

Key Points
- The oral cavity and oropharynx are spaces defined by both hard and soft tissue structures.
- The oral cavity has muscle and bony boundaries, but the pharynx is a muscular tube.
- An embryologic/evolutionary definition is useful for understanding the impact of birth defects.
- The complex and superficially idiosyncratic embryologic patterns result from well-defined evolutionary processes, producing a subtle anatomic pattern.
- A functional approach to anatomy can clarify normal and chronic or traumatic dyfunction.
- Functionally, the oral cavity and oropharynx are the pathway that food takes, their boundaries are the muscles that move food.
- Comprehension of oropharyngeal anatomy is the basis of modern clinical practice.
- Although our understanding of gross anatomy is unlikely to change in the future, advances in clinical treatment are predicated on the solid foundation of anatomic knowledge.
Introduction
The oral cavity and pharynx are anatomic spaces defined by hard and soft tissue structures (Figure 1). The shape of these two spaces changes with the normal physiologic function of the surrounding structures during speech, swallowing, and respiration. The oral cavity is bounded anteriorly by the lips, laterally by the cheeks, superiorly by the hard palate, and inferiorly by the mucosa covering the superior surface of the tongue and the sheet of muscles attaching to the inner side of the mandible, including geniohyoid, mylohyoid, and digastric. Although primarily a space through which food and air travel, several structures are found in this space, including the upper and lower dentition, the tongue, salivary glands, mucosal glands, and the mucosal tissue covering the hard palate, which bear the rugae. The oral cavity is continuous with the pharyngeal cavity, a more complex and somewhat irregular space.
Figure 1: Views of the oral cavity and pharynx.
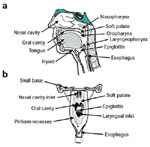
a: Midline sagittal view. b: Posterior view of the oral cavity and pharynx. In this view, the spinal column has been removed, so that the view is into the pharynx. The esophagus and pharynx have been opened to show the connections of the nasal cavity, oral cavity, larynx, and esophagus into the pharynx. The arrow (b, bottom left) represents the path that food takes through the piriform recesses, which lie on either side of the laryngeal opening.
The boundaries of the pharynx are the mouth and the nasal choanae anteriorly; the soft palate, or velum, and portions of the skull base superiorly; the posterior tongue inferiorly; and the pharyngeal constrictors posteriorly. In its anterior/inferior aspect, the pharynx joins the larynx; the adjacent lower portion is often referred to as the "hypopharynx" or the "laryngopharynx." In its anterior/superior potion the pharynx joins the nasal cavity and this upper portion is called the "nasopharynx." The midportion of the pharynx, where it joins the oral cavity, is called the "oropharynx." Thus the tongue lies in both the oral cavity and the pharynx. Various tonsils, composed of lymphoid tissue, are found in the pharynx. The pharyngeal tonsil is found in the roof of the nasopharynx. The palatine tonsils are bilateral structures in the oropharynx, and lingual tonsils lie on the posterior portion of the tongue.
There are several ways of describing the specifics and details of these spaces and the structures that bound them. One classic approach is based on embryology and the developmental history of these spaces.1, 2, 3 Because the embryology follows the evolutionary origin of these spaces, it has the virtue of delineating the evolutionary constraints of the spaces, and explaining some of the design flaws with respect to function. This approach is very useful for those working with the impact of morphologic birth anomalies on function, as well as the repair of those anomalies. A functional approach, starting with the physiology and mechanics of structures, followed by a description of the sensory and motor mapping to relevant structures is useful for the application of anatomy to understanding normal function, as well as chronic and traumatic dysmorphology.
There are many excellent texts of anatomy, replete with extensive detail.4, 5, 6, 7 Historically, anatomic atlases provide morphologic detail beyond that available in texts. Recent developments in imaging and publishing extend this tradition.1, 8, 9, 10 Because the substance of anatomy has changed little over the years, some of the older texts are still both relevant and useful.11, 12, 13 Additionally, there are numerous texts with a more physiologic outlook, that integrate some portion of anatomy with a more specific functional focus.12, 14, 15, 16, 17, 18 Finally, there are numerous texts describing, in detail, the embryologic history of the ultimate adult structures.2, 3, 12
This review does not duplicate those sources, but rather provides organization and overview so that basic scientists, clinicians, and other interested readers can utilize that detail as they wish. The information in this review is drawn from many of these sources. Occasionally there are conflicts among these sources (for example, in their description of the motor nerve supply to muscles in the pharynx), and we have tried to indicate such controversy. Thus, we present both an embryologic overview and a functional overview of these spaces and their associated structures.
