
Abstract
This study focused on the mechanisms that fatty acid conjugating strains - Bifidobacterium breve NCIMB 702258 and Bifidobacterium breve DPC 6330 - influence lipid metabolism when ingested with α-linolenic acid (ALA) enriched diet. Four groups of BALB/c mice received ALA enriched diet (3% (w/w)) either alone or in combination with B. breve NCIMB 702258 or B. breve DPC 6330 (109 CFU/day) or unsupplemented control diet for six weeks. The overall n-3 PUFA score was increased in all groups receiving the ALA enriched diet. Hepatic peroxisomal beta oxidation increased following supplementation of the ALA enriched diet with B. breve (P < 0.05) and so the ability of the strains to produce c9t11 conjugated linoleic acid (CLA) was identified in adipose tissue. Furthermore, a strain specific effect of B. breve NCIMB 702258 was found on the endocannabinoid system (ECS). Liver triglycerides (TAG) were reduced following ALA supplementation, compared with unsupplemented controls (P < 0.01) while intervention with B. breve further reduced liver TAG (P < 0.01), compared with the ALA enriched control. These data indicate that the interactions of the gut microbiota with fatty acid metabolism directly affect host health by modulating n-3 PUFA score and the ECS.
Similar content being viewed by others
Introduction
Several studies support a communication axis between the gut microbiota and adipose tissue, which influence the development of metabolic alterations associated with obesity. Conventionally raised mice have long been known to have >40% more fat mass than their isocaloric-fed germ-free counterparts1 and more recently, it was shown that transplantation of the gut microbiota from obese mice to germ-free mice resulted in a greater increase in total body fat mass accumulation compared with those colonised with gut microbiota isolated from lean donors2. The resistance of germ-free mice to high fat diet induced body weight gain and fat mass accumulation has also been described, suggesting that the gut microbiota may promote fat storage3.
Furthermore, the gut microbiota play a role in the low grade inflammation associated with obesity and related metabolic disorders4,5,6,7,8. Gut microbiota-derived lipopolysaccharide (LPS) may represent a cause for the onset and progression of inflammation (metabolic endotoxemia) associated with insulin resistance, type 2 diabetes and high fat diet induced increased fat mass. The endocannabinoid system (ECS) interconnects obesity, inflammation and the gut microbiota in the regulation of energy homeostasis, appetite9 and gut barrier function10 via the microbiota-gut-brain axis. It has been demonstrated that the gut microbiota control ECS activity in the intestine and adipose tissue11,12 through gut microbial regulation of CB1 receptor expression11. The ECS system is composed mainly of the bioactive lipids anandamide (AEA; an N-arachidonoylethanolamine), 2 arachidonoylgycerol (2-AG) (synthesised locally in the gastrointestinal tract), the proteins that regulate their production/degradation and the cannabinoid receptors CB1 and CB2, through which they signal. Obesity is associated with an increase in ECS tone and an altered expression of CB1. An increase in endogenous production and content of AEA in the colon and both visceral and subcutaneous adipose tissue11,12 was blunted by prebiotic treatment11. By altering gut microbiota composition through prebiotic feeding, colonic CB1 mRNA expression is reduced and antibiotic treatment also decreased the expression of the CB1 receptor in the colon. These results correlated with reduced colonic AEA (endogenous CB1 ligand), increased fatty acid amine hydrolase (the main enzyme in degradation of AEA) and reduced plasma LPS11. Therefore, the gut microbiota, the innate immune system and the ECS interact in the development of obesity and related disorders.
We previously demonstrated that fatty acid composition of the host is influenced by microbial metabolism in the gut and that the effects were strain-specific. B. breve NCIMB 702258 and B. breve DPC 6330 are efficient producers of conjugated linoleic acid (CLA), converting up to 65% and 76%, respectively, of linoleic acid to cis-9, trans-11 CLA when grown in 0.5 mg/mL linoleic acid in vitro13,14. We have also demonstrated the ability of these two strains to produce other conjugated fatty acids, including conjugated α-linolenic acid (CNLA), conjugated γ-linoleic acid and conjugated stearidonic acid15. Dietary supplementation with B. breve NCIMB 702258 and B. breve DPC 6330 positively influenced tissue fatty acid profiles in different animal species and models, while also influencing host intestinal microbiota composition16,17,18,19.
These data suggest that dietary supplementation with a commensal bacterium can significantly influence health through the production of bioactive conjugated fatty acids and by increasing tissue concentrations of bioactive long-chain (LC) members of the n-3 PUFA family (eicosapentaenoic acid (EPA; C20:5), docosapentaenoic acid (C22:5) and docosahexaenoic acid (DHA; C22:6). While CLA isomers have demonstrated anti-atherosclerotic20 and anti-diabetogenic21,22 properties, both CLA and CNLA isomers have exhibited anti-carcinogenic23,24,25,26,27 and anti-obesogenic28,29,30,31,32,33 properties. Bioactive EPA, docosapentaenoic acid and DHA can partly inhibit a number of aspects of inflammation including leukocyte chemotaxis, adhesion molecule expression and leukocyte–endothelial adhesive interactions, production of n-6 PUFA derived eicosanoids from arachidonic acid, production of inflammatory cytokines and T-helper 1 lymphocyte reactivity, extensively reviewed34. Thus, interactions between resident gut microbes and dietary derived fatty acids with implications for health have been described35,36,37.
While optimal dietary intakes of n-6:n-3 PUFA should be in the ratio of ~1–4:1; recent shifts in dietary PUFA consumption have established this ratio as ~20:1 in Western diets, favouring n-6 PUFA metabolism38. Considering that n-6 PUFA are precursors of inflammatory eicosanoids (2-series prostaglandins, 2-series thromboxanes and 4-series leukotrienes), it is unsurprising that the incidence of diseases strongly associated with inflammatory processes such as cardiovascular disease (CVD), obesity, inflammatory bowel disease (IBD) and cancer have all been linked to high intakes of n-6:n-3 PUFA34. In contrast, eicosanoids derived from n-3 PUFA (3-series prostaglandins, 3-series thromboxanes and 5-series leukotrienes) are broadly anti-inflammatory, however, the benefits of ALA consumption on cardiovascular health, immune regulation and inflammation have been described as moderately positive39.
The elongation and desaturation of PUFAs, primarily occurs in the liver38, but ALA is considered a poor precursor for DHA due to the efficiency to undergo beta-oxidation in the mitochondria and because biosynthesis of DHA requires a crucial step in the peroxisome for partial beta-oxidation40. However, in specific dietary supplementation, ALA increased tissue concentrations of EPA and DHA17,41,42, in fact, we have previously demonstrated that supplementing an ALA enriched diet with B. breve NCIMB 702258 and B. breve DPC 6330 increased tissue concentrations of EPA, docosapentaenoic acid and DHA, compared with dietary ALA alone17,19, favouring DHA biosynthesis. Flaxseed has thus emerged as an important functional food ingredient, as it is one of the richest sources of ALA (1 tablespoon of flaxseed oil contains ~8 g ALA)43.
