
Abstract
The mechanism of selectivity in ion channels is still an open question in biology for more than half a century. Here, we suggest that quantum interference can be a solution to explain the selectivity mechanism in ion channels since interference happens between similar ions through the same size of ion channels. In this paper, we simulate two neighboring ion channels on a cell membrane with the famous double-slit experiment in physics to investigate whether there is any possibility of matter-wave interference of ions via movement through ion channels. Our obtained decoherence timescales indicate that the quantum states of ions can only survive for short times, i.e. ≈100 picoseconds in each channel and ≈17–53 picoseconds outside the channels, giving the result that the quantum interference of ions seems unlikely due to environmental decoherence. However, we discuss our results and raise few points, which increase the possibility of interference.
Similar content being viewed by others
Introduction
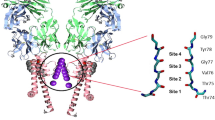
Recently, the quantum world has opened up new perspectives in the field of complex systems and biology1,2,3,4,5,6. Energy, charge, or information transfer are important phenomena in physical and biological systems taking place at scales ranging from atoms to large macro-molecular structures, and the idea has been put forward that quantum mechanics might have a positive effect on the efficiency of energy or charge transport in such systems. Energy transfer in photosynthetic structures7,8,9,10, avian quantum compass in migratory birds11, and charge transport through DNA12 are good examples in this context. The living cell is an information replicating and processing system that is replete with naturally-evolved nanomachines, which at some level may require a quantum mechanical description despite critical limitations for application of quantum theory in biology13. In this case, ion channels are good examples in living cells in which quantum effects may play a role14,15,16,17,18,19,20. They are proteins in the membrane of cells that can cooperate for the onset and propagation of electrical signals across membranes by providing a highly selective conduction of charges bound to ions through a channel like structure. In fact, each ion channel is specialized for specific ions, e.g. potassium channels only permit potassium ions to pass the membrane while they reject other ions (e.g. sodium) to pass. This property is called selectivity and the important part of the ion channel, which is responsible for selectivity, is called selectivity filter. Numerous investigations of ion selectivity have been conducted over more than 50 years, yet the mechanisms whereby the channels select certain ions and reject others are not well understood21. The selectivity filter is a part of the protein forming a narrow tunnel inside the ion channel which is responsible for the selection process and fast conduction of ions across the membrane22. The 3.4 nanometer long KcsA channel (i.e. potassium crystallographically-sited activation channel) is comprised of a 1.2 nanometer long selectivity filter that is composed of four P-loop strands. The Carbonyl groups (i.e. C=O’s) are responsible for trapping and displacement of ions in the filter (see Fig. 1).

(Left) A representation of KcsA ion channel. (Right) Two P-loop monomers in the selectivity filter, composed of the sequences of TVGYG amino acids [T(Threonine, Thr75), V(Valine, Val76), G(Glycine, Gly77), Y(Tyrosine, Tyr78), G(Glycine, Gly79)] linked by peptide units H-N-C =O20.
Here, we would like to investigate the possibility of matter-wave interference of ions via passing the ion channels. This possibility can be important as it may explain the “selectivity” property in ion channels because interference occurs between similar particles through similar slits with specific distances between them.
Our paper is organized as follows: we first introduce our simulation for two neighboring ion channels as a double-slit and then briefly investigate the possibility of matter-wave interference through the slits. Then, we estimate the plausible distances between the ion channels to produce ionic interference. In the next step, we consider the effect of environmental decoherence on quantum states of ions inside and outside the slits, and accordingly we obtain the coherence lengths of ions for making interference. Finally, we summarize and discuss our results.
A Simulation for two neighboring ion channels as a double-slit
Feynman believed that we can see the whole mysteries of quantum theory in the double-slit experiment23. Basically, quantum physics is centered on microscopic phenomena with photons, electrons and atoms, however objects of increasing complexity have attracted a growing scientific interest recently. For example, the matter-wave interference has been investigated theoretically and experimentally confirmed for large molecules such as C60 (i.e. 60 atoms), Tetraphenylporphyrin (i.e. 78 atoms) and functionalized Tetraphenylporphyrin (i.e. 810 atoms) in double-slit and gratings interferometers24,25,26,27,28,29. Now the question is: are these effects applicable in biology, too? If yes, can such matter-wave effects play any role in biological functions?
In this section, we would like to apply a simulation for quantum interference through neural ion channels. We expect to see quantum effects appearing due to small dimensions of the selectivity filter during the crossing of ions through ion channels. One of the quantum effects is the matter-wave interference of ions, which may show some quantum roots for action potential production in excitable cells (e.g. neurons).
Speed and wavelength of ions inside the selectivity filter
Here, we would like to obtain the velocity and consequently the de Broglie wavelength of ions (i.e. λdB = h/mv where h is the Planck’s cnstant, m is the mass of ion, and v is the velocity of ion) inside the selectivity filter via molecular dynamics (MD) simulation. In fact, our method (see Supplementary Information) is the same as what we used previously17,20 but we use it here with higher cutoffs, i.e. 1 nm for the van der Waals interaction and 1 nm for electrostatic interactions. Taking typical values for membrane potentials in neurons, −70 mV and +30 mV, for resting and firing states are considered30,31. To be more general, we also considered other membrane potentials −100 mV and +100 mV 32 to obtain other velocities as well17,20. The obtained results are shown in the Table 1, in which the order of the wavelength of ions (λ0 ≈ 0.1 nm) is the same as the order of the width of each slit (i.e. 0.3 nm), O(λ0) ≈ O(b), which is the necessary condition for interference.
The distance between ion channels
Now, we simulate two neighboring ion channels on a neural cell membrane with a double-slit experiment (see Fig. 2).

