
Abstract
Autism spectrum disorders (ASDs) affect as many as 1 in 45 children and are characterized by deficits in sociability and communication, as well as stereotypic movements. Many children also show severe anxiety. The lack of distinct pathogenesis and reliable biomarkers hampers the development of effective treatments. As a result, most children with ASD are prescribed psychopharmacologic agents that do not address the core symptoms of ASD. Autoantibodies against brain epitopes in mothers of children with ASD and many such children strongly correlate with allergic symptoms and indicate an aberrant immune response, as well as disruption of the blood–brain barrier (BBB). Recent epidemiological studies have shown a strong statistical correlation between risk for ASD and either maternal or infantile atopic diseases, such as asthma, eczema, food allergies and food intolerance, all of which involve activation of mast cells (MCs). These unique tissue immune cells are located perivascularly in all tissues, including the thalamus and hypothalamus, which regulate emotions. MC-derived inflammatory and vasoactive mediators increase BBB permeability. Expression of the inflammatory molecules interleukin (IL-1β), IL-6, 1 L-17 and tumor necrosis factor (TNF) is increased in the brain, cerebrospinal fluid and serum of some patients with ASD, while NF-kB is activated in brain samples and stimulated peripheral blood immune cells of other patients; however, these molecules are not specific. Instead the peptide neurotensin is uniquely elevated in the serum of children with ASD, as is corticotropin-releasing hormone, secreted from the hypothalamus under stress. Both peptides trigger MC to release IL-6 and TNF, which in turn, stimulate microglia proliferation and activation, leading to disruption of neuronal connectivity. MC-derived IL-6 and TGFβ induce maturation of Th17 cells and MCs also secrete IL-17, which is increased in ASD. Serum IL-6 and TNF may define an ASD subgroup that benefits most from treatment with the natural flavonoid luteolin. Atopic diseases may create a phenotype susceptible to ASD and formulations targeting focal inflammation of the brain could have great promise in the treatment of ASD.
Similar content being viewed by others

Introduction
Autism spectrum disorders (ASDs) are pervasive neurodevelopmental disorders characterized by deficits in communication and social interactions, as well as the presence of stereotypic behaviors.1, 2, 3 Numerous gene mutations have been identified in patients with ASD, but no direct link has so far been uncovered except for a small percentage of cases associated with Tuberous Sclerosis, Fragile X syndrome, Rett syndrome and PTEN deficiency.4, 5 As a result, even though there are a number of genetically-engineered mice with phenotypes resembling autism,6 they do not adequately reflect ASD and there is an urgent need for appropriate animal ‘models’ of ASD.7 In fact, mouse ‘models’ are increasingly considered unreliable with respect to inflammatory human diseases.8 We recently reported that a small number of bull terriers develop symptoms consistent with autism and have increased serum neurotensin (NT) and corticotropin-releasing hormone (CRH), also found to be elevated in children with ASD.9
ASD may affect as many as 1 in 45 children in the USA,10 but the global prevalence is still under-recognized.11 The lack of reliable biomarkers12 and specific pathogenesis,13 as well as the existence of subgroups or comorbidities14 (Table 1), makes the development of specific treatments and conducting clinical studies difficult.13 As a result, child and adolescent outpatient mental health services in the USA have increased considerably.15 Moreover, the annual economic burden for ASD was recently estimated at $268 billion for 2015 and is projected to reach $416 billion in 2025.16
A number of perinatal allergic, genetic, environmental, immune and infectious factors may increase the risk of or contribute to the pathogenesis of ASD17, 18, 19 (Table 2). These could act through activation of a unique tissue immune cell, the mast cell (MC).20, 21 MCs derive from bone marrow progenitors and mature in tissues depending on microenvironmental conditions.22 In addition to histamine, stimulated MCs secrete other vasoactive and pro-inflammatory mediators such as the preformed kinins and proteases, as well as the de novo synthesized leukotrienes, prostaglandins, chemokines (CCXL8, CCL2), cytokines (interleukin (IL)-4, IL-6, IL-1, tumor necrosis factor (TNF)) and vascular endothelial growth factor (VEGF).20
MCs are not only considered critical for the development of allergic reactions,20 but also for immunity22 and inflammation.23 In fact, many studies have reported that allergic diseases in preschoolers are strongly associated with psychological and behavioral problems.24 We had proposed that MC-derived mediators could disrupt the blood–brain barrier (BBB) and cause 'allergy of the brain’25 or ‘focal encephalitis’,26 thus contributing to the pathogenesis of ASD.26, 27 A number of recent reviews have now confirmed and expanded on these findings.28, 29
Maternal health, prematurity and low birth weight are linked to increased risk of ASD
Obesity during gestation has been strongly associated with prematurity and low birth weight.30, 31 Obesity is considered as an inflammatory state32 and has been associated with activation of MCs.33, 34 Moreover, MCs secrete leptin35 and its deficiency switches MC to an anti-inflammatory phenotype.36 Leptin is increased both in obesity37 and ASD.38 Premature births account for about 15% of all births in the USA and premature infants (32–36 weeks) make up most of the increased rate of prematurity.39 Such infants are at risk for neurologic injury40, 41 associated with decreased attention, increased anxiety, as well as social interaction and learning difficulties.42
A retrospective study reported that children <33 weeks gestation were associated with a twofold higher risk of ASD.43 One prospective study found that 26% very low birth weight (<1500 g) infants ((n=91), mean age of 22 months) developed ASD.44 There was a higher risk of infantile autism among children with low birth weight especially in mothers >35 years, foreign born and those who had psychoactive medicines during pregnancy.45 Another case-control population-based cohort study among Swedish children (n=408, born 1974–1993), reported that the risk of ASD was associated with being small for gestational age, daily maternal smoking in early pregnancy, maternal birth outside Europe and North America, a 5-min APGAR score <7 and congenital malformations.46
Perinatal stress has been linked to increased risk of ASD.18, 47 Such stress may be linked to sexual abuse that has been associated with higher risk of ASD.48, 49 ASD patients are prone to stress50 and a meta-analysis showed a strong correlation between the presence of anxiety disorders and ASD.51 In fact, anxiety was significantly correlated with repetitive behaviors in children with ASD.52 We reported that the peptides NT53 and CRH9 secreted under stress were increased in the serum of young children with ASD, as compared with normal controls.53 The highest expression of NT receptors in the human brain is in the amygdala,54 hypothalamus and area of Broca,55 which regulate emotions and language, respectively. Stress can activate MCs through CRH leading to increased BBB permeability.56 Moreover, CRH has synergistic actions with NT, stimulating secretion of VEGF and increasing vascular permeability.53 Human MCs express CRHR-1,57 activation of which by CRH leads to VEGF secretion and BBB disruption58 and NT stimulates secretion of VEGF.57
A recent review concluded that stress during gestation increases the risk for developing atopic diseases in infants.59 Moreover, stress has been associated with precipitating or worsening asthma60 and multiple sclerosis.61
Atopic diseases are strongly correlated with increased risk of ASD
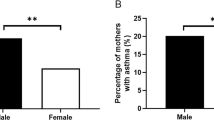
Recent studies have shown strong associations between allergies, asthma, autoimmune diseases and psoriasis in the mother with increased risk for ASD in their children.62, 63, 64 Moreover, mothers with mastocytosis or MC activation syndrome were much more likely to have children who developed ASD.65
Allergies66 and auto-immune diseases67, 68 have been increasing significantly. Early reports indicated more frequent allergies in ASD children,69, 70 with food allergies being the most prevalent complaint, often in the absence of elevated serum IgE or positive skin tests.71, 72, 73 A large epidemiological study of noninstitutionalized children (n=92 642; 0–17 years old) showed that eczema was strongly associated with ASD and attention deficit hyperactivity disorder.74 Another study of atopic subjects (n=14 812; 3 years old) and non-atopic subjects (n=6944) also showed a strong association between atopy and risk of both ASD and attention deficit hyperactivity disorder.75 A case control study of children and young patients with ASD (n=5565) and controls (n=27 825) matched to birth year (1980–2003) and sex reported that allergies, asthma and autoimmune disorders were diagnosed more frequently, with psoriasis occurring more than twice as often, in ASD patients than controls.76 An experimental study actually reported neurochemical changes and autistic-like behavior in a mouse model of food allergy.77