Therefore, the aim of this study was to investigate the impact of dietary ALA enrichment with or without fatty acid conjugating microbial supplementation on fat composition and distribution and to investigate the mechanisms through which commensal gut microbes may alter lipid metabolism.
Results
Survival and transit of B. breve DPC 6330 and B. breve NCIMB 702258 in Balb/c mice
Quantification of the numbers of administered B. breve strains in the feces of mice confirmed gastrointestinal transit and survival. Stool recovery of B. breve NCIMB 702258 and B. breve DPC 6330 were approximately 1 × 107 CFU/g feces and 5 × 106 CFU/g feces, respectively, after 1 week of dietary supplementation and remained at similar numbers at weeks 2 and 4. At week 6, there was a decline in the numbers of excreted B. breve strains, with stool recovery of B. breve NCIMB 702258 and B. breve DPC 6330 being 9 × 105 CFU/g feces and 8 × 105 CFU/g feces, respectively. Mice that received the bacterial strains had similar CFU/g after the treatment and importantly, B. breve was not isolated from any of the unsupplemented mice or mice receiving ALA alone.
Dietary supplementation with B. breve NCIMB 702258 and B. breve DPC 6330 reduced liver TAG levels
No differences in food intake, body weight gain, visceral fat mass or serum TAG levels were observed between the groups (Table 1). Analysis of total liver TAG showed that ALA supplementation, either alone (ALA-CON) or in combination with either B. breve strain (ALA + NCIMB 702258/ALA + DPC 6330) resulted in reduced liver TAG levels, compared with the unsupplemented control (CON) (P < 0.01; Table 1). Furthermore, ALA + NCIMB 702258 and ALA + DPC 6330 exhibited reduced liver TAG, compared with ALA-CON (P < 0.01; Table 1).
Effect of dietary enrichment with ALA alone or supplemented with B. breve NCIMB 702258 and B. breve DPC 6330 on tissue fatty acid composition
To investigate the effects of the ALA enriched diet on tissue fatty acid composition, fatty acid profiling (saturated and unsaturated) was performed on liver and epididymal adipose tissue. Following 6 weeks of dietary intervention, all ALA enriched groups had increased caprylic acid (C8:0) in the liver (P < 0.05; Table 2), compared with CON. For unsaturated fatty acids, all ALA enriched groups had increased stearidonic acid (C18:4) in the liver (P < 0.05; Table 2), increased ALA (C18:3n-3) and EPA (C20:5) in both liver (P < 0.05; Table 2) and adipose tissue (P < 0.05; Table 3) and decreased arachidonic acid (C20:4) and oleic acid (C18:1) in adipose tissue (P < 0.05; Table 3), compared with CON.
Comparing the effects of dietary supplementation with ALA + NCIMB 702258 with ALA-CON, there were significant increases in the saturated fatty acids caprylic acid (P < 0.05; Table 2), capric acid (C10:0; P < 0.05; Table 2), lauric acid (C12:0; P < 0.05; Table 2) and myristic acid (C14:0; P < 0.05; Table 2) in liver and increased pentadecyclic acid (C15:0; P < 0.05; Table 3) and stearic acid (C18:0; P < 0.05; Table 3) in adipose tissue. Oleic acid was decreased in adipose tissue (P < 0.05; Table 3), while the PUFAs EPA (P < 0.05; Table 3) and DHA (C22:6; P < 0.05; Table 3) were increased and DHA was increased in liver (P < 0.05; Table 2) in ALA + NCIMB 702258, compared with ALA-CON.
Supplementation of the ALA enriched diet with B. breve DPC 6330 decreased stearic acid in liver (P < 0.05; Table 2) and increased pentadecyclic acid in adipose tissue (P < 0.05; Table 3), compared with ALA-CON. B. breve DPC 6330 also increased hepatic linoleic acid (C18:2; P < 0.05; Table 2) and DHA in the liver (P < 0.05; Table 2) and adipose tissue (P < 0.05; Table 3) and decreased adipose tissue oleic acid (P < 0.05; Table 3) compared with ALA-CON.
Unexpectedly, the n-3 PUFA score was significantly increased in all ALA enriched groups compared with CON, irrespective of microbial supplementation (P < 0.05; Table 2). It was found that hepatic peroxisomal beta oxidation was significantly increased following supplementation of the ALA enriched diet with B. breve (both strains), compared with CON and ALA-CON (P < 0.05; Table 2). This result correlated with significantly reduced hepatic levels of CNLA (P < 0.05) and c9t11CLA (P < 0.05) following supplementation with B. breve compared with CON and ALA-CON (Table 2). Interestingly, CNLA (P < 0.05) and c9t11CLA (P < 0.05) were increased in adipose tissue following supplementation with both B. breve strains compared with CON and ALA-CON (Table 3).
Strain specific effects of B. breve on endocannabinoid and congener levels in liver and adipose tissue
To understand the dietary and microbial impact of ALA and B. breve on the ECS, liver and adipose tissue levels of endocannabinoids (EC) and their congeners were determined. Hepatic levels of AEA, oleoylethanolamide (OEA) and 2-AG were reduced in all ALA enriched dietary groups, compared with CON (P < 0.05; Supplementary Table 1 (displaying significance) and Fig. 1). Furthermore, both B. breve strains reduced AEA and N-palmitoylethanolamide (PEA) levels in liver, compared with ALA-CON (P < 0.05; Supplementary Table 1 and Fig. 1), while B. breve DPC 6330 also reduced hepatic OEA levels, compared with ALA-CON (P < 0.05; Supplementary Table 1 and Fig. 1).

Actual values and significant differences are highlighted in Supplementary Table 1. AEA, N-arachidonoylethanolamide; PEA, N-palmitoylethanolamide; OEA, oleoylethanolamide; 2AG, 2-arachidonoylglycerol; DHEA, docosahexaenoylethanolamide.
B. breve supplementation had a profound effect on EC and congener levels in adipose tissue. Both strains increased AEA and docosahexaenoylethanolamide (DHEA) levels here compared with CON and ALA-CON (P < 0.05; Supplementary Table 1 and Fig. 1). Interestingly, a greater effect of B. breve NCIMB 702258 on adipose tissue EC levels was identified, as this strain also increased PEA (P < 0.05) and OEA (P < 0.05) levels compared with CON and ALA-CON (Supplementary Table 1 and Fig. 1).