(Left) Ions passing through two neighboring ion channels on a cell membrane in a neuron, (Right) Simulation of two neighboring ion channels as a double-slit interferometer.
Here, we consider potassium ions passing the KcsA ion channels by focusing on the selectivity filter structure. As a matter of fact, ions act as wavepackets in the filter since they are trapped by carbonyl groups in the filter and consequently getting longer de Broglie wavelengths according to our MD simulation. We use the “macroscopicity” method33 to obtain the possible distances between the slits because in this method it is supposed that ion behaves like a Gaussian state, and after the slits the ion behaves like a free particle in two directions x and y. To clarify the macroscopicity method, we first introduce the dimensionless form of the Schrodinger equation. This method is described in more details in ref. 33 (see Supplementary Information) for obtaining interference pattern. In the dimensionless regime, we introduce the characteristic parameters for length R0 and energy U0 as constant units of length and energy of a quantum system, respectively. Also, for an ion mass, M, one can introduce the characteristic time as τ0 = R0/(U0/M)1/2. Since, U0 acts like the kinetic energy of quantum system, the unit of momentum could be expressed as  . Subsequently, the conjugate variables of position q and momentum p are defined as q = R/R0 and p = P/P0 where R and P are the conventional position and momentum, respectively34.
. Subsequently, the conjugate variables of position q and momentum p are defined as q = R/R0 and p = P/P0 where R and P are the conventional position and momentum, respectively34.
Moreover, the potential energy  and the Hamiltonian
and the Hamiltonian  operators of the system can be defined in this regime as
operators of the system can be defined in this regime as

where  and
and  are the potential energy and the Hamiltonian operators in the ordinary Schrodinger equation. Finally, the dimensionless Schrodinger equation can be written as
are the potential energy and the Hamiltonian operators in the ordinary Schrodinger equation. Finally, the dimensionless Schrodinger equation can be written as

Also, the canonical commutator in the dimensionless form is  , where
, where  defined as
defined as

So far, many studies have been done to investigate double-slit interference pattern of particles, atoms and molecules in experimental and theoretical contexts. In some of these works, the incoming state in double-slit experiment has been described by Gaussian wave packets35,36. Our approach in this study is based on Gaussian wave packet as a simulation for potassium ions which move through two neighboring ion channels in two dimensions. The use of Gaussian wave packet is sufficiently general, because it includes the limit case of plane waves. On the other hand, due to the development of experimental techniques, possible deviations from the standard form of the interference pattern can be better explained by Gaussian states37,38,39. As we mentioned above, the macroscopicity method is used for obtaining interference patterns by using a dimensionless form of the Schrodinger equation in which a new dimensionless parameter  appears showing quantitatively the quantum behavior of the system. We can define
appears showing quantitatively the quantum behavior of the system. We can define  as (see Supplementary Information)
as (see Supplementary Information)

