
Abstract
Both social environment and genetic factors are critical for smoking initiation and nicotine addiction. We reported that rats developed conditioned flavor (i.e., taste and odor) aversion to intravenously self-administered (IVSA) nicotine, and that social learning promoted nicotine IVSA with flavor cues. We thus tested the hypothesis that socially acquired nicotine IVSA is a heritable trait by using female rats of six inbred strains and six F1 hybrids. Each strain was tested for 10 daily IVSA sessions. We found that the intake of nicotine (15 and 30 μg/kg/inf) varied among these strains by 33.7–56.6-fold. The heritability of nicotine intake was estimated to be 0.54–0.65. Further, there was a strong correlation in nicotine intake (R2 = 0.85, p < 0.0001) between the two nicotine doses. Another cohort of rats was given three daily IVSA sessions followed by five sessions that tested conditioned flavor aversion. Nicotine intake was highly correlated with the extinction of the conditioned aversion (R2 = 0.58, p < 0.005). These data showed that nicotine intake in the socially acquired nicotine self-administration model is controlled by genetic factors and that the role of social learning is likely in facilitating the extinction of conditioned aversive response to nicotine.
Similar content being viewed by others


Introduction
Nicotine is the principal psychoactive ingredient of tobacco products1. It exerts marked reinforcing effects and maintains drug-seeking behavior2, 3. Paradoxically, nicotine is also aversive within the dose range where it is reinforcing. For example, doses of nicotine that maintained a high rate of responding in squirrel monkeys also induced vomiting4. In humans, the initial smoking experience is usually unpleasant and accompanied by physiological symptoms such as coughing, dizziness, and nausea5. The aversive nature of nicotine indicates that smoking initiation is likely influenced by other factors that could reduce the negative motivational effect of nicotine. One example of such a factor is the social environment. Not only is peer smoking the most significant predictor of smoking initiation6, but social interaction also increased the perceived reward of smoking7.
We previously established a model of socially acquired nicotine intravenous self-administration (IVSA) with an olfactogustatory cue in rats. We found that nicotine IVSA with a contingent appetitive oral flavor cue (i.e., taste and odor) resulted in conditioned flavor aversion (CFA)8. We also reported that social learning in this model was mediated by two factors: the odor (but not the taste) component of the flavor cue8 and carbon disulfide9, a component of mammalian breath.
Genetic factors contribute to approximately half of the variation in smoking behavior10, 11. Many studies have confirmed that variations in the CHRNA3-CHRNA5-CHRNB4 gene cluster are associated with smoking, particularly the number of cigarettes consumed per day12. Genes in this cluster, especially CHRNA5, have been shown to underlie the aversive effect of nicotine13, 14. However, this cluster accounted for only a small fraction of the heritability for smoking. Because of the large effect of social environment on smoking behavior, we hypothesized that genetic factors also play a role in this socially acquired nicotine IVSA model.
Although mouse has been the species of choice for behavioral genetic studies, the rat is emerging as a viable alternative, especially when complex behavioral paradigms are used15. We previously reported that genetic factors contributed to nicotine IVSA and food reward by using a lever press model in isogenic strains of adolescent rats16. Using female rats of the same panel of isogenic strains, we tested the above hypothesis using the socially- acquired nicotine self-administration model. Our results showed that nicotine intake in this model was highly heritable. In addition, we found a significant correlation between nicotine intake and the extinction of nicotine-conditioned aversion.
Results
Socially-acquired nicotine IVSA among 12 isogenic strains
IVSA, starting between postnatal day 41 and 44, were conducted in operant chambers that were divided by a partition. The partition had a row of six holes that allowed orofacial interaction between the two rats placed on each side. A flavor cue was provided to the self-administration rat via one of the two spouts (i.e., active) upon completing a fixed ratio 10 reinforcement schedule. Intravenous nicotine was delivered at that same time as the flavor cue. The flavor cue was also available without restriction on the side where a demonstrator rat was placed.
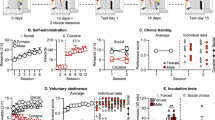
The numbers of nicotine infusions, licks on the active and inactive spouts during the ten nicotine IVSA sessions for the 12 isogenic strains are shown in Fig. 1 (15 μg/kg/inf) and Fig. 2 (30 μg/kg/inf). When 15 μg/kg/inf nicotine was provided, three strains (F344, FD, and FL) did not show a significant difference in the number of licks between the two spouts. All other strains licked more on the active spouts than on the inactive spouts (Table 1). At 30 μg/kg/inf nicotine, five strains licked more (BN, DA, FS, LB, and WL), and three strains (F344, FL, and LS) licked less on the active spout compared to the inactive spout. The other four strains did not show a main effect by spout (LEW, SHR, WKY, and FD) (Table 1).

