
Abstract
α-Amylases are glycoside hydrolase enzymes that act on the α(1→4) glycosidic linkages in glycogen, starch, and related α-glucans, and are ubiquitously present in Nature. Most α-amylases have been classified in glycoside hydrolase family 13 with a typical (β/α)8-barrel containing two aspartic acid and one glutamic acid residue that play an essential role in catalysis. An atypical α-amylase (BmaN1) with only two of the three invariant catalytic residues present was isolated from Bacillus megaterium strain NL3, a bacterial isolate from a sea anemone of Kakaban landlocked marine lake, Derawan Island, Indonesia. In BmaN1 the third residue, the aspartic acid that acts as the transition state stabilizer, was replaced by a histidine. Three-dimensional structure modeling of the BmaN1 amino acid sequence confirmed the aberrant catalytic triad. Glucose and maltose were found as products of the action of the novel α-amylase on soluble starch, demonstrating that it is active in spite of the peculiar catalytic triad. This novel BmaN1 α-amylase is part of a group of α-amylases that all have this atypical catalytic triad, consisting of aspartic acid, glutamic acid and histidine. Phylogenetic analysis showed that this group of α-amylases comprises a new subfamily of the glycoside hydrolase family 13.
Similar content being viewed by others
Introduction
α-Amylases are ubiquitously present in nature. They act on the α(1→4) glycosidic linkages in glycogen, starch, and related α-glucans and thereby play an important role in the digestion of starch in humans, plants and microorganisms1,2,3. Most α-amylases belong to glycoside hydrolase (GH) family 134, constituting 30 different reaction and product specificities including, glycoside hydrolases (EC 3.2.1.x), glucanotransferases (EC 2.4.1.x and EC 2.4.99.16), and isomerases (EC 5.4.99.x), all sharing a conserved structural scaffold4,5. The crystal structure of Taka α-amylase A from Aspergillus oryzae (TAA), the first experimentally determined three-dimensional (3D) structure of α–amylase6, revealed that α-amylases have three characteristic domains: A, B, and C6,7. The A domain containing the catalytic residues is the most conserved domain, with a typical (β/α)8- or TIM-barrel comprised of eight stranded parallel β-sheet surrounded by eight α-helices. Domain B is inserted between the third β-strand and the third α-helix of the (β/α)8-barrel and varies in length and structure. The C domain folds into eight antiparallel β-strands, is connected to the A domain by loops and seems to be an independent domain with unknown function5,8. Despite low similarity between the amino acid sequences of α-amylases from animals, plants, and microorganisms, the GH13 enzymes share seven highly conserved regions7 that are involved in the formation of the catalytic site. The α-amylase active site is located in an open cavity between the A and B domains, and contains the invariably carboxylic acid Asp206, Glu230 and Asp297 (TAA numbering) being essential for catalysis, acting as the nucleophile, and as the general acid/base and transition state stabilizer, respectively6.
Twenty years ago, several α-amylases and related enzymes composed of a (β/α)7-barrel (an irregular TIM-barrel domain) were classified into the family GH57; more recently also the family GH119 was established9,10. Both of these enzyme families are at present considerably smaller than GH13 and only few members have been characterized in detail11. The first determined 3D structure of GH57 was that of the 4-α-glucanotransferase from Thermococcus litoralis (TLGT). X-ray crystallography supported by site-directed mutagenesis of TLGT revealed that it has two catalytic residues, Glu123 and Asp214, as the catalytic nucleophile and the general acid/base, respectively12. No 3D structure is currently available for GH119 members. In addition to the structural differences between GH13 and GH57-GH119 family members, there are also distinctive conserved regions between these families9. The GH57 and GH119 families share five conserved sequence regions13.
Several microbial strains isolated from a unique land-locked marine lake located in Kakaban island, part of the Derawan Islands, East Kalimantan, Indonesia, were screened for the production of α-amylases. The lake originally was the lagoon of an atoll, formed by corals over a period of two million years. As a result of movements in the earth’s crust the coral reef was raised above the sea level, trapping 5 km2 of seawater within a 50 meter high ridge, effectively creating a land-locked marine lake14. One of the isolates, Bacillus megaterium NL3, contained an active GH13 α-amylase with only the general acid/base residue (Glu231) at the conserved position. Amino acid sequence alignments and 3D homology modeling showed that the nucleophile may be shifted one position downstream (Asp203) and that the transition state stabilizing residue is not the canonical Asp but a His residue (His294). Phylogenetic analysis clustered this new α-amylase and its homologs, which also possess the incomplete GH13 catalytic machinery, as a separate branch in family GH13, representing a novel subfamily.