MCs can be activated by fungi,78 such as Aspergillus fumigatus which triggers IgE-independent MC degranulation79 and fungal zymosan induces leukotriene production from human MCs.80 Moreover, MCs can be stimulated by aluminum and mercury.81, 82
Perinatal epigenetic environmental triggers contribute to inflammation of the brain and increase risk of ASD
Environmental triggers have been increasingly invoked in ASD.17, 19, 83, 84, 85, 86 Chemical intolerant mothers were three times more likely to have a child who developed ASD and these children were more prone to allergies and sensitivities, including odors.87 Exposure to mold has been linked to decreased cognitive function in children88 and volatile mycotoxins have been reported to induce neuropsychiatric symptoms.89
Both mercury90 and aluminum91, 92 have been associated with symptom severity in children with ASD and both can stimulate MCs.81 Aluminum has replaced mercury as an adjuvant in vaccines, but aluminum can cause DNA damage93 and induce microglia TNF release.94 The adjuvant activity of aluminum was shown to be mediated through DNA released from dying cells, possibly through production of IgE and IgG1, known MC triggers.95 Such ‘damage-associated molecular patterns’ can act as ‘alarmins’96 and cause inflammatory responses through toll-like receptors, which participate in immunity against bacterial infections97, 98 and are also expressed on MCs.99
Stimulated human MCs can secrete mitochondrial DNA (mtDNA) and ATP extracellularly without cell death.100 These mitochondrial components augmented allergic responses101 and could act as ‘innate pathogens’ triggering inflammation and potentially contributing to ASD.102 mtDNA is also directly neurotoxic in rat brain slices.103 We reported that serum mtDNA is elevated in young autistic children as compared with controls.104 The pathological importance of extracellular mtDNA could be even more relevant in the subgroup of ASD patients with mitochondrial dysfunction.105
MCs are therefore considered important for inflammation.23,106
Evidence for inflammation of the brain in ASD patients
Increasing evidence indicates that perinatal brain inflammation,18, 107 may contribute to the pathogenesis of neuropsychiatric disorders,108, 109 including ASD.26, 110 It was previously reported that ASD pathogenesis involves some immune17, 111, 112, 113 and autoimmune102, 114 components. Circulating auto-antibodies directed against fetal brain proteins have been reported in mothers of children with ASD115, 116 and in about 37% of ASD patients,117 implying BBB disruption which is regulated through MCs.56, 118 The presence of auto-brain antibodies significantly correlated with allergic symptoms.119
A number of inflammatory molecules have been shown to be increased in the brain and cerebrospinal fluid of many ASD patients including IL-1β, IL-6, TNF, MCP-1 and CCL8 (IL-8) 120, 121, 122 (Table 3). Plasma levels of IL-1β, IL-6 and IL-8 were increased in children with ASD and correlated with regression, as well as impaired communication and aberrant behavior.123
Analysis of cytokines in neonatal blood showed that IL-1β and IL-4 linked to severe ASD.124 In a previous study by some of the same authors, these cytokines were not detected apparently due to the sensitivity of the assay used.125 Increased maternal serum concentrations of IFN-γ, IL-4 and IL-5 during midgestation were significantly associated with a 50% increased risk of ASD.126
MC-derived TNF can promote Th17-dependent neutrophil recruitment.127 Moreover, MC-derived IL-6 and TGFβ promote the devlopment of Th17 cells.128 In fact, MCs can also secrete IL-17129 and IL-17 was reported to be increased in the serum of children with ASD.130 There was an increased IL-17 production from peripheral blood immune cells following mitogen stimulation, and IL-17 was further increased in ASD children with comorbid asthma.131 A recent paper reported that selective elimination of Th17 cells in the maternal immune activation (MIA) mouse model prevented the development of autism-like behaivor in the offspring.132
The MIA model was also associated with increased serum IL-6,133 and the autism-like behavior was absent in IL-6−/− mice.134 We had reported that acute stress significantly increases serum IL-6 in mice that was entirely dependent on MCs, as it was absent in MC-deficient W/Wv mice.135 In fact, human MC can undergo selective release of IL-6 without degranulation.136 Mastocytosis patients have increased serum IL-6 that correlates with disease activity137, 138, 139 and children with mastocytosis had a 10-fold higher risk of developing ASD than the general population,65 implying activation of MCs.27
MCP-1 in amniotic fluid was strongly correlated with increased risk for infantile autism140 and MCP-1 was also elevated in archived neonatal blood specimens.125 MCP-1 is chemotactic for MCs,23 which can secrete both pre-formed and newly synthesized TNF.141 TGF-beta has been reported to be low in the brains of children with ASD,142 a finding that may contribute to the inflammatory state since TGF-beta inhibits MCs.143, 144
Peripheral blood mononuclear cells from patients with ASD (n=23) produced twice as much TNF as those from controls (n=13) when stimulated even by gliadin, cow’s milk protein or soy.145 NF-κB DNA-binding activity, involved in TNF production, was twice as much in peripheral blood from patients with ASD (n=67) than controls (n=29).146 Neurons, astrocytes and microglia from patients with ASD had higher expression of NF-κB p65 as compared with matched controls.147 Moreover, signaling through NF-κB was prominent in interacting gene networks constructed from brains of ASD patients.148
MCs have recently been considered important in neuroinflammation.149
MC–microglia interactions in the pathogenesis of ASD
Microglia, the innate brain immune cells,150 are important during healthy brain development because they may ‘prune’ neural circuits.151, 152 However, abnormal microglia activation and proliferation could lead to focal inflammation and ‘choking’ of normal synaptic traffic as has been reported in brains of patients with ASD.39, 153, 154, 155 A recent study of the transcriptomes from 104 human brain cortical tissue samples from patients with ASD identified gene clusters associated with increased microglia activation (M2) and decreased neuronal activity.156 As a result, microglia are now considered an important component of the pathogenesis of ASD.157, 158
Human microglia express functional CRHR1159 and NTR3 (sortilin), activation of which leads to microglia proliferation.160 NTR3 has been implicated in neuronal viability and function161 and increased soluble sortilin has been associated with depression, corresponding to elevated levels of BDNF and VEGF.162 NT can be neurotoxic by facilitating N-Methyl-d-aspartate-induced excitation of cortical neurons.163 We recently reported that NT stimulates activation and proliferation of human microglia.164 We believe this is the first time that a neuropeptide elevated in ASD is shown to stimulate human microglia that are now believed to play a major role in the pathogenesis of ASD.39, 153, 154 NT can therefore stimulate both microglia and MCs (Figure 1).53

Schematic representation of the interactions among mast cells–microglia–neurons and the blood–brain barrier. Curved arrows, along with mediators associated with them, indicate action from one type of cell to another. The inhibitory action of luteolin (in box) is indicated by the inhibitory symbols (T). CRH, corticotropin-releasing hormone; IL, interleukin; NT, neurotensin; TNF, tumor necrosis factor; VEGF, vascular endothelial growth factor.
Signaling through the mammalian target of rapamycin (mTOR) has been implicated in ASD5, 165 and mutations of the mTOR upstream regulatory molecule phosphatase and tensin homolog (PTEN)166 and tuberous sclerosis complex 1 and 2 (TSC 1/2)167 have been associated with higher risk of ASD.167 We recently showed that activation of NTR3 induced activation of human cultured microglia, which was regulated by mTOR.164 PTEN and mTOR are also involved in MC activation and proliferation.168
MC-derived histamine169 and tryptase170 can stimulate microglia, findings that have led to the proposal that MC-microglia interactions are important in neuroinflammation.171, 172 Stimulation of brain MC in mice was recently shown to induce microglia activation and brain inflammation, inhibited by a MC stabilizer.172
It is, therefore, important to address neuroinflammation as a possible treatment option for ASD.