Female adolescent rats from 12 isogenic strains were tested by using the socially acquired nicotine (15 μg/kg/inf) IVSA procedure. See Methods for a detailed description of the procedure. A logarithmic scale is used for the Y-axis.

Female adolescent rats from 12 isogenic strains were tested by using the socially acquired nicotine (30 μg/kg/inf) IVSA procedure. See Methods for a detailed description of the procedure. The Y-axis used a logarithmic scale.
Statistical analysis for the effect of the session on the number of licks and infusions is provided in Table 2 (15 μg/kg/inf) and Table 3 (30 μg/kg/inf). At 15 μg/kg/inf, eight strains (BN, DA, LEW, FD, FS, LB, LS, and WL) significantly increased the number of licks on the active spout across the sessions. Except FS, these strains also significantly increased the number of nicotine infusions across the sessions. At 30 μg/kg/inf, eight strains (except SHR, WKY, FD, and FL) increased the number of active licks across the sessions. Most of these strains (except FS) also significantly increased the number of nicotine infusions. F344 decreased the number of active licks and infusions across the sessions.
We found a strong correlation between the average number of nicotine infusions obtained when the two nicotine doses were tested across the 12 isogenic strains (Pearson coefficient = 0.917, p < 0.0001. Figure 3a). Further, the amount of nicotine intake was very similar between the two doses across the strains (Fig. 3b. F1,159 = 1.06, p > 0.05 for the effect of dose). On average, the ratio of nicotine intake at 30 μg/kg/inf. was 1.05 ± 0.1-fold of that at 15 μg/kg/inf. The stable nicotine intake at difference doses across 12 strains strongly suggested that nicotine, rather than the flavor cue, is the primary reinforcer of the operant behaviors.

Average nicotine intake during the last three IVSA sessions. The number of nicotine infusions was highly correlated when two different doses were tested (a). The gray area represents the 95% confidence interval of the linear model. When compared to the 30 μg/kg/inf dose, the number of infusions almost doubled across the strains when 15 μg/kg/inf was used. As a result, the amount of nicotine obtained was very similar between the two doses (b). Three-way repeated measures ANOVA found there was a significant effect of strain on nicotine intake (F11,160 = 9.0, p < 0.001) but dose (F1,166 = 0.13, p > 0.05) and session (F2,320 = 0.17, p > 0.05) had no effect.
We further compared the amount of nicotine intake during the last three IVSA sessions between the strains. Post-hoc analysis using Tukey HSD showed numerous significant strain differences. These results are provided in Fig. 4. The strain differences are very similar between the 15 and 30 μg/kg/inf doses, although the lower does tends to be more sensitive in detecting strain differences. Overall, these data suggested that genetic factors had a large effect on nicotine intake. We calculated the narrow sense heritability (h2) for nicotine intake to be 0.54 for 15 μg/kg/inf and 0.65 for 30 μg/kg/inf.

Strain difference. Strain differences in the average number of nicotine infusions obtained during the last three IVSA sessions were analyzed by post-hoc Tukey HSD tests. Each color block represented the p-value when the two strains listed on the x and y-axis were compared.
Conditioned flavor aversion
Our previous study showed that rats developed conditioned aversion to the flavor associated with self-administered nicotine8. We tested CFA after three nicotine (30 μg/kg/inf) IVSA sessions. Control rats received i.v. saline (Fig. 5). The data collected during the first three IVSA sessions showed that all 12 isogenic strains emitted significantly fewer licks on the active spout and obtained fewer infusions when nicotine was provided, compared to the saline groups (Table 4), indicating that nicotine was aversive. Ten strains (except SHR, and LS) showed significant CFA when tested on the fourth session (Table 4). After four more extinction sessions (i.e., rats received the flavor cue but not nicotine), six strains (BN, DA, FS, LB, LS, and WL) extinguished the CFA response (i.e., the number of active licks and saccharin reward were not statistically different between the saline and nicotine groups in these strains. Table 4).