Results and Discussion
Screening of Kakaban lake isolates producing extracellular amylases
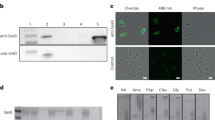
Eight of twenty bacterial isolates from Kakaban landlocked marine lake tested positive for the hydrolysis of starch by producing a clear halo around their colonies on red-dyed amylopectin agar plates. Isolate NL3 showed the largest clearing zone, indicating a relatively high α-amylase activity and was selected for further study. 16S rDNA sequence analysis showed that strain NL3 was most closely related to Bacillus megaterium. This result was in agreement with biochemical and physiological properties (data not shown) and hence the selected isolate was designated as B. megaterium NL3. The culture medium of strain NL3 showed activity towards soluble starch. The 50–80% ammonium sulphate precipitate of the culture medium gave a single protein band with molecular mass of approximately 55 kDa on SDS-PAGE in combination with activity staining with soluble starch (data not shown).
Molecular identification of the NL3 amylase
Using degenerate α-amylase specific primers and inverse PCR, a DNA fragment of 2.3 kb was obtained from genomic DNA of strain NL3. Analysis of the nucleotide sequence of this fragment showed that an open reading frame of 1515 bp with clear α-amylase sequence similarity was present. This gene was designated as bmaN1. A putative ribosomal binding site (RBS) corresponding to the AGGAGG sequence located 12 nucleotides upstream of the start codon was predicted. A probable catabolite responsive element (CRE) was found together with possible −10 (TATAAT) and −35 (TTAACA) regions. The CRE sequence showed only one mismatch in the last position when compared to the consensus sequence (TGT/AAANCGNTNA/TCA)15. The BmaN1 polypeptide deduced is 505 amino acid residues in length with a clear putative signal peptide sequence of 23 residues preceding the mature enzyme, as predicted by SignalP 4.0 Server16. The molecular weight and pI of BmaN1 were predicted using ExPASy server (http://web.expasy.org/protparam) as 56934 Da and 9.05, respectively. The full-length DNA sequence of the putative α-amylase gene of B. megaterium NL3, bmaN1, has been deposited in the GenBank database17 under the accession no. AGT45938.
In silico analysis of BmaN1 and its homologues
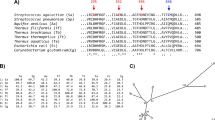
The BLAST search using the BmaN1 amino acid sequence as a query resulted in retrieving of more than 30 highly similar sequences of putative α-amylases (Fig. 1) some of which have already been classified in the family GH1318. Although all of them possess variations in the three residues forming the family GH13 catalytic machinery, it is possible to divide them into two groups: (i) the first, larger group (Nos 1–27 in Fig. 1) with Lys202 and His294 in the positions of the catalytic nucleophile and transition state stabilizer, respectively (instead of normally occurring aspartates); and (ii) a second, smaller group (Nos 28–34 in Fig. 1) exhibiting substitutions in positions of the entire catalytic triad, but rather without an obvious regularity (Fig. 2). While the sequences of the members of the former group are almost identical to BmaN1, those of the latter one are slightly different (Fig. 2). Interestingly, there is a strictly conserved aspartic acid residue succeeding the “strange” lysine, which corresponds with the position of the catalytic nucleophile (Fig. 2). The sequences of both these groups, proposed here to constitute a novel GH13 subfamily xy around the α-amylase from B. megaterium BmaN1, are all highly similar to those of α-amylases around the α-amylase from B. aquimaris BaqA (Nos 35–39 in Fig. 1) suggested recently to define also a new and independent GH13 subfamily xx19. The main difference between the α-amylases around the BmaN1 and those around BaqA is that the BaqA α-amylase and the members of its subfamily possess the complete catalytic machinery (Fig. 2) characteristic for the α-amylase family GH137. The other feature of interest is the presence of a tryptophan pair in both BmaN1 and BaqA (Fig. 2) between the CSR-V (loop 3) and CSR-II (strand β4), located in the helix α3 of the catalytic (β/α)8-barrel19.

aThe list involves: (i) the members of the newly proposed GH13 subfamily xy represented by the “α-amylase” from Bacillus megaterium BmaN1 (Nos 1–27) and its closely related homologues (Nos 28–34) - probably intermediates between BmaN1 and the α-amylase from Bacillus aquimaris BaqA - caught by BLAST; (ii) the members of the recently proposed GH13 subfamily xx19 represented by the BaqA (Nos 35–39); (iii) representatives of the individual GH13 subfamilies with the specificity of α-amylases - subfamilies GH13_1, 5, 6, 7, 15, 19, 24, 27, 28, 32, 36 and 37 (Nos 40–63); and (iv) the currently unassigned cyclomaltodextrinase (GH13_??; No. 64); for details, see the Materials and methods section. b,cAccession numbers from the UniProt (UniParc) and GenBank sequence databases, respectively. dThe length of the entire amino acid sequence of the protein. eThe length of the polypeptide chain spanning the segment from the beginning of the CSR-VI (strand β2) to the end of the CSR-VII (strand β8). fThe GH13 subfamily (if available).