where  . Here, λ0 is the de Broglie wavelength of the system. Strictly speaking, the situation in which one obtains
. Here, λ0 is the de Broglie wavelength of the system. Strictly speaking, the situation in which one obtains  , the system behaves quasi-classically. The values of
, the system behaves quasi-classically. The values of  between 0.01 to 0.1 are fair enough to show the macroscopic disposition of the proposed system to behave quantum mechanically34. If we let λ ≈ 0.1 nm as the average de Broglie wavelength of the ion inside the selectivity filter and then let R0 ≈ 0.2 nm as the size (or diameter) of the ion then
between 0.01 to 0.1 are fair enough to show the macroscopic disposition of the proposed system to behave quantum mechanically34. If we let λ ≈ 0.1 nm as the average de Broglie wavelength of the ion inside the selectivity filter and then let R0 ≈ 0.2 nm as the size (or diameter) of the ion then  which falls in the quantum region. Now, we obtain the real values of the distance between two neighboring ion channels, d′, according to different measures of
which falls in the quantum region. Now, we obtain the real values of the distance between two neighboring ion channels, d′, according to different measures of  . For this purpose, we draw the interference patterns according to the equation Supplementary Information - 13 in which b is the width of selectivity filter, and d is a variable parameter similar to the real range of distances between ion channels for potassium ion. Notice that we use dimensiomless form of b and d for drawing of the interference patterns. In the dimensionless regime, d can be defined as d = d′/R0 and b = b′/R0 where d′ is the distance between the slits, b′ is the width of selectivity filter (i.e. 0.3 nm) and R0 is the size of potassium ion (≅0.2 nm) as a characteristic parameters for length. For interference patterns in the regions of our investigation, Figures (SI-1–SI-4) and Fig. 3, the different values of the distance between ion channels in relation with macroscopicity measure
. For this purpose, we draw the interference patterns according to the equation Supplementary Information - 13 in which b is the width of selectivity filter, and d is a variable parameter similar to the real range of distances between ion channels for potassium ion. Notice that we use dimensiomless form of b and d for drawing of the interference patterns. In the dimensionless regime, d can be defined as d = d′/R0 and b = b′/R0 where d′ is the distance between the slits, b′ is the width of selectivity filter (i.e. 0.3 nm) and R0 is the size of potassium ion (≅0.2 nm) as a characteristic parameters for length. For interference patterns in the regions of our investigation, Figures (SI-1–SI-4) and Fig. 3, the different values of the distance between ion channels in relation with macroscopicity measure  are given in Table 2. Based on our results, the macroscopicity measure,
are given in Table 2. Based on our results, the macroscopicity measure,  is approximately a threshold for the distances between the ion channels for interference. It means the maximum double-slit distance between the slits is roughly d′ = 5.90 nm. The other d′ values obtained from quantum values
is approximately a threshold for the distances between the ion channels for interference. It means the maximum double-slit distance between the slits is roughly d′ = 5.90 nm. The other d′ values obtained from quantum values  (d′ = 0.18 nm) and
(d′ = 0.18 nm) and  (d′ = 0.38 nm) are not biologically feasible.
(d′ = 0.38 nm) are not biologically feasible.
 , indicates the region of quantum coherence for interference.
, indicates the region of quantum coherence for interference.
The values lower than  are classical values that show the interference pattern cannot be formed. The values from
are classical values that show the interference pattern cannot be formed. The values from  to
to  are quantum values which makes the interference pattern possible.
are quantum values which makes the interference pattern possible.
 is approximately a threshold for the distances between the ion channels for interference.
is approximately a threshold for the distances between the ion channels for interference.In the following section we will investigate the effect of biological environment on the quantum states of ions inside and outside of the selectivity filter. Then, we will obtain the coherence length of ions outside the selectivity filter.
Decoherence
The biological system is a very noisy and hot environment for quantum states and therefore there is a serious problem against quantum interference. In fact, “high vacuum is a prerequisite for all matter-wave interferometers”29. Additionally, in most interferometers vibrations are important source of dephasing29. Based on quantum mechanics and the decoherence model, every system rapidly entangle with the surrounding environment, which causes dephasing of their quantum states. Now, the question is: how ions can keep their quantum states during the whole crossing mechanism through ion channels while they face environmental particles in the hot, wet and noisy environment of the cell? Normally, crossing of each ion through ion channels takes at least 10–20 ns40 and therefore the decoherence time should be at least bigger than this time interval. This is rather a big decoherence time from quantum mechanical point of view and consequently it is a fundamental problem against the matter-wave interference of ions in biological environment. Considering the fact that even delocalized electrons (with much lighter mass than potassium ions) in low temperature quantum dots will dephase in ≈1 ns, how would it be feasible for massive potassium ions to remain coherent, showing interference patterns over 10–20 ns time scale? Tegmark already did very interesting calculations about decoherence timescales in neurons30 and obtained that the decoherence times are very short (≈10−20 s) for superposed ions to be effective in any cognitive processes. In fact, Tegmark didn’t consider the ion channel structure at all and his model was very simple as he only considered the superposition distance of ions equal to the membrane thickness. We already corrected the Tegmark’s calculations30 and obtained the decoherence timescales for a single ionic superposition in a single selectivity filter, which was in order of picoseconds, i.e. a hundered million times bigger than what he obtained17. Additionally, the Tegmark’s decoherence rate function leads to decoherence times that are directly proportional to temperature indicating his calculations do not address the temperature dependence of decoherence times correctly41,42. Despite the general controversial aspects of the decoherence model41,43,44 we would like to use the standard approach to investigate the decoherence timescales to see how fast an ionic superposition becomes decohered as a consequence of vibrations and environmental scattering.
Decoherence Formulation
Scattering with environmental particles is the most important reason of decoherence in every quantum system. The time evolution of a quantum superposed state can be shown by a reduced density matrix ρ(x, x′, t) in the following form