Impact on mRNA gene expression levels of enzymes involved in n-3 PUFA metabolism within liver and uptake of fatty acids to liver following dietary enrichment with ALA either alone or supplemented with B. breve NCIMB 702258 or B. breve DPC 6330
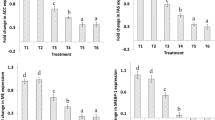
The mRNA expression of enzymes involved in n-3 PUFA metabolism within liver (Fig. 2(A)) and genes involved in the transport and uptake of fatty acids to the liver (Fig. 2(B)) were determined using real-time PCR. An increase in hepatic mRNA expression of Δ-6-desaturase was found in all ALA enriched groups, compared with CON (P < 0.01; Fig. 2(A)). Hepatic mRNA expression of elongation of very long chain fatty acids (ELOVL)-5 was reduced in ALA-CON compared with CON (P < 0.05; Fig. 2(A)). Hepatic mRNA expression of sterol regulatory element binding protein (SREBP)-1c was increased following supplementation with ALA, compared with CON (P < 0.05; Fig. 2(B)). The CON group exhibited increased hepatic mRNA expression of fatty acid synthase (FAS), compared with all ALA enriched groups (P < 0.01; Fig. 2(B)).

Expression in the unsupplemented and ALA enriched diets supplemented with either B. breve NCIMB 702258 or B. breve DPC 6330 is relative to the ALA enriched diet, which was set to 1. Values are means ± SD, represented by vertical bars. Data was analysed by one-way ANOVA, followed by post hoc Tukey’s multiple comparison test. Significant differences are represented by *(P < 0.05), **(P < 0.05), ***(P < 0.05).
Hepatic mRNA expression of ELOVL-5 was elevated in ALA + DPC 6330 (P < 0.05; Fig. 2(A)), compared with ALA-CON. ALA + NCIMB 702258, ALA + DPC 6330 and CON demonstrated significantly reduced hepatic mRNA expression of Δ-5-desaturase, compared with ALA-CON (P < 0.01; Fig. 2(A)).
Both ALA + NCIMB 702258 and ALA + DPC 6330 exhibited decreased hepatic mRNA expression of CD-36, compared with ALA-CON and CON (P < 0.05; Fig. 2(B)). In addition, ALA + NCIMB 702258 and ALA + DPC 6330 had decreased hepatic mRNA expression of fatty acid binding protein (FABP)-1, compared with ALA-CON (P < 0.05; Fig. 2(B)). Hepatic mRNA expression of SREBP-1c was increased in ALA + DPC 6330, compared with CON (P < 0.05; Fig. 2(B)).
Impact on mRNA gene expression levels of enzymes involved in the transport and uptake of fatty acids to the ileum following dietary enrichment with ALA either alone or supplemented with B. breve NCIMB 702258 or B. breve DPC 6330
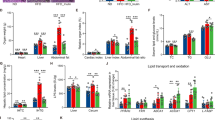
ALA + NCIMB 702258 and ALA + DPC 6330 decreased ileal mRNA gene expression of FABP-2, compared with ALA-CON (P < 0.05; Fig. 3). Although not significant, the expression of FABP-1 and CD-36 were increased in ALA + DPC 6330 (Fig. 3). The expression of fatty acid transport protein (FATP)-4 and diacylglycerol O-acyltransferase (DGAT)-2 were unchanged between all groups (Fig. 3).

Expression in the unsupplemented diet and ALA enriched diets supplemented with either B. breve NCIMB 702258 or B. breve DPC 6330 is relative to the ALA enriched diet, which was set to 1. Values are means ± SD, represented by vertical bars. Data was analysed by one-way ANOVA, followed by post hoc Tukey’s multiple comparison test. Significant differences are represented by *(P < 0.05).
Discussion
This study investigated the impact of dietary ALA enrichment with or without fatty acid conjugating B. breve supplementation on fat composition/distribution and the mechanisms through which commensal gut microbes alter lipid metabolism. The work presented herein tested the hypothesis that the alterations in host tissue fatty acid composition by B. breve could be linked to modulation of fat absorption processes in the small intestine and/or metabolism of fatty acids to longer-chain unsaturated derivatives in the liver.
Previously, we reported that dietary supplementation with linoleic acid and B. breve NCIMB 702258 significantly increased bioactive c9t11CLA concentrations in the liver of mice16, while dietary supplementation with B. breve NCIMB 702258 together with ALA increased tissue n-3 LC-PUFA in mice, compared with those that did not receive the strain17. More recently, we demonstrated an increase in brain DHA associated with B. breve NCIMB 702258 supplementation in mice18.
In general, ALA is a poor precursor for n-3 LC-PUFA, particularly DHA, first due to the efficiency for ALA to undergo beta-oxidation in the mitochondria40, rendering this fatty acid less available for desaturation and elongation and second because biosynthesis of DHA requires a crucial step in the peroxisome for partial beta-oxidation40. Therefore, any other event that increases desaturation and peroxisomal beta-oxidation would also favour DHA biosynthesis. Our recent data demonstrate that dietary CLA, in a specific range of ratio with ALA, significantly increased DHA biosynthesis (unpublished data), rendering some food, for example CLA enriched cheese, an unexpected source of DHA. Furthermore, associations between a reduced n-3 PUFA score following consumption of a CLA enriched cheese, decreased plasma AEA and an improved blood lipid profile in hypercholesterolaemic patients have been described44, thus exposing a link between n-3 PUFA, CLA, the ECS and adiposity. Interestingly, dietary CLA in a model of fatty liver was associated with increased hepatic PEA and OEA45. Based on these data, we were prompted to evaluate whether B. breve, possibly by increasing CLA formation, could modify the tissue fatty acid profile (as before), but this time identify whether EC and their congener biosynthesis were also affected. While CLA was increased in adipose tissue following supplementation of the ALA enriched diet with both B. breve strains, no change to the CLA concentration was observed in the liver, most likely due to increased hepatic peroxisomal beta-oxidation to CD16:2. This was demonstrated by the increased ratio of CD16:2: c9t11CLA, however we did not detect a subsequent increase in hepatic DHA concentration. Furthermore, we demonstrated that dietary ALA supplementation increased liver and adipose tissue ALA and EPA with decreased arachidonic acid levels in the adipose tissue. Highly polyunsaturated n-3 fatty acids replace arachidonic acid as an eicosanoid substrate in cell membranes46, resulting in decreased production of arachidonic acid-derived proinflammatory eicosanoids such as prostaglandin E2 and leukotriene B447 thus, increased tissue EPA could be beneficial in a variety of chronic inflammatory settings such as non-alcoholic fatty liver disease (NAFLD) and IBD. The overall hepatic n-3 PUFA score was unsurprisingly increased in all ALA enriched dietary groups, compared with the control. B. breve NCIMB 702258 in combination with the ALA enriched diet increased EPA and DHA in adipose tissue, while B. breve DPC 6330 increased DHA only in the adipose tissue, compared with the ALA enriched control group.