Treatment approaches
Most children with ASD are often prescribed psychotropic medications,173 primarily risperidone and aripiprazole to reduce disruptive and aggressive behaviors, but these drugs have no effect on the core symptoms of ASD.174, 175 In fact, recent studies have questioned the benefit of psychotropic agents and have highlighted frequent adverse effects such as weight gain, sedation, tremor, movement disorders and drooling.176 As a result, there is increased polypharmacy174, 177 and risk of unwanted drug interactions.178
There should be concerted efforts toward developing effective treatments for ASD, such as the European Autism Interventions-A MultiCentre Study for Developing New Medications (EU-AIMS) Initiative.179
Immunomodulatory treatments have been considered for ASD,180 but few studies have been published. Some reports have hypothesized that the increase in ASD is linked to the increased use of the antipyretic acetaminophen.181 On the contrary, some families report that high fever reduces symptoms temporarily.182
Immune Ig
Intravenous (i.v.) immunoglobulin treatment (commonly known as immune Ig) has been used in ASD.183, 184 In one study, i.v. Ig (200 to 400 mg kg−1, every 6 weeks × 2) was administered to children with ASD (n=10) with one child showing significant and four children showing mild improvement.185 Three pilot open clinical trials showed some benefit.186, 187, 188
The usefulness of this approach may be even more apparent in children with ASD whose plasma levels of IgG and IgM were reported to be low in spite of apparently normal numbers of B cells.189
Macrophage activating factor (GcMAF)
This molecule, an endogenous glycosylated vitamin D-binding protein, which promotes macrophage cell activation, downregulated the over-activation of blood monocyte-derived macrophages observed in autistic children (n=22, 3–11 years old) compared with age-matched healthy developing controls (n=20).190
Antioxidant compounds
A recent double-blind, placebo-controlled, study using the broccoli-derived anti-oxidant sulforaphane in adult patients with ASD (n=40, 13–27 years old, selected for their history of reduced symptoms during febrile episodes) for 18 weeks showed significant improvement (34%) in social interaction and communication using the Aberrant Behavior Checklist (ABC) scale;191 however, the apparent significance was due to the uncharacteristically low placebo effect (<3.3%). Placebo effects have been reported as high as 40–60% in studies of neuropsychiatric diseases.192
Another antioxidant, N-acetylcysteine (NAC), has also been tested. In one randomized, placebo-controlled, trial (n=13) increasing doses of NAC (900 mg per day × 4 weeks, then 900 mg twice daily × 4 weeks and finally 900 mg three times daily × 4 weeks) found no difference on the total ABC, but significant improvement on the irritability subscale.193 In another also randomized, double-blind, placebo-controlled, study (n=40), NAC added to a stable dose of risperidone, again had no effect on total ABC, but decreased the irritability subscale.194, 195 NAC treatment appears to be safe and well-tolerated.195 Similar results were obtained in a more recent randomized, double-blind, placebo-controlled clinical trial of children with ASD (n=40) who were given NAC (600–900 mg per day) and risperidone titrated (between 1 and 2.0 mg per day); by week 10, the NAC group showed significantly less irritability using the ABC-C irritability subscale (P=0.02).196
Anti-inflammatory compounds
An open-label study investigated a formulation containing the natural flavonoids luteolin and quercetin ((100 mg each per softgel capsule in olive kernel oil to increase oral absorption) 1 capsule per 10 kg weight per day for 6 months) in children with ASD (4–10 years old, n=50) and reported significant (P<0.005) improvement in attention and behavior (34% in total ABC and 8.43 months in age-equivalent scores in the VABS communications domain).197 A subgroup of children in that study improved even more (65%) and were the ones with highest serum TNF and IL-6 at the beginning of the study, the levels of which dropped below basal levels at the end of treatment.122 These results indicate that objective inflammation markers may identify a subgroup of children with ASD, who are most amenable to treatment with luteolin or quercetin. A case series using the same formulation in children with ASD and atopic diseases (n=17, 4–12 years old) reported 65% improvement in attention and communication.198 Luteolin also improved ‘brain fog’, characterized by reduced attention span, memory and learning199 in adults.
Luteolin (5, 7, 3′, 4′-tetrahydroxyflavone) is naturally found in green plants, herbs and seeds 200 and is structurally related to 7, 8-dihydroxyflavone, which was shown to have brain-derived neurotrophic factor (BDNF) activity201 (Table 3). Low BDNF was associated with autistic-like-behavior in mice202 and 7, 8-dihydroxyflavone reduced symptoms in a mouse model of Rett syndrome,203 which is strongly associated with ASD.204
Luteolin and its structurally related flavonol quercetin (5, 7, 11, 3′, 4′-pentahydroxyflavonol) inhibit histamine, IL-6, IL-8, TNF and tryptase release from human MCs.205, 206 We recently showed that tetramethoxyluteolin is a more potent inhibitor of human MCs than luteolin.207 Luteolin also inhibits microglial activation and proliferation,208 especially IL-6 release,209 and is neuroprotective.210 Luteolin also prevented autism-like behavior in a mouse ‘model’ of autism.211 Flavonoids are generally considered safe212, 213 and now being increasingly discussed for the treatment of neurodegenerative disorders.214
Conclusions
Substantial evidence indicates that the presence of atopic diseases increases the risk of ASD and that inflammation of the brain may be involved in the pathogenesis of ASD. Addressing allergic symptoms, as well as reducing BBB permeability and inflammation of the brain, could provide significant benefit in ASD. Luteolin analogs with better bioavailability and BDNF activity should be investigated further. Intranasal administration to penetrate the brain through the cribriform plexus could deliver anti-inflammatory molecules directly to the brain. Such formulations could further be prepared in liposomes to contain molecules that target them to microglia.
References
Lai MC, Lombardo MV, Baron-Cohen S . Autism. Lancet 2014; 383: 896–910.
Volkmar FR, McPartland JC . From Kanner to DSM-5: autism as an evolving diagnostic concept. Annu Rev Clin Psychol 2014; 10: 193–212.
Fombonne E . Epidemiology of pervasive developmental disorders. Pediatr Res 2009; 65: 591–598.
Geschwind DH, State MW . Gene hunting in autism spectrum disorder: on the path to precision medicine. Lancet Neurol 2015; 11: 1109–1120.
Willsey AJ, State MW . Autism spectrum disorders: from genes to neurobiology. Curr Opin Neurobiol 2015; 30: 92–99.
Kazdoba TM, Leach PT, Crawley JN . Behavioral phenotypes of genetic mouse models of autism. Genes Brain Behav 2015; 15: 7–26.
Ruhela RK, Prakash A, Medhi B . An urgent need for experimental animal model of autism in drug development. Ann Neurosci 2015; 22: 44–49.
Seok J, Warren HS, Cuenca AG, Mindrinos MN, Baker HV, Xu W et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc Natl Acad Sci USA 2013; 110: 3507–3512.
Tsilioni I, Dodman N, Petra AI, Taliou A, Francis K, Moon-Fanelli AA et al. Elevated serum neurotensin and CRH levels in children with autistic spectrum disorders and tail-chasing bull terriers with a phenotype similar to autism. Transl Psychiatry 2014; 4: e466.
Developmental Disabilities Monitoring Network Surveillance Year 2010 Principal InvestigatorsCenters for Disease Control and Prevention (CDC). Prevalence of autism spectrum disorder among children aged 8 years - autism and developmental disabilities monitoring network, 11 sites, United States, 2010. MMWR Surveill Summ 2014; 63: 1–21.
Elsabbagh M, Divan G, Koh YJ, Kim YS, Kauchali S, Marcin C et al. Global prevalence of autism and other pervasive developmental disorders. Autism Res 2012; 5: 160–179.
Ruggeri B, Sarkans U, Schumann G, Persico AM . Biomarkers in autism spectrum disorder: the old and the new. Psychopharmacology (Berl) 2014; 231: 1201–1216.
Theoharides TC, Doyle R, Francis K, Conti P, Kalogeromitros D . Novel therapeutic targets for autism. Trends Pharmacol Sci 2008; 29: 375–382.
Bauman ML . Medical comorbidities in autism: challenges to diagnosis and treatment. Neurotherapeutics 2010; 7: 320–327.
Olfson M, Druss BG, Marcus SC . Trends in mental health care among children and adolescents. N Engl J Med 2015; 372: 2029–2038.
Leigh JP, Du J . Brief report: forecasting the economic burden of autism in 2015 and 2025 in the United States. J Autism Dev Disord 2015; 12: 4135–4139.
Rossignol DA, Frye RE . A review of research trends in physiological abnormalities in autism spectrum disorders: immune dysregulation, inflammation, oxidative stress, mitochondrial dysfunction and environmental toxicant exposures. Mol Psychiatry 2012; 17: 389–401.