where the ρ(x, x′, t) is the density matrix of system in terms of time and position at time t, ρ(x, x′, 0) is the density matrix at t = 0, and F(x, x′, t) is the decoherence factor as follows45

where g(q) is the number density of scatterers with momentum q that ∫g(q)dq = N/V, in which N is the number of scatterers and V is the volume, v(q) = q/m′ is the speed of scatterers with mass m′ and momentum q, σtot(q) is the scattering cross section for the momentum q, and Γtot is the total scattering rate. Thus, we can rewrite the equation (5) in the following form

In general, the evolution of a quantum system regarding the decoherence can be written in the form of

where f(x, x′, t) is the delocalization rate of the superposed state in terms of time and position45. For scattering of environment particles that have a typical de Broigle wavelength λen, we have30,46

where Ω = σϕ in which σ is the scattering cross section and ϕ = nv is the flux, in which n is the density of scatterers and v is the velocity of object17, and Δx = |x − x′| is the superposition distance of the object. In the short wavelength limit, Δx ≫ λen, we can approximately rewrite30

On the other side, we have a time-dependent relation for density matrix  45. Thus, we have τdec = 1/Ω and as λ0 = h/mv (i.e. de Broglie’s relation for ion) we can finally write
45. Thus, we have τdec = 1/Ω and as λ0 = h/mv (i.e. de Broglie’s relation for ion) we can finally write

where here m is the mass of the ion, λ0 is the wavelength of the ion, n is the density of scatterers (n = N/V with N the number of scatterers and V is the volume).
In the long wavelength limit, λen ≫ Δx, the decoherence time in this case can be obtained from the equation (9) giving17

It is obvious that we are in the short wavelength limit because the superposition distance Δx = 5.9 is much bigger than the environmental de Broglie wavelength of particles, which are in the range 0.02–0.05 nm. Thus, we should calculate the decoherence times inside and outside the selectivity filter according to the equation 11. In this case, the decoherence time does not depend on the superposition distance.
Decoherence inside the selectivity filter
Assume that the selectivity filter is a cavity with volume V including N particles. The ion is the system which can be scattered by the particles in the cavity. The main scattering can happen between the ion and the particles in the cavity such as C=O bounds, water molecules and other ions (see Fig. 4). The decoherence time is obtained by letting n = N/V into the equation 11 as follows

The quantum states of ions in the selectivity filter may be decohered due to scattering with environmental particles and biomolecules in biological temperature.