Nicotine CFA and extinction of CFA. Twelve isogenic strains of adolescent rats self-ministered nicotine (30 μg/kg/inf, i.v.) with an appetitive flavor cue for three daily sessions without demonstrator rats (sessions 1–3). This was followed by five daily extinction sessions (sessions 4–8) where nicotine was not provided. Control rats received i.v. saline.
Correlation between CFA, extinction of CFA, and nicotine intake
We correlated the amount of nicotine intake and a CFA index across the 12 strains (Fig. 6a). Pearson correlation coefficient was -0.02 (R2 = 0.004, p = 0.93), indicating that the amount of nicotine intake was not correlated with the degree of CFA. However, there was a significant correlation between nicotine intake and the index for CFA extinction. The Pearson coefficient was 0.76 (R2 = 0.58, p < 0.005, Fig. 6b). This strong correlation indicated that the extinction of CFA is a main factor controlling nicotine intake. These data suggest that facilitating the extinction of nicotine CFA is a likely mechanism by which social learning promotes nicotine intake.

Correlation between nicotine intake and indices for CFA and extinction of CFA. The CFA index (a) was calculated using data obtained from the first extinction session (i.e. session four, Fig. 5). The extinction index (b) was calculated using data obtained from session eight. The correlation between the number of nicotine infusions at 30 μg/kg/inf with both indices were calculated. The gray area represents the 95% confidence interval of the linear model.
Discussion
We tested the socially- acquired nicotine IVSA model using 12 isogenic strains of adolescent rats. Our data showed that there was a large variation in the level of nicotine intake among these strains, ranging from an average of 1.8 to 42.4 infusions per 3 h session during the last three sessions. The amount of self-administered nicotine was very similar for each strain when two different doses of nicotine were provided, indicating strong self-regulation in drug intake. We estimated the heritability of nicotine intake to be 0.54–0.65. Although we showed that nicotine induced CFA in most of these strains, nicotine intake was not correlated with the degree of CFA. Instead, nicotine intake was correlated with the extinction of CFA, which suggested that the role of social learning is likely in facilitating the extinction of conditioned aversive response to nicotine.
We previously demonstrated many key features of the socially acquired nicotine self-administration model. For example, we have shown that strong aversive effect of nicotine can be overcome by social learning8. We also showed that the odor, rather than the taste cue associated with nicotine, is critical for social learning8. In addition, a gaseous component of the rodent breath, carbon disulfide, is needed for the effect of social learning9. We have further shown that social learning can facilitate nicotine IVSA even when the flavor cue is aversive (bitter)17, indicating that 1) nicotine has a strong reinforcing effect that is independent on the subjective value of the flavor cue and 2) social learning is a powerful modulator of reward. In addition to confirming many of these findings, the new data presented here demonstrated two additional salient aspects of our model: 1) the effect of social learning is likely in facilitating the extinction of nicotine CFA and 2) genetic factors play a critical role in nicotine intake in this model.
The involvement of social factors in the initiation of smoking has been well documented. For example, the majority of first-time cigarette uses usually occur in the presence of friends, and peer smoking is one of the strongest predictors of smoking initiation18, 19. However, mechanistic understanding of the role of social environment on smoking behavior is still lacking. By demonstrating the role of social learning in nicotine intake in rodents, our model provides a great opportunity to explore the mechanisms underlying the interaction between social factors and cigarette smoking.
Numerous human genetics studies have identified that variations in the CHRNA3-CHRNA5-CHRNAB4 cluster on chromosome 15 are associated with several aspects of the smoking behavior20, 21. Animal studies have shown that chrna5 is associated with the aversive effect of nicotine. Thus, we anticipated that the amount of nicotine intake in our model correlated with the aversive response measured by CFA. To our surprise, our data showed no such correlation (Fig. 6a). One likely reason is that our procedure models the acquisition phase of smoking, not chronic smoking, which is investigated in human genetic studies. Thus, although most strains developed conditioned aversive response to nicotine, it is not a strong determinant of nicotine intake during the early stages of nicotine consumption.