Embryological Overview
The bony skull of adults forms from a complex set of embryologic precursors of multiple tissues. In general, bones of the mammalian skeleton have two different histologic-level origins: either preformed in cartilage (endochondral bone), or direct ossification of mesenchyme (membranous bone). Nearly all postcranial bones are endochondral, but many bones of the head and neck region are membranous bones.
Membranous bones, including the parietal, temporal, and frontal bones, as well as portions of the occipital bones, form the vault of the skull, and are derived from the bony armor of early fish. The maxilla and mandible, as well as portions of the palate and zygomatic bone, are also formed through direct ossification.
The endochondral bones of the skull have several different sources for the cartilage templates. The cranial base bones that underlie the brain are evolutionary descendants of the chondrocranium, a cartilaginous structure that surrounded the brain in early vertebrates. In humans, these tissues are initially a set of capsules (the olfactory, the optic, and the otic) that merge with a set of midline embryologic cartilaginous structures (the prechordal cartilage, the hypophyseal cartilage, and the parachordal cartilage). Together these form the ethmoid bone, the body, the lesser and greater wings of the sphenoid bone, the petrous portion of the temporal bone, and the base of the occipital bone.
Other bones formed from cartilage are derivatives of the pharyngeal arches, complex structures evolutionarily derived from structures that supported and contained the gills in early vertebrates. This set of five serial structures, also called the branchial arches, run from anterior to posterior. They begin to form at day 22 of human development (the beginning of the fourth week) in the head and neck region, but are found in all mammalian embryos. Each arch contains an external covering of ectodermal tissue; a middle core of mesodermal tissue, which will form nerve, muscle, and cartilage; and an inner covering of endoderm. The cartilage from the arches forms numerous oropharyngeal structures: the bones of the middle ear, the styloid process, the hyoid bone, and the cartilages of the larynx (Table 1). Tissue in each of the arches also becomes muscle, nerve, and blood vessels (Table 1). In general, arch one becomes the jaws, arch two becomes aspects of the face and ear, arch three becomes structures associated with the hyoid and upper pharynx, and arches four and six (arch five disappears) become structures associated with the larynx and lower pharynx. Numerous and detailed maps of the various tissues, as well as sensory fields, exist in many embryology texts.2, 3

The formation of the face, the palate, and the superior structures of the oral cavity are based on the dynamics of first arch development. The first arch splits early on into two recognizable entities: a mandibular and a maxillary portion. By the end of the fourth week of development, these in turn form five distinct swellings: two maxillary, two mandibular, and a superior, midline swelling: the frontonasal process. These tissues are invaded by cells called neural crest cells, which are responsible for the growth of these swellings. Eventually the fusion of these swellings forms the external face. A small gap becomes the mouth, and thickenings in the midline ectoderm eventually become pits that will become nares. Outgrowths of these tissues also become the palate.
Tissue from the pharyngeal arches is also responsible for the formation of the mucosa of the tongue. The complex patterns of both general and special sensation of the tongue are a reflection of its complex development. Mesoderm from the first arch will form the anterior two thirds of the tongue, up to the foramen cecum, whereas third and fourth arch mesoderm forms the posterior one third. Additional tissue from the occipital somites contribute to the tongue musculature. This tissue, originally posterior to the pharyngeal arches, is supplied by cranial nerve (CN) XII, the hypoglossal nerve.
The spaces between the arches externally are called clefts, internally pouches. The outer cleft between the first two arches contributes to the formation of the external acoustic meatus. The other clefts are absorbed during development. The corresponding inner pouch between the first two arches becomes the tympanic cavity and auditory or eustachian tube. The remainder of the pouches contribute to glandular tissue of the head and neck. These include the palatine tonsils from pouch two, the inferior parathyroid glands and thymus from pouch three, the superior parathyroid glands from pouch four, and the ultimobranchial body from the inferiormost portion of pouch four. Subsequent to formation, these tissues migrate to their adult locations during normal development.