Analysis of the EC revealed a decrease in hepatic AEA, OEA and 2-AG in all ALA enriched groups, compared with the control. Furthermore, both B. breve strains further reduced AEA and PEA in the liver, compared with the ALA enriched control group. In contrast, further supplementation of the ALA enriched diet with B. breve NCIMB 702258 specifically increased AEA, PEA, OEA and DHEA in the adipose tissue. These data suggest that the effect of B. breve NCIMB 702258 on the ECS may be similar to those triggered by inflammation. From our experimental conditions, intake of B. breve NCIMB 702258 triggered a cascade linked to activation of toll-like receptors (TLRs)48, but we cannot rule out the possibility that some of the effects might be mediated by CLA, such as the increase in adipose tissue PEA and OEA, as previously demonstrated in the liver of rats fed CLA45. Interestingly, feeding of both strains increased CLA in adipose tissue. However, only B. breve NCIMB 702258 enhanced AEA, PEA, OEA and DHEA adipose tissue levels, which suggest that even among strains of the same species, different metabolic responses occur on the ECS. The increased adipose tissue AEA could be related to TLR activation as previously demonstrated in lymphocytes, where fatty acid amide hydrolase activity was downregulated49. Therefore, these data suggest that concurrent effects of these nutritional factors may result in changes to the ECS.
We have demonstrated an ALA enriched dietary effect on the hepatic mRNA expression of Δ-6-desaturase. This enzyme is involved in the biosynthetic pathway of long-chain, highly unsaturated PUFA50 and so may partly explain the increased EPA concentrations in the liver and adipose tissue. Beyond docosapentaenoic acid formation, Δ-6-desaturase also metabolises tetracosapentaenoic acid (C24:5n-3), a precursor to DHA, however since Δ-6-desaturase is used by two substrates in the same pathway of n-3 PUFA metabolism, there is intrinsic potential for competitive substrate inhibition to occur51. Previous studies revealed that high dietary ALA content up-regulates hepatic Δ-6-desaturase gene expression levels in different models42,52. The hepatic mRNA expression of ELOVL-5 was not found to be increased for all ALA enriched dietary groups. One recent study demonstrating the effect of ALA supplementation on Δ-6-desaturase and ELOVL-5 expression in the common carp showed that after 42 days, while Δ-6-desaturase activity decreased, there was an increase in ELOVL-5 expression42. This suggests that while the activity of one rate-limiting enzyme involved in the metabolism of n-3 PUFA increases, there may not be a corresponding increase in the activity of the other at the same time, thus the sequence of enzyme activity is time dependent.
Supplementation of B. breve DPC 6330 in an ALA enriched diet had a significant effect on increasing the mRNA expression of genes involved in the transport and uptake of fatty acids to the distal small intestine. The mRNA expression of FABP-1 and CD-36 were increased in the ileum following supplementation of the ALA enriched diet with B. breve DPC 6330. FABP-1 and CD-36 glycoprotein, which play a role in cellular adhesion and in the regulation of fatty acid transport and uptake, have previously been shown to be increased in the intestine following in vivo exposure of human intestinal mucosa to the probiotic species Lactobacillus plantarum WCFSI53. Furthermore, the role of the microbiota in stimulating dietary fatty acid absorption in the intestinal epithelium have previously been demonstrated54.
Hepatic TAG levels were reduced following 6 weeks of dietary intervention with the ALA enriched diet. It has been reported that EPA may be a promising novel therapy to decrease hepatic TAG55,56 and may therefore be of importance for the treatment of NAFLD. Coincidently, all ALA dietary groups were associated with increased hepatic EPA. In addition, all ALA enriched dietary groups had reduced hepatic FAS mRNA expression and lower SCD-1 expression whereby the decrease in SREBP-1c regulated enzymes had a significant effect on fatty acid synthesis in the liver57. Supplementation of the ALA enriched diet with both B. breve strains decreased hepatic TAG levels compared with the ALA only group. These data demonstrate a potential for administered B. breve to reduce liver TAG levels in the host. Hepatic TAG levels, the mRNA expression of CD-36 and FABP-1 were all decreased following further supplementation of the ALA enriched diet with B. breve, compared to the ALA enriched group, whereby CD-36 and FABP-1 are involved in transport and uptake of fatty acids to the liver58,59. This indicates a strain-specific effect of these two B. breve strains toward reducing hepatic TAG levels while also reducing the hepatic mRNA expression of genes relating to reduced fatty acid uptake to the liver. A potential therapeutic role for commensal bacteria in the treatment of NAFLD have been described60. For example, it was reported that dietary supplementation with Lactobacillus plantarum MA2 with a cholesterol enriched diet resulted in reduced liver TAG and increased numbers of fecal Lactobacillus and Bifidobacterium in rats61. Furthermore, the potential of VSL#3, a multistrain preparation composed of Streptococcus thermophilus and several strains of Lactobacillus and Bifidobacterium for reducing inflammation (reduced TNFα and cyclooxygenase expression, increased PPARα expression) and oxidative damage (inducible nitric oxide synthase) to the liver while ameliorating the liver lipid profiles has been shown62.
The data presented in this study confirmed that dietary ALA positively impacted on tissue fatty acid composition, increased hepatic n-3 PUFA metabolism and reduced hepatic TAG levels, hepatic fatty acid synthesis enzymes and hepatic uptake of fatty acids and demonstrated effects on the ECS. Previously, we demonstrated that B. breve NCIMB 702258 in an ALA enriched diet increased hepatic EPA concentrations, compared with ALA alone17. We have elaborated on these previous findings by demonstrating an impact not only of B. breve NCIMB 702258 in an ALA enriched diet, but also of B. breve DPC 6330 toward reducing liver triglyceride levels, altering fatty acid and EC profiles and expression of key enzymes involved in the metabolism of fatty acids. Supplementation with either B. breve strain in combination with an ALA enriched diet reduced liver TAG levels which was associated with reduced hepatic CD-36 and FABP-1 mRNA expression, involved in the uptake of fatty acids to the liver. While it is clear from the data that ALA impacted on desaturase activity, there was little effect of B. breve supplementation on n-3 PUFA metabolism gene expression in the liver. Instead, the increased peroxisomal beta-oxidation caused by the strains, could explain why there was no increase in CLA identified in the liver. Adipose tissue CLA was increased, as a result of B. breve supplementation. Future studies will be devoted to evaluating the possible synergistic effects of CLA enriched products with B. breve: (i) in maintaining/re-equilibrating energy and lipid metabolism, by modulating tissue EC and congener profiles, (ii) acting on energy metabolism and EPA and DHA related molecules, and (iii) to assess if there is an optimal level of dietary EPA and DHA to allow such a synergistic activity. This novel study identifies mechanisms through which these lipid-modulating Bifidobacterium strains may be acting in vivo. Considering that the relative proportions of Bifidobacterium in the gut microbiota of C57BL/6 J have previously been reported to compose 2–3% only of the total gut microbial content18 (mainly Bifidobacterium animalis and Bifidobacterium pseudolongum63), it is indeed intriguing to identify probiotic manipulations of the gut microbiota which can have a positive impact on health. This nutritional approach may prove helpful in the alleviation of metabolic disorders such as those causing NAFLD, where CLA45, n-3 PUFA64,65 and manipulation of the gut microbiota66 have been shown to decrease adiposity accumulation in the liver.
