. 1). Recent studies (not shown) using Southern hybridization to determine genome relationships between Lm and the different genomes of Fa, support the conclusions made here for close homology between Lm and Fp and lower homology with Fg/Fg1. This ability to distinguish all three genomes using a single probe facilitates a rapid cytological analysis of hybrids involving these species.
We observed (Fig. 1) a number of reciprocal recombinants involving the Lm, Fp and Fg/Fg1 genomes. In contrast, Humphreys & Ghesquière (1994) were unable to observe any Lm−Fg or Lm−Fg1 recombinants among the BC1 in their breeding programme. We therefore conclude that gametes carrying Lm−Fg/Fg1 recombinants must face severe selection pressure or, following fertilization, produce nonviable zygotes. Indeed, certain gene combinations resulting from Lm−Fg/Fg1 chromosome recombination which can be recovered from anther culture may never be transmitted by conventional breeding methods.
In genotype 219, a large Fp chromosome recombinant occupies nearly the entire length of the long arm of chromosome two of Lm (Fig. 1b(3)). Because this chromosome arm of Fp is known to carry genes for drought resistance (Humphreys & Pašakinskien˙e, 1996), it is likely that these Fp genes have also contributed to the genotype's extreme drought resistance.
In order to utilize the variation obtained by anther culture, it will be necessary to restore fertility to selected androgenic plants and to incorporate them in breeding programmes. Improved fertility and genome stability should result from colchicine treatment, which will induce chromosome doubling and thereby encourage preferential pairing and regular disjunction between homologous chromosomes.
References
Humphreys, M. W. (1989). The controlled introgression of Festuca arundinacea genes into Lolium multiflorum. Euphytica. 42: 105–116.
Humphreys, M. W. (1995). Gene Introgression Following Intergeneric Hybridization Within the Lolium/Festuca Complex Ph.D. Thesis, University of Wales, Aberystwyth.
Humphreys, M. W. and Dalton, S. J. (1992). Stability at the phosphoglucoisomerase (PGI/2) locus in Lolium multiflorum(2n=4x=28)×Festuca arundinacea (2n=6x= 42) plants regenerated from cell suspension. Genome. 35: 461–467.
Humphreys, M. W. and Ghesquière, M. (1994). Assessing success in gene transfer between Lolium multiflorum and Festuca arundinacea. Euphytica. 77: 283–289.
Humphreys, M. W. and Pašakinskienė, I. (1996). Chromosome painting to locate genes for drought resistance transferred from Festuca arundinacea into Lolium multiflorum. Heredity. 77: 530–534.
Humphreys, M. W. and Thorogood, D. (1993). Disturbed Mendelian segregations at isozyme marker loci in early backcrosses of Lolium multiflorum×Festuca pratensis hybrids to L. multiflorum. Euphytica. 66: 11–18.
Humphreys, M. W., Zare, A. G. and Paškinskienė, I., Kerlan, M- C (1996). The potential of anther culture derived plants from a L. multiflorum × F. arundinacea (2n=5x=35) hybrid as a novel forage grass crop. Proceedings of the Eucarpia Fodder Crops and Amenity Grasses Section Plant Breeding and Acclimatization Institute, Radzikow, Poland, Oct. 7–10, 1996, pp. 155–156.
Mayer, M., Gland, G., Ceccarelli, S. and Geiger, H. H. (1995). Comparison of doubled haploid lines and F2 bulks for the improvement of barley in the dry areas of North Syria. Pl Breed. 114: 45–49.
Morgan, W. G., Thomas, H. and Lewis, E. J. (1988). Cytogenetic studies of hybrids between Festuca gigantea Vill. and Lolium multiflorum Lam. Pl Breed. 101: 335–343.
Thomas, H. and Humphreys, M. O. (1991). Progress and potential of interspecific hybrids of Lolium and Festuca. J Agric Sci Camb. 117: 1–8.
Thomas, H. M., Morgan, W. G., Meredith, M. R., Humphreys, M. W., Thomas, H. and Leggett, J. M. (1994). Identification of parental and recombined chromosomes in hybrid derivatives of Lolium multiflorum×Festuca pratensis by genomic in situ hybridization.. Theor Appl Genet. 88: 909–913.
Ward, S. P. (1993). The Use of Interspecific Hybrids in Forage Grass Breeding. M.Sc. Thesis, University of Wales, Aberystwyth.
Zare, A. G. (1996). Anther Culture in Intergeneric Hybrids of Forage Grasses. Ph.D. Thesis, University of Liverpool, Liverpool.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Humphreys, M., Zare, A., Pašakinskienė, I. et al. Interspecific genomic rearrangements in androgenic plants derived from a Lolium multiflorum×Festuca arundinacea (2n=5x = 35) hybrid. Heredity 80, 78–82 (1998). https://doi.org/10.1046/j.1365-2540.1998.00285.x
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.1998.00285.x
Keywords
This article is cited by
-
Gametic embryogenesis and haploid technology as valuable support to plant breeding
Plant Cell Reports (2011)
-
Androgenesis as a means of dissecting complex genetic and physiological controls: selecting useful gene combinations for breeding freezing tolerant grasses
Euphytica (2007)
-
Genome constitution and evolution in Lolium × Festuca hybrid cultivars (Festulolium)
Theoretical and Applied Genetics (2006)
-
Introgression-mapping of genes for drought resistance transferred from Festuca arundinacea var. glaucescens into Lolium multiflorum
Theoretical and Applied Genetics (2005)