These embryologic relationships, not to mention the adult anatomy, seem only marginally patterned. That is, although the cranial nerves are numbered from I to XII, their fields of innervation are overlapping, and not in a strict superior to inferior or anterior to posterior pattern. In organisms that retain a more primitive anatomy, such as early chordates and even some fish, amphibians, and reptiles, the adult structures are serially organized and accurately reflect the embryologic order of the arches, so that structures supplied by CN II are always anterior to those supplied by CN III. Significant evolutionary reorganization of the head and neck regions results in a mammalian pattern that is no longer linear and neatly organized.19
The evolutionary history that generated these modifications is fascinating. Two distinct factors operate here: constraint and adaptation. Evolution seldom generates morphology of optimal design for function. Instead, to meet the challenges of a changing environment and competition from other organisms, selection operates on variation generated through mutation and frequently small genetic changes that produce more significant changes in the developmental program of an organism.20 Thus the existing forms, such as the branchial or pharyngeal arches, pose a constraint. However, in adapting to new environments, such as land-dwelling and air-breathing, organisms modify the old structures for new functions.21 Subsequent adaptations for endothermy ("warm-blooded"), the requirement for higher energetic consumption, and for separation of deglutition and respiration modified the existing morphology further.22, 23, 24, 25
It is still useful, however, to appreciate the evolutionary linkage that is manifest in the embryologic development of humans. There are numerous clinical conditions that appear to influence diverse structures. Yet, the affected structures are united by embryologic origin (Table 1). These problems, often referred to as syndromes of the first and second arch include Goldenhar syndrome, mandibulofacial dysostoses such as Treacher-Collins or Hallermann-Streiff, and DiGeorge syndromes.26
Adult Anatomical Structures
There is a functional axis (the foodway) that follows the pathway of ingested food, from the mouth at the anterior end, moving posteriorly through the oral cavity, into the oropharynx, and then inferiorly through the hypopharynx, into the esophageal opening.
The bones that surround the first part of the foodway are the maxilla and mandible, the bones that bear the dentition, and the hard palate, which is composed of the palatine process of the maxilla and the maxillary process of the palatine bones. The cranial base, which forms the roof of the pharynx, is the body of the pterygoid bone, an endochondral bone. The latter part of the foodway is defined by muscle, but the hyoid bone and the cartilages of the larynx are pharyngeal arch structures that anchor this muscle.
The other significant hard tissue structures in the oral cavity are the teeth. The human dentition, rooted in the maxilla and mandible, consists of 32 teeth in four quadrants, left and right, upper and lower. Humans are born edentulous, and the first deciduous teeth erupt approximately 6 to 8 months after birth. There are five deciduous teeth per quadrant: a medial and a lateral incisor, a canine, and the first and second molars. These teeth are replaced by permanent or adult teeth, and additionally, two premolars and a third molar erupt as part of the adult dentition. All but the third molar are in place by approximately 12 years of age. The third molar, in modern humans, erupts much later, and often fails to erupt at all.
The muscles that form the walls of the oral cavity are simpler than those that form the walls of the pharynx. The orbicularis oris, which circles the opening of the mouth, and functions as a sphincter to close it, is the anterior boundary of the oral cavity. Other muscles that control the lips, and therefore the opening to the oral cavity are the labial muscles: levator labii superioris, depressor anguli oris, and risorius. The sides of the oral cavity can be considered the dentition, when the mouth is closed and jaw elevated, or the buccinator, the muscle that forms the cheeks. These are considered superficial facial muscles, and receive motor supply from branches of CN VII, the facial nerve.
The muscles of mastication, although not forming the boundaries of the oral cavity or pharynx, are critical for moving the jaws, and therefore oral function. Many of these muscles are supplied by V3 (mandibular branch of the trigeminal): temporalis, masseter, and medial pterygoid and lateral pterygoid.