The volume of the cavity as a cylinder is V = πr2L where r is the radius of the cavity and L is the length of the cavity. We already calculated the decoherence times in the filter17 based on the volume of the selectivity filter with radius r = 0.15 nm, however in this volume the C=O bonds cannot play the role as scatterers since they are behind the above radius. In fact, the whole radius in which carbonyl groups can be considered as scatterers in a cavity is approximately r = 0.4 nm. The scattering cross section can be approximately obtained from σ ≈ πa2 where a is the radius of the ion.
The results are shown in the Table 3. The results indicate that the decoherence time is around 100 picoseconds (i.e. ≈0.1 ns) inside the selectivity filter mainly due to scattering with C=O bonds. However, we should also consider that the selectivity filter backbone has vibrations that means the above cavity is vibrating. As we mentioned before, one of the important sources of dephasing can be due to channels vibrations in biological temperature. In fact, if ions are delocalized in the two channels then the vibrations of each channel can give random noise to the wave function and decohere it. We should notice here that the membrane fluctuations vary between 0.3–30 Hz47 and consequently the periods of vibrations vary between 0.03–3.3 sec which are much bigger than the pico– nano seconds timescales of the ionic translocation times. Therefore, at this scale the vibrations are not so effective. Indeed, the ion channel creates a fluctuating potential and coupling to phonons will be likely to dephase the quantumness in the system. If a particular system–environment interaction leads to dissipation in the system, then the strength of the system–environment interaction is a measure of the relaxation time. As the interaction strength decreases, the relaxation times become longer, and vice versa. The relation between the relaxation time and decoherence time for an object is45

where λdB is the thermal de Broglie wavelength of the ion. Based on our MD simulation, we let λdB ≈ 0.1 nm. The estimations for the relaxation times are shown in Table 3. It is seen that the relaxation times are mainly in order of nanoseconds but the decoherence times are in order of picoseconds. The obtained decoherence time (i.e. 0.1 ns) is a hundred times shorter than the ion translocation time in the filter (i.e. 10–20 ns), which makes the delocalization of ions in the two channels unlikely.
In the following subsection, we will obtain the decoherence times outside the channels.
Decoherence outside the selectivity filter
Basically each ion is bounded by eight water molecules outside the selectivity filter48. Considering thermal energy at room temperature, kBT = 4.11 × 10−21 J, the velocity of the hydrated water will be  m/s, thus the de Broglie wavelength of the ion is
m/s, thus the de Broglie wavelength of the ion is  nm. The speed of potassium ion (in biological temperature) that is not hydrated is
nm. The speed of potassium ion (in biological temperature) that is not hydrated is  m/s. Thus, its de Broglie wavelength is λdB = 0.02 nm.
m/s. Thus, its de Broglie wavelength is λdB = 0.02 nm.
The maximum superposition distance should be Δx = 5.9 nm for the maximum distance between the ion channels. Thus, for the superposition state of the ion outside of the selectivity filter and regarding the delocalization of the ion between the two ion channels we should consider the Eq. 11, which is used in the short wavelength limit (i.e. Δx ≫ λen) for scattering between the system and the environmental particles. For the ion density, let us write  where the density of water molecules
where the density of water molecules  is about (1 g/cm3)/(18 mp) ≈ 1023/cm3 = 1027/m3 and η is the relative concentration of ions (positive and negative combined)30. Typical ion concentrations during the resting state are [Na+] = 9.2 mmol/l = 92 × 1016/cm3 for inside and [Na+] = 120 mmol/l = 12 × 1018/cm3 outside the axon membrane and [K+] = 140 mmol/l = 14 × 1018/cm3 for inside and [K+] = 2.5 mmol/l = 25 × 1016/cm3 outside the axon membrane30.
is about (1 g/cm3)/(18 mp) ≈ 1023/cm3 = 1027/m3 and η is the relative concentration of ions (positive and negative combined)30. Typical ion concentrations during the resting state are [Na+] = 9.2 mmol/l = 92 × 1016/cm3 for inside and [Na+] = 120 mmol/l = 12 × 1018/cm3 outside the axon membrane and [K+] = 140 mmol/l = 14 × 1018/cm3 for inside and [K+] = 2.5 mmol/l = 25 × 1016/cm3 outside the axon membrane30.
Letting σ ≈ 10−20 m2 we obtain the decoherence times as in the Table 4. The results indicate that the superposition states can only survive about 17–53 picoseconds.
Discussion
In previous sections, we have obtained decoherence times of ions inside and outside the selectivity filter. The decoherence time inside the filter is about 100 ps, which is at least a hundred times shorter than 10–20 ns translocation time in the filter, making interference unlikely. However, the average velocity of ion in the filter is v ≈ 100 m/s and the length of selectivity filter is L ≈ 1.2 nm. We expect the translocation time to be t = L/v ≈ 10 ps but in reality the translocation time is 1000 times bigger than this time. It means that ions are trapped in the filter. We have already shown that there is a substantial “cooling effect” of ions in the filter by solving Schrodinger equation for ions and the electric potential in the selectivity filter19, indicating that ions are strongly trapped by carbonyl groups via electric interactions. This can make a weaker scattering with C=O bonds and therefore longer decoherence times. In fact, the present obtained decoherence times in this paper are based on the classical MD simulation that ignores the quantum effects. In order to have a better analysis, the ion movement in the filter should be investigated by quantum simulations or QM/MM (quantum mechanics/molecular mechanics) methods49.
Regarding the ps decoherence timescales outside the filter, we should calculate the coherence length as well. Based on the data for both isolated and hydrated ions and their velocities outside the selectivity filter as well as their decoherence times (see Table 4) we calculated the coherence length of ions,