Methods
Preparation and administration of B. breve NCIMB 702258 and B. breve DPC 6330
Rifampicin resistant variants of B. breve NCIMB 702258 and B. breve DPC 6330 were isolated by spread-plating ~109 colony forming units (CFU) from an overnight culture onto MRS agar (de Man, Rogosa & Sharpe; Difco Laboratories, Detroit, MI, USA), supplemented with 0.05% (w/v) L-cysteine hydrochloride (Sigma-Aldrich, Wicklow, Ireland) (mMRS), containing 500 μg/mL rifampicin (Sigma-Aldrich). Following anaerobic incubation (anaerobic jars with Anaerocult A gas packs; Merck, Darmstadt, Germany) at 37 °C for 3 days, colonies were stocked in mMRS broth containing 40% (v/v) glycerol and stored at −80 °C. To confirm that the rifampicin resistant variants were identical to the parent strain, molecular fingerprinting using pulse-field gel electrophoresis was employed.
Prior to freeze drying, B. breve NCIMB 702258 and B. breve DPC 6330 were grown in mMRS by incubating for 48 hr at 37 °C under anaerobic conditions. The culture was washed twice in phosphate buffered saline (PBS) and resuspended at a concentration of ~2 × 1010 cells/mL in 15% (w/v) trehalose (Sigma-Aldrich) in dH2O. One millilitre aliquots were freeze-dried using a 24 hr programme (freeze temp. −40 °C, condenser set point −60, vacuum set point 600 m Torr). Animals were each administered approximately 1 × 109 live microorganisms per day, which was achieved by resuspending appropriate quantities of freeze-dried powder in water which was consumed ad libitum. Mice that did not receive the bacterial strains received placebo freeze-dried powder (15% (w/v) trehalose in dH2O). Water containing either the bacterial strains or placebo freeze-dried powder was the only water supplied to the mice throughout the trial. Freeze dried powders with the bacterial strains underwent continuous quality control checks of cell counts for the duration of the trial, by plating serial dilutions on mMRS agar supplemented with 100 μg/mL of mupirocin (Oxoid) and 100 μg/mL rifampicin (Sigma-Aldrich) and incubating plates anaerobically for 72 hr at 37 °C.
Animals and Treatment
All animal experiments were approved by University College Cork (UCC) Animal Ethics Committee and experimental procedures were conducted under the appropriate license from the Irish Government. Female BALB/c mice were purchased from Harlan Ltd. (Briester, Oxon, UK) at 8 weeks of age and housed under barrier-maintained conditions within the Biological Services Unit, UCC. Mice were allowed to acclimatise for 1 week prior to commencement of the study and were fed ad libitum with Teklad Global Rodent Standard Diet (Harlan Laboratories, Madison, WI, USA, #2018 S), with free access to water at all times. Mice were housed in groups of five per cage and maintained in a controlled environment at 25 °C under 12-hr-light/12-hr-dark cycle. After 1 week of acclimatisation, animals were randomly divided into four groups (n 10 mice per group): (1) unsupplemented control group (CON) fed a standard rodent diet with placebo freeze-dried powder (15% w/v trehalose in drinking water); (2) flaxseed oil (FXO, enriched in ALA) supplemented group (ALA-CON) with placebo freeze-dried powder (15% w/v trehalose in drinking water); (3) FXO supplemented group with B. breve NCIMB 702258 (ALA + NCIMB 702258) (approximate daily dose of 109 microorganisms); (4) FXO supplemented group with B. breve DPC 6330 (ALA + DPC 6330) (approximate daily dose of 109 microorganisms).
The unsupplemented control diet contained the following composition: corn starch (32.45%), casein (20.0%), sucrose (15.0%), maltodextrin (12.0%), cellulose (5.0%), mineral mix (3.5%), vitamin mix (1.5%), L-cysteine (0.3%), choline bitartrate (0.25%), TBHQ antioxidant (0.002%) and the following composition of fat: palm oil (3.0%), safflower oil (3.0%), olive oil (3.0%), FXO (1.0%). The n-6:n-3 ratio of this diet was ~5.3 and the diet contained ~0.5% ALA. The FXO supplemented diet contained the following nutrient composition: corn starch (32.45%), casein (20.0%), sucrose (15.0%), maltodextrin (12.0%), cellulose (5.0%), mineral mix (3.5%), vitamin mix (1.5%), L-cysteine (0.3%), choline bitartrate (0.25%), TBHQ antioxidant (0.002%) and the following composition of fat: FXO (5.5%), palm oil (1.5%), safflower oil (1.5%), olive oil (1.5%). The n-6:n-3 ratio of this diet was ~0.75 and the diet contained ~3% ALA.
Body weight and food intake were assessed weekly. Following 6 weeks on experimental diets, the animals were sacrificed by cervical dislocation. Liver, brain, fat pads (epididymal, perirenal and mesenteric), gastrointestinal tract from stomach to anus, and cecal contents were removed, blotted dry on filter paper, weighed and flash-frozen immediately in liquid nitrogen. All samples were stored at −80 °C until processed. Blood samples were collected into serum collection tubes (BD Diagnostics, Oxford, UK) from fasted animals, and allowed to clot for at least 30 min at 4 °C before centrifugation for 20 min at 10,000 g to separate the serum.
Culture Dependent Microbial Analysis
Fresh fecal samples were taken from BALB/c mice every second week for microbial analysis. Microbial analysis of the fecal samples involved enumeration of the B. breve strains by plating serial dilutions on MRS agar supplemented with 100 μg/mL of mupirocin (Oxoid), 100 μg/mL of rifampicin (Sigma-Aldrich) and 50 units/mL of nystatin (Sigma-Aldrich). Agar plates were incubated anaerobically for 72 h at 37 °C.