The two consecutive tryptophans characteristic for the novel α-amylase subfamily are also shown. Colour code for the selected residues: W, yellow; F, Y - blue; V, L, I - green; D, E - red; R, K - cyan; H - brown; C - magenta; G, P - black. The positions of the three catalytic residues are boxed and signified by asterisks under the alignment. The label of the protein source consists of the name of the organisms and the UniProt (UniParc) accession number. The number at the beginning of the protein source label indicates the number of known (already established) GH13 subfamily. For the newly proposed BmaN1 GH13 subfamily, the label “xy” is used; similarly (“xx”) for until now non-defined subfamily around the BaqA. The alignment of all 64 enzymes spanning the sequence segment from the beginning of the strand β2 (CSR-VI) to the end of the strand β8 (CSR-VII) is shown in Fig. S2.
In addition to the incomplete catalytic machinery mentioned above, the most striking differences of BmaN1 and its close homologues discriminating them from other well-established GH13 subfamilies with the α-amylase specificity (Fig. 2) are the presence of a glutamic acid instead of aspartate at the beginning of the CSR-I (strand β3) and in addition the position of the histidine in the middle of the CSR-V (loop3), a position usually occupied by aspartic acid20. With regard to alignment of the representative α-amylases studied here (Fig. 1), its substantial part covering almost the entire (β/α)8-barrel including domain B clearly document a very close homology of both eventual GH13 subfamilies, i.e. BmaN1 and BaqA. All these sequences (Nos 1–39 in Fig. 1) go well together exhibiting their own pattern of the alignment in comparison to remaining α-amylases that represent well-established GH13 subfamilies (Nos 40–63 in Fig. 1). Of note is also the fact that the small group of putative α-amylases with irregular substitutions in catalytic positions (Nos 28–34 in Fig. 1) exhibits obviously a higher similarity to the BaqA subfamily than to that around BmaN1, especially in domain B (preceding the CSR-V in loop3) as well as in the segment preceding the CSR-IV at strand β7 (Fig. 2). The cyclomaltodextrinase from Flavobacterium sp. No. 9221 was added to the comparison as an interesting example since it was recently found to possess the pair of adjacent tryptophan residues19, typical for both BmaN1 and BaqA (Fig. 2). This is of interest because the cyclomaltodextrinase belongs to GH13 members intermediate between subfamilies of oligo-1,6-glucosidases and neopullulanases22 that are closely related to α-amylases from the subfamily GH13_36 that, however, do not possess the tryptophan pair (Fig. 2).
A topological alignment of BmaN1 and the putative α-amylases of B. megaterium DSM319, Bacillus sp. 278922, B. flexus, B. aryabhattai, B. megaterium WSH-002, and GTA was made (Fig. S1). Almost all β-strands and α-helices of the TIM barrel in domain A and the Greek key motif in domain C are conserved in these α-amylases. A model of the 3D structure of BmaN1 was generated by the PHYRE server23 and visualized by the MacPymol software24 (Fig. 3). The BmaN1 protein displayed 40% homology (100% confidence, 85% sequence coverage) with the X-ray crystal structure of Geobacillus thermoleovorans α-amylase (GTA, PDB code: 4E2O)25 which was used as a template for the modeling. The comparison between the model and the GTA crystal structure revealed that the global topology is almost the same (Fig. 3). The BmaN1 protein model folds into three distinct domains: a central A domain of 366 residues harboring a (β/α)8 barrel, with an irregular loop domain of 37 residues (domain B) connecting the third β-sheets strand and the third α-helix of the barrel. The C domain of 79 residues has an eight-stranded anti-parallel β-sandwich-like fold (Fig. 3). The (β/α)8 barrel is quite similar to the (β/α)8-barrel found in maltogenic amylase from Pseudomonas saccharophila and Bacillus stearothermophilus26, in that there is an additional helix between Aα6 and Aβ7, which is a three-turn helix lying nearly parallel with the Aα6 strand.

(A) BmaN1 model structure, (B) Structure of BmaN1 (orange) superimposed on GTA (grey) structure, (C) Active-site region in a superposition of BmaN1 with GTA including the acarbose bound in its subsites −2 to +2 (white carbon atoms). Active-center residues of BmaN1 (orange) and GTA (grey) are given as stick models and labeled in orange (BmaN1) and black (GTA).
Superposition of acarbose-bound GTA with the BmaN1 model demonstrated that of the three catalytic residues found in GH13 α-amylases, only residue Glu231 of BmaN1 superimposes with the corresponding residue in GTA (Glu246), and presumably is the general acid/base in BmaN1 (Fig. 3). As already concluded from sequence alignments, two of the three catalytic residues are not conserved in BmaN1. Lys202 replaces the catalytic aspartate (Asp217 of GTA); however, Asp203 directly downstream of the lysine is positioned nearby and has its carboxylic acid side chain pointing into the presumed substrate-binding groove (Fig. 3). Furthermore, at the position corresponding to the nucleophile, His294 replaces the transition-state stabilizing aspartate residue (Asp314) found in α-amylases. The absence of any one of the catalytic residues normally causes partial or complete loss of hydrolysis activity27. Remarkably, the mutant α-amylase from Xanthomonas campestris truncated from the C-terminal part of domain B and thus lacking any of the three conserved catalytic residues, still exhibited starch-hydrolyzing activity28, but that observation has never been supported by other examples.