Angelidou A, Asadi S, Alysandratos KD, Karagkouni A, Kourembanas S, Theoharides TC . Perinatal stress, brain inflammation and risk of autism-Review and proposal. BMC Pediatr 2012; 12: 89.
Herbert MR . Contributions of the environment and environmentally vulnerable physiology to autism spectrum disorders. Curr Opin Neurol 2010; 23: 103–110.
Theoharides TC, Valent P, Akin C . Mast cells, mastocytosis, and related disorders. N Engl J Med 2015; 373: 163–172.
Grimbaldeston MA . Mast cell-MrgprB2: sensing secretagogues or a means to overreact? Immunol Cell Biol 2015; 93: 221–223.
Galli SJ, Grimbaldeston M, Tsai M . Immunomodulatory mast cells: negative, as well as positive, regulators of immunity. Nat Rev Immunol 2008; 8: 478–486.
Theoharides TC, Alysandratos KD, Angelidou A, Delivanis DA, Sismanopoulos N, Zhang B et al. Mast cells and inflammation. Biochim Biophys Acta 2012; 1822: 21–33.
Chang HY, Seo JH, Kim HY, Kwon JW, Kim BJ, Kim HB et al. Allergic diseases in preschoolers are associated with psychological and behavioural problems. Allergy Asthma Immunol Res 2013; 5: 315–321.
Theoharides TC . Is a subtype of autism an 'allergy of the brain'? Clin Therap 2013; 35: 584–591.
Theoharides TC, Asadi S, Patel AB . Focal brain inflammation and autism. J Neuroinflammation 2013; 10: 46.
Theoharides TC, Stewart JM, Panagiotidou S, Melamed I . Mast cells, brain inflammation and autism. Eur J Pharmacol 2015; pii S0014-2999: 00398–2.
Young AM, Chakrabarti B, Roberts D, Lai MC, Suckling J, Baron-Cohen S . From molecules to neural morphology: understanding neuroinflammation in autism spectrum condition. Mol Autism 2016; 7: 9.
Kern JK, Geier DA, Sykes LK, Geier MR . Relevance of neuroinflammation and encephalitis in autism. Front Cell Neurosci 2015; 9: 519.
Zuckerman KE, Hill AP, Guion K, Voltolina L, Fombonne E . Overweight and obesity: prevalence and correlates in a large clinical sample of children with autism spectrum disorder. J Autism Dev Disord 2014; 44: 1708–1719.
Granich J, Lin A, Hunt A, Wray J, Dass A, Whitehouse AJ . Obesity and associated factors in youth with an autism spectrum disorder. Autism 2016; pii: 1362361315616345; e-pub ahead of print.
Gregor MF, Hotamisligil GS . Inflammatory mechanisms in obesity. Annu Rev Immunol 2011; 29: 415–445.
Theoharides TC, Sismanopoulos N, Delivanis DA, Zhang B, Hatziagelaki EE, Kalogeromitros D . Mast cells squeeze the heart and stretch the gird: their role in atherosclerosis and obesity. Trends Pharmacol Sci 2011; 32: 534–542.
Liu J, Divoux A, Sun J, Zhang J, Clement K, Glickman JN et al. Genetic deficiency and pharmacological stabilization of mast cells reduce diet-induced obesity and diabetes in mice. Nat Med 2009; 15: 940–945.
Taildeman J, Perez-Novo CA, Rottiers I, Ferdinande L, Waeytens A, De C . V et al. Human mast cells express leptin and leptin receptors. Histochem Cell Biol 2009; 131: 703–711.
Zhou Y, Yu X, Chen H, Sjoberg S, Roux J, Zhang L et al. Leptin deficiency shifts mast cells toward anti-inflammatory actions and protects mice from obesity and diabetes by polarizing M2 macrophages. Cell Metab 2015; 22: 1045–1058.
Lumeng CN, Saltiel AR . Inflammatory links between obesity and metabolic disease. J Clin Invest 2011; 121: 2111–2117.
Ashwood P, Kwong C, Hansen R, Hertz-Picciotto I, Croen L, Krakowiak P et al. Brief report: plasma leptin levels are elevated in autism: association with early onset phenotype? J Autism Dev Disord 2007; 38: 169–175.
Rodriguez JI, Kern JK . Evidence of microglial activation in autism and its possible role in brain underconnectivity. Neuron Glia Biol 2011; 7: 205–213.
Adams-Chapman I . Neurodevelopmental outcome of the late preterm infant. Clin Perinatol 2006; 33: 947–964.
Volpe JJ . The encephalopathy of prematurity—brain injury and impaired brain development inextricably intertwined. Semin Pediatr Neurol 2009; 16: 167–178.
Johnson S, Marlow N . Preterm birth and childhood psychiatric disorders. Pediatr Res 2011; 5, Pt 2 11 R–18R.
Limperopoulos C, Bassan H, Sullivan NR, Soul JS, Robertson RL Jr, Moore M et al. Positive screening for autism in ex-preterm infants: prevalence and risk factors. Pediatrics 2008; 121: 758–765.
Kleinman JM, Robins DL, Ventola PE, Pandey J, Boorstein HC, Esser EL et al. The modified checklist for autism in toddlers: a follow-up study investigating the early detection of autism spectrum disorders. J Autism Dev Disord 2008; 38: 827–839.
Maimburg RD, Vaeth M . Perinatal risk factors and infantile autism. Acta Psychiatr Scand 2006; 114: 257–264.
Hultman CM, Sparen P, Cnattingius S . Perinatal risk factors for infantile autism. Epidemiology 2002; 13: 417–423.
Ronald A, Pennell CE, Whitehouse AJ . Prenatal maternal stress associated with ADHD and autistic traits in early childhood. Front Psychol 2010; 1: 223.
Roberts AL, Koenen KC, Lyall K, Robinson EB, Weisskopf MG . Association of autistic traits in adulthood with childhood abuse, interpersonal victimization, and posttraumatic stress. Child Abuse Negl 2015; 45: 135–142.
Roberts AL, Lyall K, Rich-Edwards JW, Ascherio A, Weisskopf MG . Association of maternal exposure to childhood abuse with elevated risk for autism in offspring. JAMA Psychiatry 2013; 70: 508–515.
Vasa RA, Mazurek MO . An update on anxiety in youth with autism spectrum disorders. Curr Opin Psychiatry 2015; 28: 83–90.
van Steensel FJ, Bogels SM, Perrin S . Anxiety disorders in children and adolescents with autistic spectrum disorders: a meta-analysis. Clin Child Fam Psychol Rev 2011; 14: 302–317.
Rodgers J, Glod M, Connolly B, McConachie H . The relationship between anxiety and repetitive behaviours in autism spectrum disorder. J Autism Dev Disord 2012; 42: 2404–2409.
Donelan J, Boucher W, Papadopoulou N, Lytinas M, Papaliodis D, Theoharides TC . Corticotropin-releasing hormone induces skin vascular permeability through a neurotensin-dependent process. Proc Natl Acad Sci USA 2006; 103: 7759–7764.
Lantos TA, Palkovits M, Rostene W, Berod A . Neurotensin receptors in the human amygdaloid complex. Topographical and quantitative autoradiographic study. J Chem Neuroanat 1996; 11: 209–217.
Najimi M, Sarrieau A, Kopp N, Chigr F . An autoradiographic study of neurotensin receptors in the human hypothalamus. Acta Histochem 2014; 116: 382–389.
Esposito P, Chandler N, Kandere-Grzybowska K, Basu S, Jacobson S, Connolly R et al. Corticotropin-releasing hormone (CRH) and brain mast cells regulate blood-brain-barrier permeability induced by acute stress. J Pharmacol Exp Ther 2002; 303: 1061–1066.
Cao J, Papadopoulou N, Kempuraj D, Boucher WS, Sugimoto K, Cetrulo CL et al. Human mast cells express corticotropin-releasing hormone (CRH) receptors and CRH leads to selective secretion of vascular endothelial growth factor. J Immunol 2005; 174: 7665–7675.
Theoharides TC, Konstantinidou A . Corticotropin-releasing hormone and the blood-brain-barrier. Front Biosci 2007; 12: 1615–1628.
Hartmann K, Escribano L, Grattan C, Brockow K, Carter MC, varez-Twose I et al. Cutaneous manifestations in patients with mastocytosis: Consensus report of the European Competence Network on Mastocytosis; the American Academy of Allergy, Asthma & Immunology; and the European Academy of Allergology and Clinical Immunology. J Allergy Clin Immunol 2016; 137: 35–45.
