
Abstract

The tail of most sharks has an elongated upper lobe that differs from the externally symmetrical tail structure common among bony fishes, but the hydrodynamic purpose of this asymmetric tail shape is unclear1,2,3. Here we quantify water flow patterns in the wakes of freely swimming dogfish sharks and find that they have a ring-within-a-ring vortex structure, in contrast to the single rings shed by symmetrical fish tails. The branched-ring vortex is generated by the inclined angle of the tail's trailing edge and by its motion at an angle to the horizontal body axis; the vortex directs water backwards and downwards, which may increase the shark's vertical manoeuvrability.
Similar content being viewed by others

Main
We used digital particle-image velocimetry4,5 to analyse the hydrodynamic function of the tail in four spiny dogfish (Squalus acanthias; mean body length, 69 cm) swimming in a flow tank equipped with a vertical laser-light sheet. By using high-speed video (250 frames per second) to obtain particle images, we were able to analyse the time-dependent features of wake flow from the tail: six pairs of images were recorded throughout a tailbeat for five tailbeats from each of four individuals.
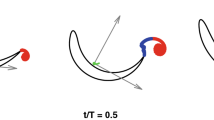
Three centres of vorticity were detected (Fig. 1a). The smaller dorsal vortex ring includes centres (1) and (2) and is contained within a larger vortex ring, which includes centres (1) and (3). Counter-rotating centres in the dorsal vortex ring produce jet A; a second jet B develops later as a result of flow induced by the ventral tip vortex. Jets A and B are directed on average at −35° to the freestream flow, before combining to produce a single broad jet C. Initially, little downstream flow is seen between same-sign vorticity centres (2) and (3) (Fig. 1a).

a, Vorticity plot of a vertical slice through tail vortex rings; curved arrows indicate the three centres of vorticity. b, Theoretically predicted velocity distributions across a planar section of a vortex ring (left) and a similar plot based on empirical data from the sequence shown in a along the ring axis (right). X', horizontal velocity; V', vertical velocity (both relative to the ring axis). c, Side and top views of a shark vortex wake. The axis of tail rotation (red dotted line) is inclined to the horizontal axis of locomotion (black dotted line), generating a ring-within-a-ring vortex structure by a mechanism similar to a piston with an inclined exit orifice10. Successive tailbeats generate linked rings (bottom). Green arrows indicate dorsal and ventral tip vortices and roll-up of the vortex sheet along the trailing edge.
Velocity transects though the centres of vorticity confirm the presence of a small dorsal vortex ring linked to a larger vortex ring that has a diameter equivalent to the tail height (Fig. 1b,c). Circulation calculations show that mean vortex-centre (1) circulation (0.0160 m2 s−1) is not significantly different from the sum of negative circulation in vortex centres (2) and (3) (−0.0019 and −0.0165, which is −0.0184 m2 s−1 in total; paired t-test, P = 0.3), as required by Kelvin's law. These findings contrast with the single vortex rings generated by the symmetrical tail of the bony fishes6,7. A strong posteroventrally directed jet flow is visible as a result of shark-tail motion (Fig. 1a,c), corroborating one classical model of shark-tail function2,8.
We propose that the mechanism for generating the ring-within-a-ring vortex structure in swimming dogfish is similar to that for fluid ejected from a pipe with an inclined opening (refs 910). Dorsal and ventral tip vortices (1) and (3) are shed as the tail beats from side to side. The dorsal lobe leads the ventral, so the tail presents an inclined edge to oncoming flow8. In a pipe with an inclined opening, a ring-within-a-ring vortex structure is generated owing to the temporal asymmetry in vortex roll-up between the longer and shorter edges9,10.
Previous work on the symmetrical tail of the bony fishes6,7,11 has shown that the vortex wake consists of single-ring vortices with dimensions similar to tail height and tailbeat width. The difference in wake structure between sharks and bony fishes seems to be due to their different tail shape and motion. The effects of these wake morphology differences on manoeuvring performance and locomotor energetics remain to be determined.
References
Grove, A. J. & Newell, G. E. Annu. Mag. Nat. Hist. 17, 280–290 (1936).
Alexander, R. M. T. J. Exp. Biol. 43, 131–138 (1965).
Thomson, K. S. Paleobiology 2, 19 (1976).
Drucker, E. G. & Lauder, G. V. J. Exp. Biol. 202, 2393–2412 (1999).
Wilga, C. D. & Lauder, G. V. J. Exp. Biol. 205, 2365–2374 (2002).
Lauder, G. V. Am. Zool. 40, 101–122 (2000).
Nauen, J. C. & Lauder, G. V. J. Exp. Biol. 205, 1709–1724 (2002).
Ferry, L. A. & Lauder, G. V. J. Exp. Biol. 199, 2253–2268 (1996).
Webster, D. R. & Longmire, E. K. Phys. Fluids 9, 655–666 (1997).
Webster, D. R. & Longmire, E. K. Phys. Fluids 10, 400–416 (1998).
Lauder, G. V. Am. Zool. 29, 85–102 (1989).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Wilga, C., Lauder, G. Hydrodynamic function of the shark's tail. Nature 430, 850 (2004). https://doi.org/10.1038/430850a
Published:
Issue Date:
DOI: https://doi.org/10.1038/430850a
This article is cited by
-
Fluid dynamic properties of shark caudal fin morphology and its relationship to habitats
Ichthyological Research (2024)
-
First insight into the swimming behavior of the paedomorphic fish Schindleria sp. (Gobiidae)
Ichthyological Research (2022)
-
Scalloped hammerhead sharks swim on their side with diel shifts in roll magnitude and periodicity
Animal Biotelemetry (2020)
-
Integument-based inferences on the swimming ability and prey hunting strategy of the bigeye thresher shark, Alopias superciliosus (Lamniformes: Alopiidae)
Zoomorphology (2020)
-
A survey on fabrication, control, and hydrodynamic function of biomimetic robotic fish
Science China Technological Sciences (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