where lc is the coherence length of ion, and vion is the velocity of ion.
The results indicate that the coherence length varies between 6.2–8.5 nm that is biologically feasible for interference but very close to the membrane. However, we should also compare the obtained coherence length with the mean free path (MFP) of ions regarding their collisions with water molecules around. The MFP is obtained as follows50

where πs2 is the effective collision area, and s is the size of water molecule (≈0.25 nm), which gives lMFP ≈ 5.1 nm. The obtained coherence lenghts and MFP are seen in the Table 5 in which the coherence lengths are close to the MFP, indicating that only one collision may change the direction of a superposed ion during the displacement of ions to the channels. This result makes interference plausible outside the filter despite short decoherence timescales.
Conclusion
In this paper, we investigated the possibility of quantum interference of ions through ion channels to see whether quantum interference can be the cause of selectivity in ion channels. Regarding the selectivity property, the main question is: what properties make the ion channels so selective? A convincing theory has to explain how a channel can permit passage of a particular ion, while excluding all ions of smaller diameter, including some that are much smaller51. To answer this question we suggested that the matter-wave interference can be a solution for selectivity, since interference happens between similar ions regarding the same size of slits. Additionally, quantum interference can make the transport of ions faster (i.e. 108 ions per second) as the ions don’t choose wrong channels. Here, we have investigated potassium ions passing the two neighboring KcsA ion channels via simulation with the physical double-slit experiment. Our results can be summerized as follows: (1) There is an estimated upper bound of 5.9 nm for inter-channel distance in order potentially to obtain quantum interference, (2) quantum states can cohere for about 100 picoseconds inside the selectivity filter, (3) coherence can be maintained for about 17–53 picoseconds outside the filter, and (4) the coherence length of an ion varies between 6.2–8.5 nm outside the filter, which are close distances to the membrane. Despite the feasibility of coherence length outside the filter, our present estimations inside the filter indicate that quantum interference seems unlikely. We have discussed above that ions can be trapped and cooled in the filter based on our previous quantum simulation19, thus the decoherence time can be increased due to weaker scattering effects.
Testing our hypothesis seems a difficult task in real conditions of the cell, but we would like to motivate other research teams to evaluate our approach experimentally. For example, it may be possible with the aid of mass spectroscopy techniques to see if any pattern can be formed. Also, it is useful to design a double slit experiment in the lab and simulate the similar conditions of the cell with the same size of channels in nano dimensions and investigate the interference patterns by counting ions at different places after the double-slit.
Additional Information
How to cite this article: Salari, V. et al. Quantum Interference and Selectivity through Biological Ion Channels. Sci. Rep. 7, 41625; doi: 10.1038/srep41625 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Ball, P. The dawn of quantum biology. Nature 474, 272–274 (2011).
Zimboras,. Z. et al. Quantum transport enhancement by time-reversal symmetry breaking. Sci. Rep. 3, 1–6 (2013).
Caruso, F. Universally optimal noisy quantum walks on complex networks. New J. Phys. 16, 1–17 (2014).