Theoharides TC, Enakuua S, Sismanopoulos N, Papadimas E, Angelidou A, Alysandratos K . Stress contributes to asthma worsening through mast cell activation. Ann Allergy Asthma Immunol 2012; 109: 14–19.
Karagkouni A, Alevizos M, Theoharides TC . Effect of stress on brain inflammation and multiple sclerosis. Autoimmun Rev 2013; 12: 947–953.
Lyall K, Van de Water J, Ashwood P, Hertz-Picciotto I . Asthma and allergies in children with autism spectrum disorders: results from the CHARGE study. Autism Res 2015; 8: 567–574.
Chen SW, Zhong XS, Jiang LN, Zheng XY, Xiong YQ, Ma SJ et al. Maternal autoimmune diseases and the risk of autism spectrum disorders in offspring: a systematic review and meta-analysis. Behav Brain Res 2015; 296: 61–69.
Kotey S, Ertel K, Whitcomb B . Co-occurrence of autism and asthma in a nationally-representative sample of children in the United States. J Autism Dev Disord 2014; 44: 3083–3088.
Theoharides TC . Autism spectrum disorders and mastocytosis. Int J Immunopathol Pharmacol 2009; 22: 859–865.
Douwes J, Brooks C, van DC, Pearce N . Importance of allergy in asthma: an epidemiologic perspective. Curr Allergy Asthma Rep 2011; 11: 434–444.
Vermeersch P, Bossuyt X . Prevalence and clinical significance of rare antinuclear antibody patterns. Autoimmun Rev 2013; 12: 998–1003.
Hayter SM, Cook MC . Updated assessment of the prevalence, spectrum and case definition of autoimmune disease. Autoimmun Rev 2012; 11: 754–765.
Gurney JG, McPheeters ML, Davis MM . Parental report of health conditions and health care use among children with and without autism: National Survey of Children's Health. Arch Pediatr Adolesc Med 2006; 160: 825–830.
Magalhaes ES, Pinto-Mariz F, Bastos-Pinto S, Pontes AT, Prado EA, Deazevedo LC . Immune allergic response in Asperger syndrome. J Neuroimmunol 2009; 216: 108–112.
Bakkaloglu B, Anlar B, Anlar FY, Oktem F, Pehlivanturk B, Unal F et al. Atopic features in early childhood autism. Eur J Paediatr Neurol 2008; 12: 476–479.
Jyonouchi H . Autism spectrum disorders and allergy: observation from a pediatric allergy/immunology clinic. Exp Rev Clin Immunol 2010; 6: 397–411.
Angelidou A, Alysandratos KD, Asadi S, Zhang B, Francis K, Vasiadi M et al. Brief report: "allergic symptoms" in children with autism spectrum disorders. More than meets the eye?. J Autism Dev Disord 2011; 41: 1579–1585.
Yaghmaie P, Koudelka CW, Simpson EL . Mental health comorbidity in patients with atopic dermatitis. J Allergy Clin Immunol 2013; 131: 428–433.
Chen MH, Su TP, Chen YS, Hsu JW, Huang KL, Chang WH et al. Is atopy in early childhood a risk factor for ADHD and ASD? a longitudinal study. J Psychosom Res 2014; 77: 316–321.
Zerbo O, Leong A, Barcellos L, Bernal P, Fireman B, Croen LA . Immune mediated conditions in autism spectrum disorders. Brain Behav Immun 2015; 46: 232–236.
de Theije CG, Wu J, Koelink PJ, Korte-Bouws GA, Borre Y, Kas MJ et al. Autistic-like behavioural and neurochemical changes in a mouse model of food allergy. Behav Brain Res 2014; 261: 265–274.
Saluja R, Metz M, Maurer M . Role and relevance of mast cells in fungal infections. Front Immunol 2012; 3: 146.
Urb M, Pouliot P, Gravelat FN, Olivier M, Sheppard DC . Aspergillus fumigatus induces immunoglobulin E-independent mast cell degranulation. J Infect Dis 2009; 200: 464–472.
Olynych TJ, Jakeman DL, Marshall JS . Fungal zymosan induces leukotriene production by human mast cells through a dectin-1-dependent mechanism. J Allergy Clin Immunol 2006; 118: 837–843.
Kempuraj D, Asadi S, Zhang B, Manola A, Hogan J, Peterson E et al. Mercury induces inflammatory mediator release from human mast cells. J Neuroinflammation 2010; 7: 20.
McKee AS, Munks MW, MacLeod MK, Fleenor CJ, van RN, Kappler JW et al. Alum induces innate immune responses through macrophage and mast cell sensors, but these sensors are not required for alum to act as an adjuvant for specific immunity. J Immunol 2009; 183: 4403–4414.
Goines PE, Ashwood P . Cytokine dysregulation in autism spectrum disorders (ASD): possible role of the environment. Neurotoxicol Teratol 2012; 36: 67–81.
Rossignol DA, Genuis SJ, Frye RE . Environmental toxicants and autism spectrum disorders: a systematic review. Transl Psychiatry 2014; 4: e360.
Ming X, Brimacombe M, Malek JH, Jani N, Wagner GC . Autism spectrum disorders and identified toxic land fills: co-occurrence across States. Environ Health Insights 2008; 2: 55–59.
Wong CT, Wais J, Crawford DA . Prenatal exposure to common environmental factors affects brain lipids and increases risk of developing autism spectrum disorders. Eur J Neurosci 2015; 42: 2742–2760.
Heilbrun LP, Palmer RF, Jaen CR, Svoboda MD, Miller CS, Perkins J . Maternal chemical and drug intolerances: potential risk factors for autism and attention deficit hyperactivity disorder (ADHD). J Am Board Fam Med 2015; 28: 461–470.
Jedrychowski W, Maugeri U, Perera F, Stigter L, Jankowski J, Butscher M et al. Cognitive function of 6-year old children exposed to mold-contaminated homes in early postnatal period. Prospective birth cohort study in Poland. Physiol Behav 2011; 104: 989–995.
Empting LD . Neurologic and neuropsychiatric syndrome features of mold and mycotoxin exposure. Toxicol Ind Health 2009; 25: 577–581.
Bernard S, Enayati A, Roger H, Binstock T, Redwood L . The role of mercury in the pathogenesis of autism. Mol Psychiatry 2002; 7 (Suppl 2): S42–S43.
Geier DA, Kern JK, King PG, Sykes LK, Geier MR . Hair toxic metal concentrations and autism spectrum disorder severity in young children. Int J Environ Res Public Health 2012; 9: 4486–4497.
Tomljenovic L, Shaw CA . Do aluminum vaccine adjuvants contribute to the rising prevalence of autism? J Inorg Biochem 2011; 105: 1489–1499.
Kumar V, Bal A, Gill KD . Aluminium-induced oxidative DNA damage recognition and cell-cycle disruption in different regions of rat brain. Toxicology 2009; 264: 137–144.
Sood PK, Nahar U, Nehru B . Stress proteins and glial cell functions during chronic aluminium exposures: protective role of curcumin. Neurochem Res 2012; 37: 639–646.
Marichal T, Ohata K, Bedoret D, Mesnil C, Sabatel C, Kobiyama K et al. DNA released from dying host cells mediates aluminum adjuvant activity. Nat Med 2011; 17: 996–1002.
Zhang Q, Raoof M, Chen Y, Sumi Y, Sursal T, Junger W et al. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature 2010; 464: 104–107.
Galli SJ, Tsai M . Mast cells in allergy and infection: versatile effector and regulatory cells in innate and adaptive immunity. Eur J Immunol 2010; 40: 1843–1851.
Abraham SN St, John AL . Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol 2010; 10: 440–452.
Sandig H, Bulfone-Paus S . TLR signaling in mast cells: common and unique features. Front Immunol 2012; 3: 185.
Zhang B, Asadi S, Weng Z, Sismanopoulos N, Theoharides TC . Stimulated human mast cells secrete mitochondrial components that have autocrine and paracrine inflammatory actions. PLoS One 2012; 7: e49767.
Asadi S, Theoharides TC . Corticotropin-releasing hormone and extracellular mitochondria augment IgE-stimulated human mast-cell vascular endothelial growth factor release, which is inhibited by luteolin. J Neuroinflammation 2012; 9: 85.