Several reports have shown that a social environment per se can be rewarding for rats in general22, 23. However, the data shown in Fig. 5b indicated that a potential function of the social environment has a more specific role in our model, which is to facilitate the extinction of conditioned aversive response to nicotine. This idea is in agreement with our previous finding that a social environment per se is not sufficient to promote nicotine intake with a flavor cue8. The distinction between enhancing reward vs. enhancing extinction of CFA is critical, because brain regions underlying these processes are likely to differ. For example, the mesolimbic circuit is likely involved in enhancing reward. On the other hand, many studies have shown that the infralimbic cortex is involved in the extinction of conditioned aversive stimuli, such as fear24. The infralimbic cortex also has been implicated in the extinction of cocaine25 and heroine-seeking behavior26. Therefore, our data suggested that the infralimbic cortex could potentially promote drug intake by facilitating the extinction of conditioned aversive response to the drug. We are currently investigating this hypothesis.
One limitation of this study is that we included only female rats. Possible sex differences in nicotine IVSA is a critical aspect of the animal models27 and have been studied by many research groups. For example, Lynch28 showed more females acquired nicotine IVSA than males. Swalve et al.29 found the opposite trend in acqusition but no sex difference in nicotine infusion post-acquisition. Chaudhri et al.30 studied the sex difference of nicotine IVSA using three nicotine doses (0.03, 0.06, 0.15 mg/kg) and found females obtained more nicotine when 0.06 mg/kg was used. However, Feltenstein31 showed there was no sex difference in nicotine IVSA when 0.03 or 0.05 mg/kg was used, and no estrous cycle dependent changes. Donny et al.32 showed that nicotine intake was similar between males and females but the motivation to obtain nicotine was higher in females (no effect of estrous cycle). More recently, Peartree et al.33 showed that a social context enhances the initial reinforcing effects of nicotine in males, but protects against nicotine intake during later sessions especially in females (again, estrous cycle had no effect). We have shown that in adolescent Sprague-Dawley rats, there was no sex difference in nicotine IVSA using either a 23-h access paradigm (lever pressing)34 or the socially acquired model8. However, in heterogenous stock rats, females self-administered significantly more nicotine than did males when the social model was used35. Thus, sex differences in nicotine IVSA are likely sensitive to the experimental condition and genetic background.
We have found many strain differences in nicotine intake in this study (Fig. 4). Most strains used in this study were also investigated in a previous report where nicotine self-administration was conducted using a lever response16. Although the estimated heritability of nicotine intake is very similar between these two studies, nicotine intake was very different for most strains. One potential difference is sex difference because only females were used in this study as discussed above. However, a more likely cause is the different cues used for nicotine intake: while a flavor cue and a social learning environment were provided in this study, a visual cue was used in our previous study, and rats were tested in a non-social setting. A third potential difference is that rats used in our previous experiment were food deprived and trained to press the lever for a food reward before nicotine was made available. In contrast, rats were not deprived of food or water in the socially acquired nicotine IVSA model, and thus were less likely to be affected by the motivational effect introduced by food restriction36.
Despite these differences, the F344 strain consistently had the lowest nicotine intake, while the LEW strain took moderate to high amounts of nicotine in both studies. This is consistent with a few studies that compared the effect of nicotine between these two strains. For example, F344 did not acquire nicotine IVSA in a 23-h access model, while the LEW strain did37,38,39. This difference is likely specific to nicotine, because these strains had similar behavior profiles when food reward was provided38; they also have a similar profile when tested for conditioned taste aversion included by LiCl40. Interestingly, the LEW strain is more sensitive to mecamylamine-precipitated withdrawal aversion than was the F344 strain41. Thus, one potential mechanism that can potentially reconcile these data is the hypothesis that the F344 strain has a strong acute aversive response to nicotine, as shown in Fig. 5, day 4. This effect is likely a dominant genetic effect, because the F1 crosses FD, FS, and FL all showed strong CFA, while minimal CFA was shown for the DA and SHR strains.
Together, these data showed that nicotine intake in the socially acquired nicotine IVSA model is under the control of the reward as well as the aversive effects of nicotine. The rewarding effect of nicotine was shown by the increased operant response when a lower dose (15 μg/kg/inf) of nicotine was given. The effect of the aversive property was shown by the strong CFA in most strains. In addition, social learning facilitates the extinction of CFA. By dissociating the aversive effect of nicotine and its cue (i.e., the oral flavor), social learning steers the balance towards a greater amount of nicotine intake. Last, the large strain difference indicated that there is a strong genetic control of nicotine intake.