Lipid Extraction and Fatty Acid Analysis
An aliquot of the lipid fraction for each sample was mildly saponified using a procedure in order to obtain free fatty acids (FFA) for high performance liquid chromatography (HPLC) analysis. Lipid extracts were dissolved in 5 ml of ethanol, 100 μl of desferal (25 mg/ml ddH2O), 1 ml of a 25% (v/v) solution of ascorbic acid in water, 0.5 ml of 10 N KOH, and left for 14 h in the dark at room temperature. Following this, 10 ml of n-hexane and 7 ml of ddH2O were added. The samples were then acidified with 0.35 ml of 37% (v/v) HCI, to pH 3–4. Samples were centrifuged for 1 h at 900 g. The hexane phase containing FFAs was collected, the solvent evaporated, and the residue was dissolved in 0.5 ml of CH3CN/0.14% (v/v) CH3COOH.
Separation of FAs, including CLA and its metabolites, was performed on an Agilent 1100 HPLC system (Agilent, Palo Alto, Calif., USA) equipped with a diode array detector. A C-18 Inertsil 5 ODS-2 Chrompack column (Chrompack International BV, Middleburg, The Netherlands), 5 μm particle size, 150 × 4.6 mm, was used with a mobile phase of CH3CN/H2O/CH3COOH (70/30/0.12, v/v/v) at a flow rate of 1.5 ml/min67. Conjugated diene unsaturated FAs were detected at 234 nm. Spectra (195–315 nm) of the eluate were obtained every 1.28 s and were electronically stored. Second-derivate UV spectra of the conjugated diene FAs were generated using Phoenix 3D HP Chemstation software (Agilent, Palo Alto, CA). These spectra were acquired to confirm identification of the HPLC peaks68.
Since SFAs are transparent to UV, after derivatization, they were measured as fatty acid methyl esters (FAME) using gas chromatography (Agilent, Model 6890, Palo Alto, CA) equipped with split ratio of 20:1 injection port, a flame ionization detector (FID), an autosampler (Agilent, Model 7673), a 100 m HP-88 fused capillary column (Agilent). Finally, data were analysed by the Agilent ChemStation software system. The injector and detector temperatures were set up at 250 °C and 280 °C, respectively. H2 served as carrier gas (1 ml/min), and the FID gases were H2 (30 ml/min), N2 (30 ml/min), and purified air (300 ml/min). The temperature program was as follows: initial temperature was 120 °C, programmed at 10 °C/min to 210 °C and 5 °C/min to 230 °C, then programmed at 25 °C/min to 250 °C and held for 2 min64.
Measurement of TAG in the Liver and Serum
The lipids from 50 mg of frozen liver were extracted and purified according to the method of Folch69. Liver lipids were extracted using chloroform-methanol (2:1, v:v; Thermo Scientific) and an aliquot of the organic phase was collected, dried and resuspended in duplicate in Infinity-TAG lipid-stable reagent (Thermo Scientific). TAG levels were determined according to the manufacturer’s instructions and lipids were quantified using a TAG chemistry calibrator (Pointe Scientific Inc., MI, USA). Serum TAG levels were analysed in duplicate using the commercial L-Type TAG M kit (Wako Diagnostics, Neuss, Germany).
Analysis of Endocannabinoids and their Congeners
Aliquots of the organic phase containing extracted lipids were evaporated to dryness under vacuum, and reconstituted with 0.4 ml of 100% (v/v) methanol. Quantification of AEA, 2-AG, PEA and OEA, was performed by liquid chromatography-atmospheric pressure chemical ionization-mass spectrometry (LC-APCI-MS), and using selected ion monitoring (SIM) at M + 1 values for the four compounds and their deuterated homologues70. A C-18 Zorbax Eclipse Plus column (Agilent, Palo Alto, CA) 5 μm particle size, 50 × 4.6 mm was used with a mobile phase of CH3OH/H2O/CH3COOH (80/20/0.3, v/v/v) at a flow rate of 0.5 ml/min.
RNA Extractions and Complementary DNA Synthesis
Total RNA was isolated from liver and ileum using the commercial RNeasy Mini-Kit (Qiagen, West Sussex, UK), according to the manufacturer’s instructions. Total isolated RNA was quantified using the Nanodrop (Thermo Scientific). Single stranded complementary DNA (cDNA) was synthesised from 1 μg of total RNA using 2.5 ng/μL random primers (Promega, WI, USA), 10 mM PCR nucleotide mix (Promega), 40 units/μL RNasin Plus RNase inhibitor (Promega) and the Im-Prom II reverse transcriptase (Promega).
Real-Time PCR Analysis
Amplification of generated cDNA was performed in the Lightcycler 480 system (Roche Diagnostics Ltd., West Sussex, UK) using 0.25 μM primers (MWG Eurofins, Ebersberg, Germany), 1 μL cDNA and the Lightcycler 480 SYBR Green I Master kit (Roche Diagnostics Ltd). Real-time PCR conditions were set at: 95 °C for 10 min followed by 50 cycles at 95 °C for 10 sec, 60 °C for 5 sec and 72 °C for 15 sec. Specific forward and reverse primers used to amplify cDNA were newly designed and are listed in Table 4. All samples were analysed in duplicate and normalised to β-actin, as a constitutively expressed control gene. Melting curve analysis allowed the validation of the authenticity of the real-time PCR products. Basic relative quantification of expression was determined using the comparative 2−ΔΔCt method.
Statistical Analysis
All results are presented as means ± SD (per group). To assess whether differences between the treatment groups were significant, data were analysed using one-way ANOVA followed by post hoc Tukey’s multiple comparison test using GraphPad Prism version 4.0 for Windows (GraphPad Software). Statistical significance was accepted at P < 0.05. Grubbs method was used to test for outliers71.
Additional Information
How to cite this article: Patterson, E. et al. Bifidobacterium breve with α-linolenic acid alters the composition, distribution and transcription factor activity associated with metabolism and absorption of fat. Sci. Rep. 7, 43300; doi: 10.1038/srep43300 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Backhed, F. et al. The gut microbiota as an environmental factor that regulates fat storage. Proc. Nal. Acad. Sci. USA 101, 15718–15723 (2004).
Turnbaugh, P. J. et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 444, 1027–1031 (2006).
Backhed, F., Manchester, J. K., Semenkovich, C. F. & Gordon, J. I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 104, 979–984 (2007).
Everard, A. et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 60, 2775–2786 (2011).
Cani, P. D. et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 56, 1761–1772 (2007).
Cani, P. D. et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 57, 1470–1481 (2008).
Cani, P. D. & Delzenne, N. M. The role of the gut microbiota in energy metabolism and metabolic disease. Current Pharmaceutical Design. 15, 1546–1558 (2009).