Perseguers, S., Lewenstein, M., Acin, A. & Cirac, J. I. Quantum random networks. Nat. Phys. 6, 539–543 (2010).
Fisher, M. P. A. Quantum cognition: The possibility of processing with nuclear spins in the brain. Annals of Physics 362, 593–602 (2015).
Vattay, G., Kauffman, S. & Niiranen, S. Quantum biology on the edge of quantum chaos. PLoS One 9, 1–6 (2014).
Lee, H., Cheng, Y. C. & Fleming, G. R. Coherence dynamics in photosynthesis: protein protection of excitonic coherence. Science 316, 1462–1465 (2007).
Brixner, T. et al. Two-dimensional spectroscopy of electronic couplings in photosynthesis. Nature 434, 625–628 (2005).
Engel, G. S. Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature 446, 782–786 (2007).
Ishizaki, A. & Fleming, G. R. Theoretical examination of quantum coherence in a photosynthetic system at physiological temperature. Proc. Natl. Acad. Sci. 106, 17255–17260 (2009).
Lambert, N. Quantum Biology. Nat. Phys. 9, 10–18 (2013)
Giese, B. et al. Direct observation of hole transfer through DNA by hopping between adenine bases and by tunnelling. Nature 412, 318–320 (2001).
Abbott, D., Davies, P. C. W. & Pati, A. K. Quantum Aspects of Life (World Scientific, 2008).
Vaziri, A. & Plenio, M. B. Quantum coherence in ion channels: resonances, transport and verification, New J. Phys. 12, 085001 (1–18) (2011).
Ganim, Z., Tokmakoff, A. & Vaziri, A. Vibrational excitons in ionophores: experimental probes for quantum coherence-assisted ion transport and selectivity in ion channels, New J. Phys. 13, 113030 (2–28) (2011).
Cifuentes, A. A. & Semiao, F. L. Quantum Model for a Periodically Driven Selectivity Filter in K+ Ion Channel. J. Phys. B: At. Mol. Opt. Phys. 47, 225503 (2014).
Salari, V., Moradi, N., Fazileh, F. & Shahbazi, F. Quantum decoherence time scales for ionic superposition states in ion channels. Phys. Rev. E 91, 032704 (1–6) (2015).
Bassereh, H., Salari, V. & Shahbazi, F. Noise assisted excitation energy transfer in a linear model of a selectivity filter backbone strand, J. Phys. Cond. Matter 27, 275102 (1–7) (2015).
Summhammer, J., Salari, V. & Bernroider, G. A quantum-mechanical description of ion motion within the confining potentials of voltage-gated ion channels. J. Integ. Neurosc. 11, 123–135 (2012).
Salari, V. et al. On the Classical Vibrational Coherence of Carbonyl Groups in the Selectivity Filter Backbone of the KcsA Ion Channel. J. Integ. Neurosc. 14 (2), 195–206 (2015).
Ngo, V., Stefanovski, D., Haas, S. & Farley, R. A. Non-Equilibrium Dynamics Contribute to Ion Selectivity in the KcsA Channel. PLoS One 9 (1), e86079 (2014).
MacKinnon, R. Potassium channels. FEBS Lett 555, 62–65 (2003).
Feynman, R. P., Leighton, R. B. & Sands, M. The Feynman Lectures on Physics Vol III. (Boston: Addison-Wesley, 1965).
Arndt, M. De Broglie’s meter stick: making measurements with matter-waves. Phys. Today 67, 30–36 (2014).
Juffmann, T., Ulbricht, H. & Arndt, M. Experimental methods of molecular matter-wave optics. Rep. Progr. Phys. 76, 1–28 (2013).
Eibenberger, S., Gerlich, S., Arndt, M., Mayor, M. & Tuxen, J. Matter-wave interference of particles selected from a molecular library with masses exceeding 10000 amu. Phys. Chem. Chem. Phys. 15, 14696–14700 (2013).
Gerlich, S. et al. Quantum interference of large organic molecules. Nat. Commun. 2, 1–5 (2011).
Arndt, M. et al. Wave-particle duality of C60 molecules. Nature 401, 680–682 (1999).
Arndt, M. et al. Atom Interferometry. eds Tino, G. M., Kasevich, M. A. IOS Press. 188, 89–141 (2014).
Tegmark, M. Importance of quantum decoherence in brain processes. Phys. Rev. E 61, 4194–4206 (2000).
Waxman, S. G., Kocsis, J. D. & Stys, P. K. In The Axon (Oxford University Press, 1995).