Methods
Animals
Breeders of six inbred strains of rats, including Brown Norway (BN), Dark Agouti (DA), Fisher 344 (F344), Lewis (LEW), Spontaneously Hypertensive Rat (SHR), and Wistar Kyoto (WKY) were obtained from Harlan Laboratories (Indianapolis, IN). Rats were housed in a 12:12 h reversed light cycle (lights off at 9:00 am) with food and water available ad libitum. All animals for the IVSA experiments were bred on site. Inbreeding was maintained within each strain. As shown in Table 1, six selected F1 hybrids were also generated: LEW-BN (LB), F344-LEW(FL), F344-SHR (FS), WKY-LEW (WL), F344-DA (FD), LEW-SHR (LS). These F1s were selected based on the nicotine intake of the inbred strains reported previous16. The two letters for each F1 hybrid represented the initials of the maternal and paternal strains, respectively. Only adolescent rats were used because most smokers start smoking during adolescence and because our previous study showed that adolescent rats acquired nicotine IVSA at a faster rate and maintained higher nicotine intake than did adult rats34. Only female offspring were used because males were used in another study16. All procedures were approved by the Institutional Animal Care and Use Committee of the University of Tennessee Health Science Center. All procedures followed the NIH Guide for the Care and Use of Laboratory Animals.
Nicotine IVSA
Procedure for the socially acquired nicotine IVSA has been described in detail8. The surgery for inserting a jugular catheter and a 3D printed implant was described previously42. Surgery was conducted on postnatal day 38–40 under isoflurane anaesthesia. Ketoprofen (2 mg/kg, s.c.) was given for postoperative analgesia. The jugular catheter was flushed daily using 0.1 ml heparinized saline (100 IU/ml) and 0.1 ml Baytril (3 mg/kg) for 3 days after the surgery. Rats were then given access to nicotine IVSA sessions 3 h per day for 10 days in the dark-phase of the light cycle.
A detailed illustration of the operant chamber (Med-Associates, Inc.) has been provided elsewhere35. Each self-administration rat was paired with a randomly chosen conspecific (i.e., a demonstrator rat) during the experiment. These two rats were separated by a transparent plastic panel with six round holes that allow orofacial interactions. Two drinking spouts were installed on the side of the self-administration rat. Two syringe pumps were placed outside the sound attenuating chamber; one delivered nicotine (i.v.) through a swivel located on top of the chamber, and the other delivered the flavor cue (0.4% saccharin and 0.1% unsweetened grape-flavored Kool-Aid) to the active spout. The inactive spout contained no solution. Only one spout was available on the side of the demonstrator rat. A bottle installed on top of the spout provided unrestricted access to the same flavor solution to the demonstrator rat. The demonstrator rat received no nicotine injection. Each spout was connected to a contact lickometer controller allowing the number and the timing of licks to be recorded. IVSA was conducted using a fixed-ratio 10 schedule with 20 s timeout period. Thus, ten licks on the active spout activated the delivery of a 60 μl flavor cue, and an i.v. infusion (nicotine, free base, 15 or 30 μg/kg/inf., or saline).
Conditioned flavor aversion (CFA)
Two groups of rats were used for each strain. One group received nicotine (30 μg/kg/inf, i.v.), the other group received i.v. saline. Each group received three daily IVSA sessions as described above, with the exception that no demonstrator rats were provided. This was followed by five CFA test sessions where licking resulted in the delivery of the flavor cue but no i.v. infusion was given. We calculated the CFA index as the ratio of the reward earned (i.e., drops of the flavor cue) of the nicotine group to that of the saline controls in the first CFA test (i.e., session four). In addition, we also calculated the extinction index of CFA, which used the same calculation method but the data were obtained from the last CFA test session.
Statistical analysis
The number of licks was transformed to a log scale so that the data fit a normal distribution. Data are presentes as Mean ± SEM. Two-way repeated measures ANOVA was used to analyze the effect of strain on the number of nicotine infusions and licks, as well as the effect of the spout on the number of licks. Both session (i.e., day) and spout were treated as within-subject variables. The correlation between nicotine infusion during the last three IVSA sessions, the CFA index and the extinction index were calculated. All inbred and F1 crosses are isogenic, permitting the calculation of the narrow sense heritability. The between-strain variance provided a measure of additive genetic variation (VA), while within-strain variance represented environment variability (VE). An estimate of narrow-sense heritability (i.e., the proportion of total phenotypic variation that is due to the additive effects of genes, h2) was obtained using the formula: h2 = VA/(VA + VE). Statistical analyses were performed using the R statistical language. Statistical significance was assigned when p < 0.05.