Theoharides TC, Asadi S, Panagiotidou S, Weng Z . The "missing link" in autoimmunity and autism: Extracellular mitochondrial components secreted from activated live mast cells. Autoimmun Rev 2013; 12: 1136–1142.
Lauritzen KH, Moldestad O, Eide L, Carlsen H, Nesse G, Storm JF et al. Mitochondrial DNA toxicity in forebrain neurons causes apoptosis, neurodegeneration, and impaired behavior. Mol Cell Biol 2010; 30: 1357–1367.
Zhang B, Angelidou A, Alysandratos KD, Vasiadi M, Francis K, Asadi S et al. Mitochondrial DNA and anti-mitochondrial antibodies in serum of autistic children. J Neuroinflammation 2010; 7: 80.
Rossignol DA, Frye RE . Mitochondrial dysfunction in autism spectrum disorders: a systematic review and meta-analysis. Mol Psychiatry 2012; 17: 290–314.
Galli SJ, Tsai M, Piliponsky AM . The development of allergic inflammation. Nature 2008; 454: 445–454.
Hagberg H, Gressens P, Mallard C . Inflammation during fetal and neonatal life: implications for neurologic and neuropsychiatric disease in children and adults. Ann Neurol 2012; 71: 444–457.
Jones KA, Thomsen C . The role of the innate immune system in psychiatric disorders. Mol Cell Neurosci 2013; 53: 52–62.
Chavarria A, Alcocer-Varela J . Is damage in central nervous system due to inflammation? Autoimmun Rev 2004; 3: 251–260.
Le Belle JE, Sperry J, Ngo A, Ghochani Y, Laks DR, Lopez-Aranda M et al. Maternal inflammation contributes to brain overgrowth and autism-associated behaviors through altered redox signaling in stem and progenitor cells. Stem Cell Rep 2014; 3: 725–734.
Onore C, Careaga M, Ashwood P . The role of immune dysfunction in the pathophysiology of autism. Brain Behav Immun 2012; 26: 383–392.
Estes ML, McAllister AK . Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat Rev Neurosci 2015; 16: 469–486.
Bauman MD, Iosif AM, Smith SE, Bregere C, Amaral DG, Patterson PH . Activation of the maternal immune system during pregnancy alters behavioral development of rhesus monkey offspring. Biol Psychiatry 2014; 75: 332–341.
Gesundheit B, Rosenzweig JP, Naor D, Lerer B, Zachor DA, Prochazka V et al. Immunological and autoimmune considerations of Autism Spectrum Disorders. J Autoimmun 2013; 44: 1–7.
Braunschweig D, Van de Water J . Maternal autoantibodies in autism. Arch Neurol 2012; 69: 693–699.
Zimmerman AW, Connors SL, Matteson KJ, Lee LC, Singer HS, Castaneda JA et al. Maternal antibrain antibodies in autism. Brain Behav Immun 2007; 21: 351–357.
Rossi CC, Van de Water J, Rogers SJ, Amaral DG . Detection of plasma autoantibodies to brain tissue in young children with and without autism spectrum disorders. Brain Behav Immun 2011; 25: 1123–1135.
Ribatti D . The crucial role of mast cells in blood-brain barrier alterations. Exp Cell Res 2015; 338: 119–125.
Mostafa GA, Al-Ayadhi LY . The possible relationship between allergic manifestations and elevated serum levels of brain specific auto-antibodies in autistic children. J Neuroimmunol 2013; 261: 77–81.
Zimmerman AW, Jyonouchi H, Comi AM, Connors SL, Milstien S, Varsou A et al. Cerebrospinal fluid and serum markers of inflammation in autism. Pediatr Neurol 2005; 33: 195–201.
Li X, Chauhan A, Sheikh AM, Patil S, Chauhan V, Li XM et al. Elevated immune response in the brain of autistic patients. J Neuroimmunol 2009; 207: 111–116.
Tsilioni I, Taliou A, Francis K, Theoharides TC . Children with Autism Spectrum Disorders, who improved with a luteolin containing dietary formulation, show reduced serum levels of TNF and IL-6. Transl Psychiatry 2015; 5: e647.
Ashwood P, Krakowiak P, Hertz-Picciotto I, Hansen R, Pessah I, Van de Water J . Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav Immun 2011; 25: 40–45.
Krakowiak P, Goines PE, Tancredi DJ, Ashwood P, Hansen RL, Hertz-Picciotto I et al. Neonatal cytokine profiles asAnchorsociated with autism spectrum disorder. Biol Psychiatry 2015; S0006-3223(15)00655-1.
Zerbo O, Yoshida C, Grether JK, Van de Water J, Ashwood P, Delorenze GN et al. Neonatal cytokines and chemokines and risk of Autism Spectrum Disorder: the Early Markers for Autism (EMA) study: a case-control study. J Neuroinflammation 2014; 11: 113.
Goines PE, Croen LA, Braunschweig D, Yoshida CK, Grether J, Hansen R et al. Increased midgestational IFN-gamma, IL-4 and IL-5 in women bearing a child with autism: a case-control study. Mol Autism 2011; 2: 13.
Nakae S, Suto H, Berry GJ, Galli SJ . Mast cell-derived TNF can promote Th17 cell-dependent neutrophil recruitment in ovalbumin-challenged OTII mice. Blood 2007; 109: 3640–3648.
Ye CJ, Feng T, Kwon HK, Raj T, Wilson MT, Asinovski N et al. Intersection of population variation and autoimmunity genetics in human T cell activation. Science 2014; 345: 1254665.
Chen X, Churchill MJ, Nagar KK, Tailor YH, Chu T, Rush BS et al. IL-17 producing mast cells promote the expansion of myeloid-derived suppressor cells in a mouse allergy model of colorectal cancer. Oncotarget 2015; 6: 32966–32979.
Al-Ayadhi LY, Mostafa GA . Elevated serum levels of interleukin-17 A in children with autism. J Neuroinflammation 2012; 9: 158.
Akintunde ME, Rose M, Krakowiak P, Heuer L, Ashwood P, Hansen R et al. Increased production of IL-17 in children with autism spectrum disorders and co-morbid asthma. J Neuroimmunol 2015; 286: 33–41.
Choi GB, Yim YS, Wong H, Kim S, Kim H, Kim SV et al. The maternal interleukin-17a pathway in mice promotes autismlike phenotypes in offspring. Science 2016; pii: aad0314.
Hsiao EY, McBride SW, Chow J, Mazmanian SK, Patterson PH . Modeling an autism risk factor in mice leads to permanent immune dysregulation. Proc Natl Acad Sci USA 2012; 109: 12776–12781.
Smith SE, Li J, Garbett K, Mirnics K, Patterson PH . Maternal immune activation alters fetal brain development through interleukin-6. J Neurosci 2007; 27: 10695–10702.
Huang M, Pang X, Karalis K, Theoharides TC . Stress-induced interleukin-6 release in mice is mast cell-dependent and more pronounced in Apolipoprotein E knockout mice. Cardiovasc Res 2003; 59: 241–249.
Kandere-Grzybowska K, Letourneau R, Kempuraj D, Donelan J, Poplawski S, Boucher W et al. IL-1 induces vesicular secretion of IL-6 without degranulation from human mast cells. J Immunol 2003; 171: 4830–4836.
Theoharides TC, Boucher W, Spear K . Serum interleukin-6 reflects disease severity and osteoporosis in mastocytosis patients. Int Arch Allergy Immunol 2002; 128: 344–350.
Brockow K, Akin C, Huber M, Metcalfe DD . IL-6 levels predict disease variant and extent of organ involvement in patients with mastocytosis. Clin Immunol 2005; 115: 216–223.
Mayado A, Teodosio C, Garcia-Montero AC, Matito A, Rodriguez-Caballero A, Morgado JM et al. Increased IL6 plasma levels in indolent systemic mastocytosis patients are associated with high risk of disease progression. Leukemia 2015; 30: 124–130.
Abdallah MW, Larsen N, Grove J, Norgaard-Pedersen B, Thorsen P, Mortensen EL et al. Amniotic fluid chemokines and autism spectrum disorders: an exploratory study utilizing a Danish Historic Birth Cohort. Brain Behav Immun 2012; 26: 170–176.