Cani, P. D. et al. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 50, 2374–2383 (2007).
Di Marzo, V. et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature. 410, 822–825 (2001).
Duncan, M., Davison, J. S. & Sharkey, K. A. Review article: Endocannabinoids and their receptors in the enteric nervous system. Aliment. Pharmacol. Ther. 22, 667–683 (2005).
Muccioli, G. G. et al. The endocannabinoid system links gut microbiota to adipogenesis. Molecular systems biology. 6, 392 (2010).
Geurts, L. et al. Altered gut microbiota and endocannabinoid system tone in obese and diabetic leptin-resistant mice: Impact on apelin regulation in adipose tissue. Frontiers in microbiology. 2, 149 (2011).
Barrett, E., Ross, R. P., Fitzgerald, G. F. & Stanton, C. Rapid screening method for analyzing the conjugated linoleic acid production capabilities of bacterial cultures. Appl. Environ. Microbiol. 73, 2333–2337 (2007).
Coakley, M. et al. Conjugated linoleic acid biosynthesis by human-derived bifidobacterium species. J. Appl. Microbiol. 94, 138–145 (2003).
Hennessy, A. A. et al. The production of conjugated alpha-linolenic, gamma-linolenic and stearidonic acids by strains of bifidobacteria and propionibacteria. Lipids. 47, 313–327 (2012).
Wall, R. et al. Metabolic activity of the enteric microbiota influences the fatty acid composition of murine and porcine liver and adipose tissues. Am. J. Clin. Nutr. 89, 1393–1401 (2009).
Wall, R. et al. Impact of administered bifidobacterium on murine host fatty acid composition. Lipids. 45, 429–436 (2010).
Wall, R. et al. Contrasting effects of bifidobacterium breve ncimb 702258 and bifidobacterium breve dpc 6330 on the composition of murine brain fatty acids and gut microbiota. Am. J. Clin. Nutr. 95, 1278–1287 (2012).
Barrett, E. et al. Bifidobacterium breve with alpha-linolenic acid and linoleic acid alters fatty acid metabolism in the maternal separation model of irritable bowel syndrome. Plos One. 7, e48159 (2012).
Kritchevsky, D. et al. Conjugated linoleic acid isomer effects in atherosclerosis: Growth and regression of lesions. Lipids. 39, 611–616 (2004).
Henriksen, E. J. et al. Isomer-specific actions of conjugated linoleic acid on muscle glucose transport in the obese zucker rat. Am. J. Physiol. Endocrinol. Metab. 285, E98–E105 (2003).
Ryder, J. W. et al. Isomer-specific antidiabetic properties of conjugated linoleic acid. Improved glucose tolerance, skeletal muscle insulin action, and ucp-2 gene expression. Diabetes. 50, 1149–1157 (2001).
Coakley, M. et al. Inhibitory effect of conjugated alpha-linolenic acid from bifidobacteria of intestinal origin on sw480 cancer cells. Lipids. 44, 249–256 (2009).
Kim, N. D. et al. Chemopreventive and adjuvant therapeutic potential of pomegranate (punica granatum) for human breast cancer. Breast Cancer Res Treat. 71, 203–217 (2002).
Kohno, H. et al. Pomegranate seed oil rich in conjugated linolenic acid suppresses chemically induced colon carcinogenesis in rats. Cancer Sci. 95, 481–486 (2004).
Devery, R., Miller, A. & Stanton, C. Conjugated linoleic acid and oxidative behaviour in cancer cells. Biochem. Soc. Trans. 29, 341–344 (2001).
Beppu, F. et al. Potent inhibitory effect of trans9, trans11 isomer of conjugated linoleic acid on the growth of human colon cancer cells. J. Nutr. Biochem. 17, 830–836 (2006).
Koba, K. et al. Dietary conjugated linolenic acid in relation to cla differently modifies body fat mass and serum and liver lipid levels in rats. Lipids. 37, 343–350 (2002).
Arao, K. et al. Dietary effect of pomegranate seed oil rich in 9cis, 11trans, 13cis conjugated linolenic acid on lipid metabolism in obese, hyperlipidemic oletf rats. Lipids Health Dis. 3, 24 (2004).
Koba, K. et al. Genetically modified rapeseed oil containing cis-9,trans-11,cis-13-octadecatrienoic acid affects body fat mass and lipid metabolism in mice. J. Agric. Food Chem. 55, 3741–3748 (2007).
Chardigny, J. M. et al. Effect of conjugated fa on feed intake, body composition, and liver fa in mice. Lipids. 38, 895–902 (2003).
Zambell, K. L. et al. Conjugated linoleic acid supplementation in humans: Effects on body composition and energy expenditure. Lipids. 35, 777–782 (2000).
Javadi, M. et al. The effect of six different c18 fatty acids on body fat and energy metabolism in mice. Br. J. Nutr. 92, 391–399 (2004).
Calder, P. C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Bba-Mol. Cell Biol. 1851, 469–484 (2015).
Marques, T. M. et al. Gut microbiota modulation and implications for host health: Dietary strategies to influence the gut-brain axis. Innov. Food Sci. & Emerg. Technol. 22, 239–247 (2013).
Patterson, E. et al. Gut microbiota, the pharmabiotics they produce and host health. Proc. Nutr. Soc. 73, 477–489 (2014).
Robertson, R. C. et al. Omega-3 polyunsaturated fatty acids critically regulate behaviour and gut microbiota development in adolescence and adulthood. Brain Behav. Immun. 59, 21–37 (2016).
Patterson, E., Wall, R., Fitzgerald, G., Ross, R. & Stanton, C. Health implications of high dietary omega-6 polyunsaturated fatty acids. J. Nutr. Metab. (2012), Article ID 539426, 16 pages (2012).
Pan, A. et al. Α-linolenic acid and risk of cardiovascular disease: A systematic review and meta-analysis. Am. J. Clin. Nutr. 96, 1262–73 (2012).
Burdge, G. C. Metabolism of alpha-linolenic acid in humans. Prostaglandins Leukot. Essent. Fatty Acids. 75, 161–168 (2006).
Ander, B. P. et al. Distribution of omega-3 fatty acids in tissues of rabbits fed a flaxseed-supplemented diet. Metabolism. 59, 620–627 (2010).
Ren, H. T., Yu, J. H., Xu, P. & Tang, Y. K. Influence of dietary fatty acids on muscle fatty acid composition and expression levels of delta6 desaturase-like and elovl5-like elongase in common carp (cyprinus carpio var. Jian). Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 163, 184–192 (2012).