References
Le Foll, B. & Goldberg, S. Effects of nicotine in experimental animals and humans: an update on addictive properties. Handb Exp Pharmacol 335–367 (2009).
Le Foll, B., Wertheim, C. & Goldberg, S. High reinforcing efficacy of nicotine in non-human primates. PLoS One 2, e230 (2007).
Lê, A. et al. Increased vulnerability to nicotine self-administration and relapse in alcohol-naive offspring of rats selectively bred for high alcohol intake. J Neurosci 26, 1872–1879 (2006).
Goldberg, S. & Spealman, R. Maintenance and suppression of behavior by intravenous nicotine injections in squirrel monkeys. Fed Proc 41, 216–220 (1982).
DiFranza, J. et al. Initial symptoms of nicotine dependence in adolescents. Tob Control 9, 313–319 (2000).
Flay, B. et al. Differential influence of parental smoking and friends’ smoking on adolescent initiation and escalation of smoking. J Health Soc Behav 35, 248–265 (1994).
Lochbihler, S., Miller, D. & Etcheverry, P. Extending animal models to explore social rewards associated with designated smoking areas on college campuses. J Am Coll Health 62, 145–152 (2014).
Chen, H., Sharp, B., Matta, S. & Wu, Q. Social interaction promotes nicotine self-administration with olfactogustatory cues in adolescent rats. Neuropsychopharmacology 36, 2629–2638 (2011).
Wang, T. & Chen, H. Carbon disulfide mediates socially-acquired nicotine self-administration. PLoS One 9, e115222 (2014).
Caron, L., Karkazis, K., Raffin, T., Swan, G. & Koenig, B. Nicotine addiction through a neurogenomic prism: ethics, public health, and smoking. Nicotine Tob Res 7, 181–197 (2005).
Hall, W., Gartner, C. & Carter, A. The genetics of nicotine addiction liability: ethical and social policy implications. Addiction 103, 350–359 (2008).
Tobacco and Genetics Consortium. Genome-wide meta-analyses identify multiple loci associated with smoking behavior. Nat Genet 42, 441–447 (2010).
Fowler, C., Lu, Q., Johnson, P., Marks, M. & Kenny, P. Habenular α5 nicotinic receptor subunit signalling controls nicotine intake. Nature 471, 597–601 (2011).
Frahm, S. et al. Aversion to nicotine is regulated by the balanced activity of β4 and α5 nicotinic receptor subunits in the medial habenula. Neuron 70, 522–535 (2011).
Parker, C. et al. Rats are the smart choice: Rationale for a renewed focus on rats in behavioral genetics. Neuropharmacology 250–258 (2014).
Chen, H., Hiler, K., Tolley, E., Matta, S. & Sharp, B. Genetic factors control nicotine self-administration in isogenic adolescent rat strains. PLoS One 7, e44234 (2012).
Wang, T., Han, W. & Chen, H. Socially acquired nicotine self-administration with an aversive flavor cue in adolescent female rats. Psychopharmacology (Berl) 233, 1837–1844 (2016).
Greenlund, K., Johnson, C., Webber, L. & Berenson, G. Cigarette smoking attitudes and first use among third- through sixth-grade students: the Bogalusa Heart Study. Am J Public Health 87, 1345–1348 (1997).
White, V., Byrnes, G., Webster, B. & Hopper, J. Does smoking among friends explain apparent genetic effects on current smoking in adolescence and young adulthood? Br J Cancer 98, 1475–1481 (2008).
Bierut, L. Convergence of genetic findings for nicotine dependence and smoking related diseases with chromosome 15q24-25. Trends Pharmacol Sci 31, 46–51 (2010).
Ware, J., van den Bree, M. & Munafò, M. Association of the CHRNA5-A3-B4 gene cluster with heaviness of smoking: a meta-analysis. Nicotine Tob Res 13, 1167–1175 (2011).
Thiel, K., Sanabria, F. & Neisewander, J. Synergistic interaction between nicotine and social rewards in adolescent male rats. Psychopharmacology (Berl) 204, 391–402 (2009).
Vanderschuren, L., Achterberg, E. & Trezza, V. The neurobiology of social play and its rewarding value in rats. Neurosci Biobehav Rev 70, 86–8105 (2016).
Giustino, T. & Maren, S. The Role of the Medial Prefrontal Cortex in the Conditioning and Extinction of Fear. Front Behav Neurosci 9, 298 (2015).