Zhang B, Weng Z, Sismanopoulos N, Asadi S, Therianou A, Alysandratos KD et al. Mitochondria distinguish granule-stored from de novo synthesized tumor necrosis factor secretion in human mast cells. Int Arch Allergy Immunol 2012; 159: 23–32.
Cohly HH, Panja A . Immunological findings in autism. Int Rev Neurobiol 2005; 71: 317–341.
Gomez G, Ramirez CD, Rivera J, Patel M, Norozian F, Wright HV et al. TGF-beta1 inhibits mast cell FcåRI expression. J Immunol 2005; 174: 5987–5993.
Kashyap M, Bailey DP, Gomez G, Rivera J, Huff TF, Ryan JJ . TGF-beta1 inhibits late-stage mast cell maturation. Exp Hematol 2005; 33: 1281–1291.
Jyonouchi H, Sun S, Le H . Proinflammatory and regulatory cytokine production associated with innate and adaptive immune responses in children with autism spectrum disorders and developmental regression. J Neuroimmunol 2001; 120: 170–179.
Naik US, Gangadharan C, Abbagani K, Nagalla B, Dasari N, Manna SK . A study of nuclear transcription factor-kappa B in childhood autism. PLoS One 2011; 6: e19488.
Young AM, Campbell E, Lynch S, Suckling J, Powis SJ . Aberrant NF-kappaB expression in autism spectrum condition: a mechanism for neuroinflammation. Front Psychiatry 2011; 2: 27.
Ziats MN, Rennert OM . Expression profiling of autism candidate genes during human brain development implicates central immune signaling pathways. PLoS One 2011; 6: e24691.
Nelissen S, Lemmens E, Geurts N, Kramer P, Maurer M, Hendriks J et al. The role of mast cells in neuroinflammation. Acta Neuropathol 2013; 125: 637–650.
Shemer A, Erny D, Jung S, Prinz M . Microglia plasticity during health and disease: an immunological perspective. Trends Immunol 2015; 36: 614–624.
Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P et al. Synaptic pruning by microglia is necessary for normal brain development. Science 2011; 333: 1456–1458.
Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mardinly AR, Yamasaki R et al. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 2012; 74: 691–705.
Vargas DL, Nascimbene C, Krishnan C, Zimmerman AW, Pardo CA . Neuroglial activation and neuroinflammation in the brain of patients with autism. Ann Neurol 2005; 57: 67–81.
Morgan JT, Chana G, Pardo CA, Achim C, Semendeferi K, Buckwalter J et al. Microglial activation and increased microglial density observed in the dorsolateral prefrontal cortex in autism. Biol Psychiatry 2010; 68: 368–376.
Morgan JT, Barger N, Amaral DG, Schumann CM . Stereological study of amygdala glial populations in adolescents and adults with autism spectrum disorder. PLoS One 2014; 9: e110356.
Gupta S, Ellis SE, Ashar FN, Moes A, Bader JS, Zhan J et al. Transcriptome analysis reveals dysregulation of innate immune response genes and neuronal activity-dependent genes in autism. Nat Commun 2014; 5: 5748.
Takano T . Role of microglia in autism: recent advances. Dev Neurosci 2015; 37: 195–202.
Koyama R, Ikegaya Y . Microglia in the pathogenesis of autism spectrum disorders. Neurosci Res 2015; 100: 1–5.
Wang W, Ji P, Riopelle RJ, Dow KE . Functional expression of corticotropin-releasing hormone (CRH) receptor 1 in cultured rat microglia. J Neurochem 2002; 80: 287–294.
Martin S, Vincent JP, Mazella J . Involvement of the neurotensin receptor-3 in the neurotensin-induced migration of human microglia. J Neurosci 2003; 23: 1198–1205.
Nykjaer A, Willnow TE . Sortilin: a receptor to regulate neuronal viability and function. Trends Neurosci 2012; 35: 261–270.
Buttenschon HN, Demontis D, Kaas M, Elfving B, Molgaard S, Gustafsen C et al. Increased serum levels of sortilin are associated with depression and correlated with BDNF and VEGF. Transl Psychiatry 2015; 5: e677.
Ghanizadeh A . Targeting neurotensin as a potential novel approach for the treatment of autism. J Neuroinflammation 2010; 7: 58.
Patel AB, Tsilioni I, Theoharides TC . Neurotensin and corticotropin-releasing hormone synergistically activate human microglia through mammalian target ofrapamycin complex 1 (mTORC1), poster # A300 American Societies for Experimental Biology: Bethesda, MD, 2015.
Lee dY . Roles of mTOR signaling in brain development. Exp Neurobiol 2015; 24: 177–185.
Lv JW, Cheng TL, Qiu ZL, Zhou WH . Role of the PTEN signaling pathway in autism spectrum disorder. Neurosci Bull 2013; 29: 773–778.
Tee AR, Sampson JR, Pal DK, Bateman JM . The role of mTOR signalling in neurogenesis, insights from tuberous sclerosis complex. Semin Cell Dev Biol 2016; 52: 12–20.
Kim MS, Kuehn HS, Metcalfe DD, Gilfillan AM . Activation and function of the mTORC1 pathway in mast cells. J Immunol 2008; 180: 4586–4595.
Dong H, Zhang W, Zeng X, Hu G, Zhang H, He S et al. Histamine induces upregulated expression of histamine receptors and increases release of inflammatory mediators from microglia. Mol Neurobiol 2014; 49: 1487–1500.
Zhang S, Zeng X, Yang H, Hu G, He S . Mast cell tryptase induces microglia activation via protease-activated receptor 2 signaling. Cell Physiol Biochem 2012; 29: 931–940.
Skaper SD, Facci L, Giusti P . Neuroinflammation, microglia and mast cells in the pathophysiology of neurocognitive disorders: a review. CNS Neurol Disord Drug Targets 2014; 13: 1654–1666.
Dong H, Zhang X, Wang Y, Zhou X, Qian Y, Zhang S . Suppression of brain mast cells degranulation inhibits microglial activation and central nervous system inflammation. Mol Neurobiol 2016 (in press).
Wong AY, Hsia Y, Chan EW, Murphy DG, Simonoff E, Buitelaar JK et al. The variation of psychopharmacological prescription rates for people with autism spectrum disorder (ASD) in 30 Countries. Autism Res 2014; 7: 543–554.
Spencer D, Marshall J, Post B, Kulakodlu M, Newschaffer C, Dennen T et al. Psychotropic medication use and polypharmacy in children with autism spectrum disorders. Pediatrics 2013; 132: 833–840.
Schubart JR, Camacho F, Leslie D . Psychotropic medication trends among children and adolescents with autism spectrum disorder in the Medicaid program. Autism 2014; 18: 631–637.
Young NJ, Findling RL . An update on pharmacotherapy for autism spectrum disorder in children and adolescents. Curr Opin Psychiatry 2015; 28: 91–101.
Lake JK, Weiss JA, Dergal J, Lunsky Y . Child, parent, and service predictors of psychotropic polypharmacy among adolescents and young adults with an autism spectrum disorder. J Child Adolesc Psychopharmacol 2014; 24: 486–493.
Theoharides TC, Asadi S . Unwanted interactions among psychotropic drugs and other treatments for autism spectrum disorders. J Clin Psychopharmacol 2012; 32: 437–440.
Ecker C, Spooren W, Murphy D . Developing new pharmacotherapies for autism. J Intern Med 2013; 274: 308–320.
Chez MG, Guido-Estrada N . Immune therapy in autism: historical experience and future directions with immunomodulatory therapy. Neurotherapeutics 2010; 7: 293–301.
Liew Z, Ritz B, Virk J, Olsen J . Maternal use of acetaminophen during pregnancy and risk of autism spectrum disorders in childhood: a Danish national birth cohort study. Autism Res 2015 (in press).
Curran LK, Newschaffer CJ, Lee LC, Crawford SO, Johnston MV, Zimmerman AW . Behaviors associated with fever in children with autism spectrum disorders. Pediatrics 2007; 120: e1386–e1392.
Gupta S, Samra D, Agrawal S . Adaptive and innate immune responses in autism: rationale for therapeutic use of intravenous immunoglobulin. J Clin Immunol 2010; 30 (Suppl 1): S90–S96.