Kandel, E., Schwartz, J. & Jessell, T. Principles of Neural Science (McGraw-Hill, 2000).
Naeij, H. R. & Shafiee, A. Double-slit interference pattern for a macroscopic quantum system. Found. Phys. 46, 1634–1648 (2016).
Takagi, S. Macroscopic Quantum Tunneling. Cambridge University Press, New York (2005).
Merzbacher, E. Quantum Mechanics. Wiely, New York (1970).
Holland, P. R. The Quantum Theory of Motion. Cambridge University (1993).
Zecca, A. Gaussian wave packets passing through two slits: contribution of confinement and tunneling to the diffraction pattern. Adv. Studies Theor. Phys. 7, 287 (2013).
Zecca, A. Two-slit diffraction pattern for Gaussian wave packets. Int. J. Theor. Phys. 38, 911 (1999).
Zecca, A. & Cavalleri, G. Gaussian wave packets passing through a slit: a comparison between the predictions of the Schrodinger QM and of stochastic electrodynamics with spin. Nuovo Cimento 112.B, 1 (1997).
Berneche, S. & Roux, B. A microscopic view of ion conduction through the K+ channel. Proc. Natl. Acad. Sci. 100, 8644–8648, (2003).
Hagan, S., Hameroff, S. R. & Tuszynski, J. A. Quantum computation in brain microtubules: Decoherence and biological feasibility. Phys. Rev. E 65, 061901 (1–11) (2002).
Salari, V., Tuszynski, J., Rahnama, M. & Bernroider, G. Plausibility of quantum coherent states in biological systems. J. Phys. Conf. Ser. 306(1), 012075 (2011).
Adler, S. L. Why decoherence has not solved the measurement problem: a response to PW Anderson. Stud. Hist. Phil. Sci. B 34, 135–142 (2003).
Bassi, A. et al. Models of wave-function collapse, underlying theories, and experimental tests. Rev. Mod. Phys. 85, 471–527 (2013).
Schlosshauer, M. A. Decoherence and the Quantum-To-Classical Transition Springer (2007).
Tegmark, M. Apparent wave function collapse caused by scattering, Found. Phys. Lett. 6(6), 571–590 (1993).
Tuvia, Sh. et al. Cell membrane fluctuations are regulated by medium macroviscosity: Evidence for a metabolic drivingforce. Proc Natl Acad Sci 94(10) 5045–5049 (1997).
Zhou, Y., Morais-Cabral, J. H., Kaufman, A. & MacKinnon, R. Chemistry of ion coordination and hydration revealed by a K & plus; channel-Fab complex at 2.0 Angstrom resolution. Nature 414, 43–48 (2001).
Brunk, E. & Rothlisberger, U. Mixed Quantum Mechanical/Molecular Mechanical Molecular Dynamics Simulations of Biological Systems in Ground and Electronically Excited States. Chemical Reviews. 115(12), 6217–6263 (2015).
Rohlf, J. W. Modern Physics from a to Z0 Wiley (1994).
Pollack, G. H. Cell electrical properties: reconsidering the origin of the electrical potential. Cell. Biol. Int. 9999, 1–6 (2014).
Acknowledgements
The authors thank Dr. Rouhollah Ghobadi from University of Calgary for valuable comments and Dr. Saleh Rahimi-Keshari from University of Queensland for helpful discussions.
Author information
Authors and Affiliations
Contributions
V.S. proposed the idea, H.N., A.Sh. did the macroscopicity calculations, V.S. did the MD simulation and the decoherence calculations, V.S., H.N., and A.Sh. contributed to the development and completion of the idea, analyzing the results and discussions, V.S., H.N., and A.Sh. participated in writing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Salari, V., Naeij, H. & Shafiee, A. Quantum Interference and Selectivity through Biological Ion Channels. Sci Rep 7, 41625 (2017). https://doi.org/10.1038/srep41625
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep41625
This article is cited by
-
Quantum coherence on selectivity and transport of ion channels
Scientific Reports (2022)
-
Information transmission in microbial and fungal communication: from classical to quantum
Journal of Cell Communication and Signaling (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