Phylogeny of BmaN1 and other α-amylases
The evolutionary relatedness of the α-amylase from B. megaterium BmaN1, representing all its homologues with lysine and histidine in positions of the catalytic nucleophile and transition state stabilizer, respectively (Fig. 2; Nos 1–27 in Fig. 1), to members of the recently proposed GH13 subfamily around the BaqA (Nos 35–39 in Fig. 1) as well as to representatives of remaining well-established GH13 subfamilies with α-amylase specificity (Nos 40–63 in Fig. 1), is shown in the evolutionary tree (Fig. 4). It is clear that both subfamilies BmaN1 and BaqA are most closely related to each other among all family GH13 α-amylases. Furthermore, a small group of putative α-amylases with irregular substitutions in catalytic positions (Nos 28–34 in Fig. 1) may be considered as an intermediary connection between both BmaN1 and BaqA subfamilies since, despite the lack of complete family GH13 catalytic machinery (similar to BmaN1), they cluster together with representatives of the BaqA subfamily (Fig. 4). One of the most convincing sequence-structural features characteristic for all these α-amylases is the presence of the pair of adjacent tryptophan residues in helix α3 of the catalytic (β/α)8-barrel (Fig. S2)19. Interestingly, the Flavobacterium sp. No. 92 cyclomaltodextrinase (with the tryptophan pair) is positioned in the evolutionary tree between the subfamilies of BmaN1 and BaqA and all other remaining GH13 families with the α-amylase specificity (Fig. 4).

The label of the protein source consists of the name of the organisms and the UniProt (UniParc) accession number preceded by GH13 subfamily indication. The tree is based on the alignment shown in Fig. S2.
BmaN1 encodes an active exo-acting α-amylase
The gene encoding BmaN1 was cloned in vector pMM1525 and this recombinant plasmid was transformed to B. megaterium MS941. A transformant with clear α-amylase activity, as detected on starch plates by iodine staining, was selected and grown in liquid medium. The culture medium was saturated with 50–80% concentrations of ammonium sulphate to purify the BmaN1 α-amylase enzyme. The molecular weight of the partially purified BmaN1 was estimated to be 55 kDa as judged from activity staining after protein renaturing on SDS-PAGE gels (Fig. S3). In contrast, no band was observed in the culture supernatant of B. megaterium MS941 carrying pMM1525 without any insert (Fig. S3). Amylolytic activity of BmaN1 was measured spectrophotometrically by incubating it with soluble starch and measuring the increase in the amount of reducing sugars released over a period of 40 min (Fig. 5). A clear increase in reducing ends was observed, resulting in an activity of 8.4 U/mg of protein. BmaN1 was found to be most active at 55 °C and pH 6.0. The main end products formed from soluble starch were glucose and maltose, indicating an exo-acting mode of action. Minor amounts of longer chain maltooligosaccharides were also found (Fig. 6). This mode of action is very similar to that of the amylase from Bacillus sp. IMD 435 that releases glucose and maltose as the major products on hydrolysis of both soluble starch and raw corn starch29.

1% (w/v) soluble starch and 12.5 μg/mL of the BmaN1 protein were incubated for various time at 55 °C. Each data point represents the means of triplicate experiments.

A reaction mixture containing 1% (w/v) soluble starch, 12.5 μg/mL of BmaN1, 50 mM maleate buffer pH 6.0 in a total volume of 0.1 mL was incubated at 55 °C. G1, glucose; G2, maltose; ≥G3, maltooligosaccharides.
The results presented above indicate that the substitution of an aspartate residue by a histidine, a positively charged amino acid, still gives (some) amylase activity. The reaction mechanism of BmaN1 may be essentially different from the general catalytic reaction mechanism of α-amylases. Further experiments are needed to demonstrate whether the His residue indeed is one of the catalytic residues of α-amylases.
Methods
Materials
All chemicals used were reagent grade and were obtained from either Fermentas (Maryland, USA) or Difco Laboratories (New Jersey, USA).
Bacterial strains, plasmids, and growth conditions
Twenty microbial strains (gift of Prof. Ocky Karna Radjasa of Diponegoro University, Indonesia) that had been isolated from Kakaban landlocked marine lake (Derawan Islands, East Kalimantan, Indonesia) were screened for extracellular α-amylase activity. The isolates were cultured in marine broth (MB) medium containing 0.25% (w/v) yeast extract, and 0.5% (w/v) peptone in filtered sea water (Seaworld, Ancol, Jakarta, Indonesia) at 30 °C. B. megaterium MS941 (MoBiTec, Germany) and Escherichia coli TOP10 were grown at 37 °C in LB medium (1% (w/v) Bacto-tryptone, 1% (w/v) NaCl and 0.5% (w/v) yeast extract). Ampicillin and tetracyclin were used at concentrations of 100 μg/ml and 12 μg/ml, respectively. The medium was autoclaved at 120 °C for 30 min prior to adding the antibiotics. Plasmid pGEM-T (Promega, USA) was used for PCR product cloning, whereas pMM1525 (MoBiTec, Germany) was used as expression vector.