Wong PH, White KM . Impact of immunoglobulintherapy in pediatric disease: a review of immune mechanisms. Clin Rev Allergy Immunol 2015 (in press).
Plioplys AV . Intravenous immunoglobulin treatment of children with autism. J Child Neurol 1998; 13: 79–82.
Gupta S, Aggarwal S, Heads C . Dysregulated immune system in children with autism: beneficial effects of intravenous immune globulin on autistic characteristics. J Autism Dev Disord 1996; 26: 439–452.
Giudice-Asch G, Simon L, Schmeidler J, Cunningham-Rundles C, Hollander E . Brief report: a pilot open clinical trial of intravenous immunoglobulin in childhood autism. J Autism Dev Disord 1999; 29: 157–160.
Melamed I, Gupta S, Bobbitt MS, Gilanders K, Hyland N, Moy JN . Efficacy and Safety of Gammaplex® 5% in Children and AdolescentsAnchor with Primary Immunodeficiency Diseases. IgNS Annual Conference Proceedings, 2015; 228–36.
Heuer LS, Rose M, Ashwood P, Van de Water J . Decreased levels of total immunoglobulin in children with autism are not a result of B cell dysfunction. J Neuroimmunol 2012; 251: 94–102.
Siniscalco D, Bradstreet JJ, Cirillo A, Antonucci N . The in vitro GcMAF effects on endocannabinoid system transcriptionomics, receptor formation, and cell activity of autism-derived macrophages. J Neuroinflammation 2014; 11: 78.
Singh K, Connors SL, Macklin EA, Smith KD, Fahey JW, Talalay P et al. Sulforaphane treatment of autism spectrum disorder (ASD). Proc Natl Acad Sci USA 2014; 111: 15550–15555.
King BH, Dukes K, Donnelly CL, Sikich L, McCracken JT, Scahill L et al. Baseline factors predicting placebo response to treatment in children and adolescents with autism spectrum disorders: a multisite randomized clinical trial. JAMA Pediatr 2013; 167: 1045–1052.
Hardan AY, Fung LK, Libove RA, Obukhanych TV, Nair S, Herzenberg LA et al. A randomized controlled pilot trial of oral N-acetylcysteine in children with autism. Biol Psychiatry 2012; 71: 956–961.
Ghanizadeh A, Moghimi-Sarani E . A randomized double blind placebo controlled clinical trial of N-Acetylcysteine added to risperidone for treating autistic disorders. BMC Psychiatry 2013; 13: 196.
Deepmala, Slattery J, Kumar N, Delhey L, Berk M, Dean O et al. Clinical trials of N-acetylcysteine in psychiatry and neurology: A systematic review. Neurosci Biobehav Rev 2015; 55: 294–321.
Nikoo M, Radnia H, Farokhnia M, Mohammadi MR, Akhondzadeh S . N-acetylcysteine as an adjunctive therapy to risperidone for treatment of irritability in autism: a randomized, double-blind, placebo-controlled clinical trial of efficacy and safety. Clin Neuropharmacol 2015; 38: 11–17.
Taliou A, Zintzaras E, Lykouras L, Francis K . An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clin Ther 2013; 35: 592–602.
Theoharides TC, Asadi S, Panagiotidou S . A case series of a luteolin formulation (Neuroprotek®) in children with autism spectrum disorders. Intl J Immunopathol Pharmacol 2012; 25: 317–323.
Theoharides TC, Stewart JM, Hatziagelaki E, Kolaitis G . Brain "fog," inflammation and obesity: key aspects of neuropsychiatric disorders improved by luteolin. Front Neurosci 2015; 9: 225.
Middleton EJ, Kandaswami C, Theoharides TC . The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease and cancer. Pharmacol Rev 2000; 52: 673–751.
Jang SW, Liu X, Yepes M, Shepherd KR, Miller GW, Liu Y et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc Natl Acad Sci USA 2010; 107: 2687–2692.
Scattoni ML, Martire A, Cartocci G, Ferrante A, Ricceri L . Reduced social interaction, behavioural flexibility and BDNF signalling in the BTBR T+ tf/J strain, a mouse model of autism. Behav Brain Res 2013; 251: 35–40.
Johnson RA, Lam M, Punzo AM, Li H, Lin BR, Ye K et al. 7,8-dihydroxyflavone exhibits therapeutic efficacy in a mouse model of Rett syndrome. J Appl Physiol 2012; 112: 704–710.
Theoharides T, Athanassiou M, Panagiotidou S, Doyle R . Dysregulated brain immunity and neurotrophin signaling in Rett syndrome and autism spectrum disorders. J Neuroimmunol 2015; 279: 33–38.
Kimata M, Shichijo M, Miura T, Serizawa I, Inagaki N, Nagai H . Effects of luteolin, quercetin and baicalein on immunoglobulin E-mediated mediator release from human cultured mast cells. Clin Exp Allergy 2000; 30: 501–508.
Kempuraj D, Madhappan B, Christodoulou S, Boucher W, Cao J, Papadopoulou N et al. Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. Br J Pharmacol 2005; 145: 934–944.
Weng Z, Patel A, Panagiotidou S, Theoharidess TC . The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J Allergy Clin Immunol 2014; 14: 1044–1052.
Dirscherl K, Karlstetter M, Ebert S, Kraus D, Hlawatsch J, Walczak Y et al. Luteolin triggers global changes in the microglial transcriptome leading to a unique anti-inflammatory and neuroprotective phenotype. J Neuroinflammation 2010; 7: 3.
Jang S, Kelley KW, Johnson RW . Luteolin reduces IL-6 production in microglia by inhibiting JNK phosphorylation and activation of AP-1. Proc Natl Acad Sci USA 2008; 105: 7534–7539.
Chen HQ, Jin ZY, Wang XJ, Xu XM, Deng L, Zhao JW . Luteolin protects dopaminergic neurons from inflammation-induced injury through inhibition of microglial activation. Neurosci Lett 2008; 448: 175–179.
Parker-Athill E, Luo D, Bailey A, Giunta B, Tian J, Shytle RD et al. Flavonoids, a prenatal prophylaxis via targeting JAK2/STAT3 signaling to oppose IL-6/MIA associated autism. J Neuroimmunol 2009; 217: 20–27.
Harwood M, nielewska-Nikiel B, Borzelleca JF, Flamm GW, Williams GM, Lines TC . A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem Toxicol 2007; 45: 2179–2205.
Theoharides TC, Conti P, Economu M . Brain inflammation, neuropsychiatric disorders, and immunoendocrine effects of luteolin. J Clin Psychopharmacol 2014; 34: 187–189.
Jones QR, Warford J, Rupasinghe HP, Robertson GS . Target-based selection of flavonoids for neurodegenerative disorders. Trends Pharmacol Sci 2012; 33: 602–610.
Acknowledgements
Aspects of our work discussed were funded in part by the Autism Research Institute, the National Autism Association, Safe Minds, the Nancy Lurie Marks Foundation and The Jane Botsford Johnson Foundation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
TCT has been awarded US Patents No 8268365; 9050275 and 9176146 covering treatment of brain inflammation and of ASD. He has developed the luteolin-containing dietary supplements BrainGain, NeuroProtek and NeuroProtek-low phenol. The remaining authors declare no conflicts of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Theoharides, T., Tsilioni, I., Patel, A. et al. Atopic diseases and inflammation of the brain in the pathogenesis of autism spectrum disorders. Transl Psychiatry 6, e844 (2016). https://doi.org/10.1038/tp.2016.77
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/tp.2016.77
This article is cited by
-
Causal relationships between atopic dermatitis and psychiatric disorders: a bidirectional two-sample Mendelian randomization study
BMC Psychiatry (2024)
-
Exploring sulforaphane as neurotherapeutic: targeting Nrf2-Keap & Nf-Kb pathway crosstalk in ASD
Metabolic Brain Disease (2023)
-
Neuroinflammation in autism spectrum disorders: potential target for mesenchymal stem cell-based therapy
The Egyptian Journal of Neurology, Psychiatry and Neurosurgery (2022)
-
Eczema and related atopic diseases are associated with increased symptom severity in children with autism spectrum disorder
Translational Psychiatry (2022)
-
Randomized Controlled Trial of Omega-3 and -6 Fatty Acid Supplementation to Reduce Inflammatory Markers in Children with Autism Spectrum Disorder
Journal of Autism and Developmental Disorders (2022)