Cosme, C., Gutman, A., Worth, W. & LaLumiere, R. D1, but not D2, receptor blockade within the infralimbic and medial orbitofrontal cortex impairs cocaine seeking in a region-specific manner. Addict Biol (2016).
Chen, W. et al. Activation of AMPA receptor in the infralimbic cortex facilitates extinction and attenuates the heroin-seeking behavior in rats. Neurosci Lett 612, 126–131 (2016).
Perkins, K., Donny, E. & Caggiula, A. Sex differences in nicotine effects and self-administration: review of human and animal evidence. Nicotine Tob Res 1, 301–315 (1999).
Lynch, W. Sex and ovarian hormones influence vulnerability and motivation for nicotine during adolescence in rats. Pharmacol Biochem Behav 94, 43–50 (2009).
Swalve, N., Smethells, J. & Carroll, M. Sex differences in the acquisition and maintenance of cocaine and nicotine self-administration in rats. Psychopharmacology (Berl) 233, 1005–1013 (2016).
Chaudhri, N. et al. Sex differences in the contribution of nicotine and nonpharmacological stimuli to nicotine self-administration in rats. Psychopharmacology (Berl) 180, 258–266 (2005).
Feltenstein, M., Ghee, S. & See, R. Nicotine self-administration and reinstatement of nicotine-seeking in male and female rats. Drug Alcohol Depend 121, 240–246 (2012).
Donny, E. et al. Nicotine self-administration in rats: estrous cycle effects, sex differences and nicotinic receptor binding. Psychopharmacology (Berl) 151, 392–405 (2000).
Peartree, N. et al. Social context has differential effects on acquisition of nicotine self-administration in male and female rats. Psychopharmacology (Berl) (2017).
Chen, H., Matta, S. & Sharp, B. Acquisition of nicotine self-administration in adolescent rats given prolonged access to the drug. Neuropsychopharmacology 32, 700–709 (2007).
Wang, T. et al. Propensity for social interaction predicts nicotine-reinforced behaviors in outbred rats. Genes Brain Behav 13, 202–212 (2014).
Cabeza de Vaca, S., Krahne, L. & Carr, K. A progressive ratio schedule of self-stimulation testing in rats reveals profound augmentation of d-amphetamine reward by food restriction but no effect of a “sensitizing” regimen of d-amphetamine. Psychopharmacology (Berl) 175, 106–113 (2004).
Brower, V., Fu, Y., Matta, S. & Sharp, B. Rat strain differences in nicotine self-administration using an unlimited access paradigm. Brain Res 930, 12–20 (2002).
Sharp, B. et al. Gene expression in accumbens GABA neurons from inbred rats with different drug-taking behavior. Genes Brain Behav 10, 778–788 (2011).
Chen, H., Luo, R., Gong, S., Matta, S. & Sharp, B. Protection genes in nucleus accumbens shell affect vulnerability to nicotine self-administration across isogenic strains of adolescent rat. PLoS One 9, e86214 (2014).
Foynes, M. & Riley, A. Lithium-chloride-induced conditioned taste aversions in the Lewis and Fischer 344 rat strains. Pharmacol Biochem Behav 79, 303–308 (2004).
Suzuki, T., Ise, Y., Maeda, J. & Misawa, M. Mecamylamine-precipitated nicotine-withdrawal aversion in Lewis and Fischer 344 inbred rat strains. Eur J Pharmacol 369, 159–162 (1999).
Wang, T., Wang, B. & Chen, H. Menthol facilitates the intravenous self-administration of nicotine in rats. Front Behav Neurosci 8, 437 (2014).
Acknowledgements
Funding was provided by NIDA Grant DA-026894 (HC), DA-037844(HC), and the University of Tennessee Health Science Center faculty startup fund (HC).
Author information
Authors and Affiliations
Contributions
W.H. and T.W. conducted the experiments, analyzed the data, and drafted the manuscript. H.C. conceived the experiments, analyzed the data, and revised the manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Han, W., Wang, T. & Chen, H. Social learning promotes nicotine self-administration by facilitating the extinction of conditioned aversion in isogenic strains of rats. Sci Rep 7, 8052 (2017). https://doi.org/10.1038/s41598-017-08291-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-08291-5
This article is cited by
-
Social and anxiety-like behaviors contribute to nicotine self-administration in adolescent outbred rats
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