Singh, K., Mridula, D., Rehal, J. & Barnwal, P. Flaxseed: A potential source of food, feed and fiber. Crit. Rev. Food Sci. Nutr. 51, 210–222 (2011).
Pintus, S. et al. Sheep cheese naturally enriched in alpha-linolenic, conjugated linoleic and vaccenic acids improves the lipid profile and reduces anandamide in the plasma of hypercholesterolaemic subjects. Br. J. Nutr. 109, 1453–1462 (2013).
Piras, A. et al. Effects of dietary cla on n-3 hufa score and n-acylethanolamides biosynthesis in the liver of obese zucker rats. Prostaglandins Leukot. Essent. Fatty Acids. 98, 15–19 (2015).
Simopoulos, A. P. Importance of the ratio of omega-6/omega-3 essential fatty acids: Evolutionary aspects. World Rev. Nutr. Diet. 92, 1–22 (2003).
Calder, P. C. Polyunsaturated fatty acids and inflammation. Prostaglandins Leukot. Essent. Fatty Acids. 75, 197–202 (2006).
Plantinga, T. S. et al. Differential toll-like receptor recognition and induction of cytokine profile by bifidobacterium breve and lactobacillus strains of probiotics. Clin.Vaccine Immunol. 18 (2011).
Maccarrone, M. et al. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch. Biochem. Biophys. 393, 321–328 (2001).
Stoffel, W. et al. Δ6-desaturase (fads2) deficiency unveils the role of ω3-and ω6-polyunsaturated fatty acids. The EMBO Journal. 27, 2281–2292 (2008).
Gregory, M. K., Gibson, R. A., Cook-Johnson, R. J., Cleland, L. G. & James, M. J. Elongase reactions as control points in long-chain polyunsaturated fatty acid synthesis. Plos One. 6 (2011).
Li, Y. Y. et al. The effects of dietary fatty acids on liver fatty acid composition and delta(6)-desaturase expression differ with ambient salinities in siganus canaliculatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 151, 183–190 (2008).
Troost, F. J. et al. Identification of the transcriptional response of human intestinal mucosa to lactobacillus plantarum wcfs1 in vivo . BMC Genomics. 9, 374 (2008).
Semova, I. et al. Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe 12, 277–288 (2012).
Masterton, G. S., Plevris, J. N. & Hayes, P. C. Review article: Omega-3 fatty acids - a promising novel therapy for non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 31, 679–692 (2010).
Shapiro, H. et al. The therapeutic potential of long-chain omega-3 fatty acids in nonalcoholic fatty liver disease. Clinical Nutrition. 30, 6–19 (2011).
Tandy, S. et al. Dietary krill oil supplementation reduces hepatic steatosis, glycemia, and hypercholesterolemia in high-fat-fed mice. J. Agric. Food Chem. 57, 9339–9345 (2009).
Newberry, E. P. et al. Decreased hepatic triglyceride accumulation and altered fatty acid uptake in mice with deletion of the liver fatty acid-binding protein gene. J. Biol. Chem. 278, 51664–51672 (2003).
Garcia-Monzon, C. et al. Hepatic insulin resistance is associated with increased apoptosis and fibrogenesis in nonalcoholic steatohepatitis and chronic hepatitis c. J. Hepatol. 54, 142–152 (2011).
Iacono, A., Raso, G. M., Canani, R. B., Calignano, A. & Meli, R. Probiotics as an emerging therapeutic strategy to treat nafld: Focus on molecular and biochemical mechanisms. J. Nutr. Biochem. 22, 699–711 (2011).
Wang, Y. et al. Effects of lactobacillus plantarum ma2 isolated from tibet kefir on lipid metabolism and intestinal microflora of rats fed on high-cholesterol diet. Appl. Microbiol. Biotechnol. 84, 341–347 (2009).
Esposito, E. et al. Probiotics reduce the inflammatory response induced by a high-fat diet in the liver of young rats. J. Nutr. 139, 905–911 (2009).
Shin, J. et al. Analysis of the mouse gut microbiome using full-length 16s rrna amplicon sequencing. Sci. Rep. 6 (2016).
Batetta, B. et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese zucker rats. J. Nutr. 139, 1495–1501 (2009).
Berge, K. et al. Chronic treatment with krill powder reduces plasma triglyceride and anandamide levels in mildly obese men. Lipids Health Dis. 12, 78 (2013).
Le Roy, T. et al. Intestinal microbiota determines development of non-alcoholic fatty liver disease in mice. Gut. 62, 1787–1794 (2013).
Melis, M. P. et al. Characterization of conjugated linoleic acid and its metabolites by rp-hplc with diode array detector. Eur. J. Lipid Sci. Tech. 103, 617–621 (2001).
Angioni, E. et al. Uv spectral properties of lipids as a tool for their identification. Eur. J. Lipid Sci. Tech. 104, 59–64 (2002).
Folch, L. M. & Sloane-Stanley G. H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497–509 (1957).
Di Marzo, V. et al. Changes in plasma endocannabinoid levels in viscerally obese men following a 1 year lifestyle modification programme and waist circumference reduction: Associations with changes in metabolic risk factors. Diabetologia. 52, 213–217 (2009).
Grubbs, F. E. Sample criteria for testing outlying observations. Ann. Math. Stat. 27–58 (1950).
Acknowledgements
The authors wish to acknowledge the technical assistance provided by staff of the Biological Services Unit, UCC and Dr. David Russell. The APC Microbiome Institute is a research centre funded by Science Foundation Ireland (SFI). This publication has emanated from research supported by a research grant from Science Foundation Ireland (SFI) under grant number SFI/12/RC/2273.
Author information
Authors and Affiliations
Contributions
The authors’ responsibilities were as follows - E.P., R.W., J.F.C., T.G.D., R.P.R., E.Q.Q., F.S., G.F.F. and C.S. designed the research, E.P., R.W. and S.L. conducted the research; E.P., R.W., S.L., R.P.R., S.B. and C.S. analysed the data; E.P., R.W., S.L. and C.S. wrote the manuscript; and C.S. had primary responsibility for the final content.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Patterson, E., Wall, R., Lisai, S. et al. Bifidobacterium breve with α-linolenic acid alters the composition, distribution and transcription factor activity associated with metabolism and absorption of fat. Sci Rep 7, 43300 (2017). https://doi.org/10.1038/srep43300
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep43300
This article is cited by
-
The gut microbiota in persistent post-operative pain following breast cancer surgery
Scientific Reports (2024)
-
Linoleic acid induces metabolic stress in the intestinal microorganism Bifidobacterium breve DSM 20213
Scientific Reports (2020)
-
Effect of probiotics on obesity-related markers per enterotype: a double-blind, placebo-controlled, randomized clinical trial
EPMA Journal (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