The lower boundary of the oral cavity could be considered to be either the tongue or the muscles of the oral floor, which are responsible for opening the jaw. The tongue is a muscular hydrostat,27 a structure with muscles, but no skeletal support, capable of movement. It consists of four extrinsic muscles—genioglossus (XII), hyoglossus (XII), styloglossus (XII), and palatoglossus (X or XI)—and three groups of intrinsic fibers—vertical, transverse, and longitudinal fibers—all supplied by the hypoglossal nerve (XII). The genioglossus is an extrinsic muscle in that it originates on the mandible; however, its insertion merges into the intrinsic fibers with no clear demarcation. The neural supply to the tongue consists of three parts. The motor supply, detailed above, a general sensory component, which includes the lingual nerve (V3) to the anterior two thirds, branches of the glossopharyngeal nerve (IX) to the posterior one third, and a small area near the base supplied by the internal laryngeal nerve (X). The special sensation of taste is supplied by the chorda tympani, a branch of the facial nerve (VII) to the anterior portion, and by the glossopharyngeal nerve (IX) and the internal laryngeal (X) to the posterior one third.
The muscles of the oral floor are sheets of parallel fibered tissue, running from the hyoid bone to the mandible, and include digastric (V3 and VII), mylohyoid (V3), and geniohyoid (XII and C1). Lying on the oral side or inside of geniohyoid is one of the muscles of the tongue, the genioglossus (XII). The general sensation of the oral cavity is from branches of the trigeminal nerve (V). The upper parts, including palate and teeth, are innervated from branches of the maxillary nerve (V2), and the lower parts include oral floor mucosa from the mandibular nerve (V3). There is significant autonomic nerve supply to glandular tissue in the oral cavity. The sympathetic portions derive from the T1 level of the spinal cord, synapse in the superior cervical ganglion, and the postganglionic nerves travel with blood vessels into the target tissue. The parasympathetic structures are branches of the facial nerve (VII), but are distributed with branches of V2 or V3 to the upper or lower portions of the oral cavity. The digastric muscle is believed to be the principal muscle of jaw opening, whereas the geniohyoid is the principal muscle for elevation of the hyoid bone. The thyrohyoid muscle approximates the thyroid cartilage to the hyoid bone during swallowing; thus synergistic contraction of these muscles accounts for laryngohyoid elevation in swallowing.
As food is processed, either through trituration of hard food, or transport of liquids, it leaves the oral cavity and enters the pharynx. The boundaries between these two spaces are the hard–soft palate junction, the foramen cecum marking the boundary between the anterior two thirds and the posterior one third of the tongue, and most importantly, the palatopharyngeal fold of the fauces, containing the palatopharyngeus muscles (X or XI). This muscle originates anteriorly and superiorly from both the hard palate and the aponeurosis of the soft palate and then runs in an inferior/posterior direction into the wall of the pharynx. The palatopharyngeus functionally divides the oral cavity from oropharynx.
The soft palate is the superior boundary of the oropharynx and consists of several muscles joining in an aponeurosis: tensor veli palatini (V3), levator veli palatini (X or XI), palatopharyngeus (X or XI), uvulus (X or XI), and palatoglossus (X or XI). The principal elevator of the soft palate is the levator veli palatini, but all of these muscles play an important role in opening or closing the airway during swallowing. The posterior and lateral walls of the pharynx are formed by the groups of muscles known collectively as the pharyngeal constrictors. These muscles, uniting in a midline posterior raphe include the superior, middle, and inferior constrictors, all supplied by branches of the vagus nerve (X). Inside of the tube of muscle formed by the pharyngeal constrictors and in part of the wall are several longitudinal muscles. The stylopharyngeus runs from the styloid process into the pharyngeal wall, and the palatopharyngeus runs from the soft palate into this wall. These longitudinal muscles are also responsible for the maintenance of the pharyngeal portion of the airway, and may have a role in shortening the pharynx in swallowing (by raising the hypopharynx). The distal end of the pharynx is the upper esophageal sphincter (UES, X or XI), which consists of the lowest part of the inferior constrictor and, according to most authors, the surface of the cricoid cartilage. In some sources, this muscle is divided into the cricopharyngeus and the thyropharyngeus muscles. The cricopharyngeus muscle is unique in that it has no median raphe in its transverse portion. This muscle is tonically active between swallows, holding the UES closed. It is unclear whether the lower fibers of the thyropharyngeus and the upper fibers of the esophageal musculature also contribute to this sphincter closure. The UES represents the end of the pathway of food through the pharynx, and the point of entry of the bolus into the esophagus.
The sensory nerve supply to the pharynx is primarily from the glossopharyngeal and vagus nerves (IX and X). Branches of these nerves, including the pharyngeal branch of the glossopharyngeus (IX), small branches from the external and superior laryngeal nerves, and the pharyngeal branches, both from the vagus (X), form the pharyngeal plexus that lies on the pharyngeal wall. The exception is portions of the nasopharynx, which receive branches of the maxillary nerve (V2). The pharyngeal plexus also holds the motor neurons to the pharyngeal constrictor muscles.