Screening of α-amylase producing bacteria
Bacterial isolates were inoculated on MB agar plates supplemented with 1.0% (v/v) red-dyed amylopectin30 and then incubated at 30 °C for 24 h. The appearance of a clear zone against a red background was indicative for the production of α-amylase activity. The positive isolates were then subjected to a second screening round using MB agar plates containing 1.0% (w/v) potato or wheat starch. A clearing zone around the bacterial colony indicated that the starch was hydrolyzed and thus that the isolate produced extracellular amylase activity.
Bacterial identification
The isolate showing the largest clearing zone on starch-agar plate was identified by 16SrDNA sequencing. Chromosomal DNA was isolated using Wizard Genomic DNA Purification (Promega). The 16S rDNA gene was amplified by PCR using universal primers UniB1 and BactF1 (Supplementary Table 1). The resulted 1.4 kb fragment was sequenced using the dideoxy-chain termination method (Macrogen, South Korea). The bacterial isolate was identified by aligning the 16s rDNA sequences with other known bacteria using NCBI BLASTn (http://www.ncbi.nlm.nih.gov). 16S rDNA gene sequence was submitted to GenBank.
Cloning of the α-amylase-encoding gene and plasmid construction
Two degenerate primers (Table S1) were designed based on the amino acid sequences of the well-conserved regions (region VI-VII) of α-amylases from several Bacilli. The first α-amylase gene fragment was amplified by polymerase chain reactions (PCR), using chromosomal DNA from B. megaterium NL3 as a template and the two degenerate primers. The PCR products were inserted into pGEM-T vector (Promega) and transformed into E. coli TOP10. Plasmid DNA of the transformed E. coli TOP10 was isolated and the nucleotide sequence of the inserted DNA was determined using the dideoxy-chain termination method (Macrogen). The resulting nucleotide sequence was used to design a set of primers, NL3_SP8-invF1 and NL3_SP8-invR1 (Table S1), to amplify parts of α-amylase gene beyond the conserved region. The chromosomal DNA was partially digested with EcoRV and then self-ligated using T4 DNA ligase (Fermentas). Inverse PCR (iPCR) was performed with Dream Taq polymerase (Fermentas) and the primers listed in Supplementary Table 1 using the self-ligated DNA fragment as a template. Analysis of sequence data and sequence similarity searches was performed using the BLAST program of the National Center for Biotechnology Information (NCBI). Primers pMM-NL3-F and pET/MM-NL3-R (Table S1) were used to amplify the complete open reading frame of the α-amylase gene which was designated as bmaN1.
Transformation of B. megaterium
The recombinant plasmid containing the α-amylase gene, pMM1525-bmaN1, was transformed into the expression host, B. megaterium MS941. The transformation procedure was essentially conducted as described by Puyet et al. with some modifications31. A 0.5 ml protoplast suspension was added to a tube containing 5.0 μg DNA and 1.5 ml PEG-P (40% (w/v) PEG6000 in 1x SMMP) for each transformation and incubated for 2 min at room temperature. SMMP medium contains 3.5% (w/v) AB3 (Antibiotic medium no. 3, Difco), 1 M sucrose, 40 mM disodium maleic acid and 40 mM MgCl2 (pH adjusted to 6.5 before autoclaving for 12 min) and prepared freshly before use. To the mixture, 5.0 ml SMMP was added and mixed by rolling the tube carefully. Cells were harvested by centrifugation at 2,700 × g for 10 min at room temperature and the supernatant was poured off immediately. The pellets were resuspended with 0.5 ml SMMP and incubated at 37 °C for 90 min with gentle shaking or rolling of tubes (max. 100 rpm). Then, 50 to 200 μl of cells were added into top agar and mixed gently by rolling the tube. The mixture was poured on a pre-warmed plate of LB containing 12 μg/ml of tetracycline and incubated at 37 °C overnight.
Expression and partial purification of recombinant α-amylase
α-Amylase was produced by growing the B. megaterium harboring recombinant plasmids in 20 ml of LB medium supplemented with 12 μg/ml tetracycline at 37 °C with shaking. The overnight culture was transferred into fresh media and incubated until the 546-nm absorbance reached 0.8–1.0. Subsequently, expression was induced by adding 1% (v/v) xylose, and the culture was incubated at 18 °C with constant shaking at 150 rpm for 24 h. Cells were removed by centrifugation (6000 × g, 10 min) and the resulted supernatant was subjected to ammonium sulphate precipitation at a saturation value up to 80%. The precipitate was dissolved and dialyzed against 50 mM maleate buffer pH 6.0 at 4 °C. This partially purified α-amylase was used for further studies.