The blood supply to the oral cavity and the pharynx is from branches of the external carotid artery. The branches of the external carotid that supply oral and pharyngeal structures are ascending pharyngeal a., lingual a., facial a., posterior auricular a., superficial temporal a., and numerous branches of the maxillary a. The venous drainage is through many tributaries, including facial, lingual, and pharyngeal vv.
Developmental Changes
Both continuous and discontinuous changes occur in the anatomy of the oral cavity and pharynx over the course of postnatal development. The most anatomically significant change to occur is the eruption of the deciduous dentition starting approximately 6 months after birth. However, the process of changing from a liquid diet to solid food is gradual in humans, and weaning often occurs over many months. Numerous other continuous changes are occurring during this period of time. The most functionally significant change is the descent of the larynx, and associated changes in airway protection.15, 28, 29 In the infant, the superior position of the larynx places its opening in the nasopharynx. This intranarial position means that infants, as is true of nearly all nonhuman mammals, are considered to have obligate nasal respiration.15 There are, however, pathologic conditions, such as adenoid hypertrophy, that result in oral respiration.30 Despite the "locking" of the larynx into the nasopharynx, respiration uniformly pauses during swallowing.31, 32 The laryngeal changes are coordinated with an increase in size of the oropharynx relative to the nasopharynx, changes in the angle of the basicranium, and others subtle changes in pharyngeal morphology that continue through adolescence.28, 33
Relevance of Anatomy for Clinical Practice
The descent of the hyoid bone and larynx in early postnatal development has important implications for swallowing and respiration, as well as communication. The neonatal position of the larynx definitively separates the airway from the foodway during tidal respiration. Food accumulating in the oral cavity and oropharynx is prevented from entering the hypopharynx or the larynx by the direct contact of the epiglottis and larynx with the enfolding tissues of the fauces and soft palate. This contact is broken briefly during swallowing, when the soft palate elevates and the epiglottis folds downward, but then is rapidly restored after the swallow. The intranarial position of the larynx also prevents airflow through the mouth, and thereby limits vocalization. The larynx is pulled inferiorly during infant cry, mimicking its adult position.
Laryngeal descent is essential for speech, because it enables airflow through the larynx, pharynx, and oral cavity. Speech obviously carries an enormous benefit for human communication. The question then arises, given the advantage of the descended larynx for speech, why this descent does not occur prenatally. Certainly there are issues regarding the cortical development necessary for speech, and evidence that this development occurs postnatally. But the postnatal development of cerebral capacity for spoken language does not require that development of the peripheral structural mechanisms for speech occur postnatally as well. There is no need to separate the airway from the foodway in utero, because both are filled with amniotic fluid. But the neonate must adjust to huge changes in environment at the moment of birth, with a need to rapidly fill both lungs with air. The intranarial larynx provides an upper airway that is structurally isolated from the oral cavity, with a semirigid cartilaginous framework for the upper airway. This simplifies the problem of establishing and maintaining airway patency, and reduces the likelihood of airway obstruction. Indeed, the relatively weak muscles of the neonatal pharynx and soft palate may need this structural support to maintain airway integrity for breathing, especially during sleep. The inability to maintain airway patency is a cause for obstructive sleep apnea.
The issue of swallow safety is also critical. Coordination of suckling, swallowing, and breathing is not fully developed in the neonate, creating a risk for deglutitive aspiration. The intranarial larynx prevents the bolus from entering the larynx before and after swallowing, and thus reduces the risk for airway obstruction and/or contamination by ingested milk or other material.
Conclusion
Although our understanding of gross anatomy has not changed substantially in decades, comprehension of that anatomy is the basis for modern clinical practice. Dysmorphology can only be assessed in relationship to normal morphology. Discussions of normal and abnormal function rely on a common vocabulary and a common map of structures. Oral and pharyngeal anatomy are particularly complicated, given the evolutionary and developmental constraints that dictate adult morphologic relationships. Although the field of gross anatomy is unlikely to see revolutionary changes in the future, our advances in clinical treatment are predicated on the solid foundation of anatomic knowledge.