Gel electrophoresis and activity staining
SDS-PAGE was carried out as described by Laemmli32 and gels were then stained with Coomassie Brilliant Blue (Bio-Rad). For α-amylase activity test, the protein samples were separated by SDS-PAGE containing 1% soluble starch. After electrophoresis, SDS was removed by washing the gel with water followed by 10 min incubation at room temperature. This was repeated twice. The gels were then immersed in the enzyme reaction buffer (50 mM maleate buffer pH 6.0) for 4.0 h at 55 °C and then stained with KI/I2 solution for 10 min and followed by rinsing with water. The α-amylase activity was detected as a clear zone against a purple background.
Enzyme assay
The amylase activity assay was conducted using the 3,5-dinitrosalicylic acid (DNS) method described by Miller (1955) with a slight modification33. Briefly, the assay was performed by adding 25 μl of enzyme sample into 25 μl of 1% (w/v) soluble starch (Fermentas, USA) in 50 mM of the appropriate buffer and then incubated at 55 °C for 10 min. To the reaction mixture, 50 μl of DNS reagent was added. The absorbance at 500 nm was measured and then the amount of reducing sugar-end was calculated using a glucose standard curve. One unit of α-amylase activity was defined as the amount of enzyme that releases 1 μmol of reducing sugar per min under the assay conditions. The protein concentration was determined using the Lowry method and bovine serum albumin as the standard.
Analysis of sugars
The starch hydrolysis products were analyzed by high-performance liquid chromatography (HPLC). Aliquots of 100 μl of enzyme solution were incubated at 55 °C in the presence of 1% (w/v) soluble starch, maleate buffer 50 mM. After specific time intervals, samples were withdrawn and hydrolysis was stopped by incubation at 90 °C for 10 min. After centrifugation at 12000 × g for 10 min at 4 °C, the products were then analyzed by HPLC (Aminex® HPX-87H system). The separated hydrolysis products were identified by calculating based on peak areas compared to standard glucose, maltose, and purified maltooligosaccharide (Sigma).
Bioinformatics
The sequences eventually forming the new GH13 subfamily xy were collected based on protein BLAST34 searches against the non-redundant database using the entire amino acid sequence of Bacillus megaterium NL3 α-amylase BmaN1 (UniProt accession No.: T1SIF2) as well as on previous bioinformatics analyses when the BLAST was performed with the Bacillus aquimaris α-amylase BaqA19,35. The main criterion applied for the selection was the lack of at least one residue from the catalytic triad characteristic for the α-amylase family GH13. In addition to BmaN1 and its closely related homologues, five experimentally characterized α-amylases from the recently proposed GH13 subfamily around the B. aquimaris α-amylase BaqA19,35 were added. These α-amylases – BaqA from B. aquimaris35, ASKA and ADTA from Anoxybacillus sp.36,37, GTA and GTA-II from Geobacillus thermoleovorans25,38,39 – exhibit a high degree of sequence similarity with BmaN1, but possess the complete GH13 catalytic machinery19. The entire set was finally completed by two selected representatives from well-established GH13 subfamilies with the α-amylase specificity, i.e. 1, 5, 6, 7, 15, 19, 24, 27, 28, 32, 36 and 3719 including also the related but until now unclassified cyclomaltodextrinase from Flavobacterium sp. No. 19221 so that the final number of studied enzymes and hypothetical proteins was 64 (Fig. 1).
All 64 GH13 sequences were retrieved from GenBank17 and UniProt40 sequence databases and the set was aligned using the program Clustal-Omega41 available at the European Bioinformatics Institute’s web-site (http://www.ebi.ac.uk/). A subtle manual tuning was done in order to maximize similarities, especially with regard to aligning the individual CSRs. The boundaries of the CSRs were defined based on previous bioinformatics studies19,20. The evolutionary tree was constructed based on the final alignment of the sequence segment corresponding to a 255-residue long region of BmaN1 α-amylase spanning almost the entire catalytic (β/α)8-barrel domain including the domain B from the beginning of the CSR-VI (strand β2; starting with Gly79) to the end of the CSR-VII (strand β8; ending with Ser333). The tree was calculated as a Phylip-tree type using the neighbour-joining clustering and the bootstrapping procedure - the number of bootstrap trials used was 1,00042 implemented in the Clustal-X package43, and then displayed with the program iTOL44.
The 3D structure of BmaN1 was predicted by QuickPhyre structure program server (http://www.sbg.bio.ic.ac.uk/phyre)23. Structural modeling of the BmaN1 was performed based on the crystal structures of α-amylase of G. thermoleovorans [PDB access code: 4E20]. The generated BmaN1 structures were displayed and drawn by MacPymol.
Additional Information
How to cite this article: Sarian, F. D. et al. A new group of glycoside hydrolase family 13 α-amylases with an aberrant catalytic triad. Sci. Rep. 7, 44230; doi: 10.1038/srep44230 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Van der Maarel, M. J. E. C., van der Veen, B., Uitdehaag, J. C. M., Leemhuis, H. & Dijkhuizen, L. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol. 94, 137–155 (2002).
Butterworth, P. J., Warren, F. J. & Ellis, P. R. Human α-amylase and starch digestion: An interesting marriage. Starch - Stärke 63, 395–405 (2011).
Zeeman, S. C., Kossmann, J. & Smith, A. M. Starch: its metabolism, evolution and biotechnological modification in plants. Ann. Rev. Plant Biol. 61, 209–234 (2010).
Cantarel, B. L. et al. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics. Nucleic Acids Res. 37, D233–238 (2009).
MacGregor, E. A., Janeček, Š. & Svensson, B. Relationship of sequence and structure to specificity in the α-amylase family of enzymes. Biochim. Biophys. Acta - Protein Struct. Mol. Enzymol. 1546, 1–20 (2001).
Matsuura, Y., Kusunoki, M., Harada, W. & Kakudo, M. Structure and possible catalytic residues of Taka-amylase A. J. Biochem. 95, 697–702 (1984).
Janeček, Š., Svensson, B. & MacGregor, E. A. α-Amylase: an enzyme specificity found in various families of glycoside hydrolases. Cell. Mol. Life Sci. 71, 1149–1170 (2014).
MacGregor, E. A. α-Amylase structure and activity. J. Protein Chem. 7, 399–415 (1988).
Janeček, Š. & Kuchtová, A. In silico identification of catalytic residues and domain fold of the family GH119 sharing the catalytic machinery with the α-amylase family GH57. FEBS Lett. 586, 3360–3366 (2012).
Blesák, K. & Janeček, Š. Two potentially novel amylolytic enzyme specificities in the prokaryotic glycoside hydrolase α-amylase family GH57. Microbiology 159, 2584–2593 (2013).
Henrissat, B. & Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 7, 637–644 (1997).
Imamura, H. et al. Crystal structures of 4-α-glucanotransferase from Thermococcus litoralis and its complex with an inhibitor. J. Biol. Chem. 278, 19378–19386 (2003).
Zona, R., Chang-Pi-Hin, F., O’Donohue, M. J. & Janeček, Š. Bioinformatics of the glycoside hydrolase family 57 and identification of catalytic residues in amylopullulanase from Thermococcus hydrothermalis . Eur. J. Biochem. 271, 2863–2872 (2004).
Radjasa, O. K., Limantara, L. & Sabdono, A. Antibacterial activity of a pigment producing-bacterium associated with Halimeda sp. from island-locked marine lake kakaban, Indonesia. J. Coast. Dev. 12, 100–104 (2019).
Hueck, C. J. & Hillen, W. Catabolite repression in Bacillus subtilis: a global regulatory mechanism for the Gram-positive bacteria? Mol. Microbiol. 15, 395–401 (1995).
Petersen, T. N., Brunak, S., von Heijne, G. & Nielsen, H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Meth 8, 785–786 (2011).
Benson, D. A. et al. GenBank. Nucleic Acids Res. 43, D30–5 (2015).
Lombard, V. et al. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495 (2013).
Janeček, Š., Kuchtová, A. & Petrovičová, S. A novel GH13 subfamily of α-amylases with a pair of tryptophans in the helix α3 of the catalytic TIM-barrel, the LPDlx signature in the conserved sequence region V and a conserved aromatic motif at the C-terminus. Biologia 70, 1284–1294 (2015).
Janeček, Š. How many conserved sequence regions are there in the α-amylase family? Biologia 57 (Suppl. 11), 29–41 (2002).
Fritzsche, H. B., Schwede, T. & Schulz, G. E. Covalent and three-dimensional structure of the cyclodextrinase from Flavobacterium sp. no. 92. Eur. J. Biochem. 270, 2332–2341 (2003).
Majzlová, K., Pukajová, Z. & Janeček, Š. Tracing the evolution of the α-amylase subfamily GH13_36 covering the amylolytic enzymes intermediate between oligo-1,6-glucosidases and neopullulanases. Carbohydr. Res. 367, 48–57 (2013).
Kelley, L. A. et al. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 10, 845–858 (2015).
Schrodinger LLC. The PyMOL molecular graphics system, Version 1.3r1 (2010).
Mok, S.-C. et al. Crystal structure of a compact α-amylase from Geobacillus thermoleovorans . Enzyme Microb. Technol. 53, 46–54 (2013).
Jespersen, H. M., MacGregor, E. A., Sierks, M. R. & Svensson, B. Comparison of the domain-level organization of starch hydrolases and related enzymes. Biochem. J. 280, 51–55 (1991).
Nielsen, J. E. & Borchert, T. V. Protein engineering of bacterial α-amylases. Biochim. Biophys. Acta - Protein Struct. Mol. Enzymol. 1543, 253–274 (2000).
Ke, T. et al. A mutant α-amylase with only part of the catalytic domain and its structural implication. Biotechnol. Lett. 29, 117–122 (2007).
Hamilton, L. M., Kelly, C. T. & Fogarty, W. M. Production and properties of the raw starch-digesting α-amylase of Bacillus sp. IMD 435. Process Biochem. 35, 27–31 (1999).
Jørgensen, S., Vorgias, C. E. & Antranikian, G. Cloning, sequencing, characterization, and expression of an extracellular α-amylase from the hyperthermophilic archaeon Pyrococcus furiosus in Escherichia coli and Bacillus subtilis . J. Biol. Chem. 272, 16335–16342 (1997).
Puyet, A. et al. A simple medium for rapid regeneration of Bacillus subtilis protoplasts transformed with plasmid DNA. FEMS Microbiol. Lett. 40, 1–5 (1987).
Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–685 (1970).
Miller, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426–428 (1959).
Altschul, S. F. et al. Basic local alignment search tool. J. Mol. Biol. 215, 403–10 (1990).
Puspasari, F. et al. Raw starch-degrading α-amylase from Bacillus aquimaris MKSC 6.2: isolation and expression of the gene, bioinformatics and biochemical characterization of the recombinant enzyme. J. Appl. Microbiol. 114, 108–120 (2013).
Chai, Y. Y., Rahman, R. N., Illias, R. M. & Goh, K. M. Cloning and characterization of two new thermostable and alkalitolerant α-amylases from the Anoxybacillus species that produce high levels of maltose. J. Ind. Microbiol. Biotechnol. 39, 731–741 (2012).
Chai, K. P. et al. Crystal structure of Anoxybacillus α-amylase provides insights into maltose binding of a new glycosyl hydrolase subclass. Sci. Rep. 6, 23126 (2016).
Finore, I. et al. Purification, biochemical characterization and gene sequencing of a thermostable raw starch digesting α-amylase from Geobacillus thermoleovorans subsp. stromboliensis subsp. nov. World J. Microbiol. Biotechnol. 27, 2425–2433 (2011).
Mehta, D. & Satyanarayana, T. Domain C of thermostable α-amylase of Geobacillus thermoleovorans mediates raw starch adsorption. Appl. Microbiol. Biotechnol. 98, 4503–4519 (2014).
UniProt Consortium. UniProt: a hub for protein information. Nucleic Acids Res. 43, D204–12 (2015).
Sievers, F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7, 539 (2011).
Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution (N. Y). 39, 783–791 (1985).
Larkin, M. A. et al. Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948 (2007).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL): an online tool for phylogenetic tree display and annotation. Bioinformatics 23, 127–128 (2007).
Acknowledgements
This research was funded by Asahi Glass Foundation, Institut Teknologi Bandung. We thank Prof. F. Meinhardt, University of Munster, Germany, for B. megaterium MS941 strain and pMM1525 plasmid. We are grateful to Dr. Lily Eurwilaichitr and Dr. Sutipa Tanapongpipat for allowing us to do the preliminary experiments on BIOTEC, NSTDA, Thailand. ŠJ thanks the Slovak Grant Agency VEGA – the project No. 2/0150/14 – for financial support.
Author information
Authors and Affiliations
Contributions
F.D.S., I., Z.N., D.N., and O.K.R. planned the experiments on the isolation and characterization of the strain and the isolation of the amylase gene and analysed the results; F.D.S., D.N. and M.M. planned the biochemical analysis of the amylase and analysed the results; F.D.S. performed all these experiments; F.D.S., D.N., S.J., T.P., L.D. and M.M. wrote the manuscript; F.D.S. and T.P. performed the 3D modeling and interpreted the results, F.D.S. and S.J. performed the phylogenetic analysis and interpreted the results; all authors have seen the final version of the manuscript and agree with the content.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sarian, F., Janeček, Š., Pijning, T. et al. A new group of glycoside hydrolase family 13 α-amylases with an aberrant catalytic triad. Sci Rep 7, 44230 (2017). https://doi.org/10.1038/srep44230
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44230
This article is cited by
-
Two newly established and mutually related subfamilies GH13_48 and GH13_49 of the α-amylase family GH13
Applied Microbiology and Biotechnology (2024)
-
Three marine species of the genus Fulvivirga, rich sources of carbohydrate-active enzymes degrading alginate, chitin, laminarin, starch, and xylan
Scientific Reports (2023)
-
In silico investigation of sequences and predicted three-dimensional homology models of thermoalkaliphilic GH13 α-amylases
Biologia (2023)
-
Characterization of an Amylolytic Enzyme from Massilia timonae of the GH13_19 Subfamily with Mixed Maltogenic and CGTase Activity
Applied Biochemistry and Biotechnology (2023)
-
Sequence-structural features and evolution of the α-amylase family GH119 revealed by the in silico analysis of its relatedness to the family GH57
Biologia (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
