
Abstract
Age-related macular degeneration (AMD) is the most frequent cause of blindness in the elderly. There is evidence that nutrition, inflammation and genetic risk factors play an important role in the development of AMD. Recent studies suggest that the composition of the intestinal microbiome is associated with metabolic diseases through modulation of inflammation and host metabolism. To investigate whether compositional and functional alterations of the intestinal microbiome are associated with AMD, we sequenced the gut metagenomes of patients with AMD and controls. The genera Anaerotruncus and Oscillibacter as well as Ruminococcus torques and Eubacterium ventriosum were relatively enriched in patients with AMD, whereas Bacteroides eggerthii was enriched in controls. Patient’s intestinal microbiomes were enriched in genes of the L-alanine fermentation, glutamate degradation and arginine biosynthesis pathways and decreased in genes of the fatty acid elongation pathway. These findings suggest that modifications in the intestinal microbiome are associated with AMD, inferring that this common sight threatening disease may be targeted by microbiome-altering interventions.
Similar content being viewed by others

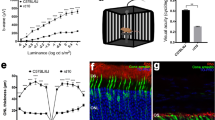

Introduction
Age-related macular degeneration (AMD; Fig. 1a) is the leading cause of severe visual impairment in the elderly population of industrialized countries. AMD has become a significant global health issue1, and the prevalence of AMD is projected to grow by 50 percent in the next decades2. It is a complex disease that is caused by a combination of environmental and genetic factors. Several studies have shown that diet is a major modifiable risk factor for AMD development3,4,5,6,7,8. Intestinal microbial organisms and their genomes, collectively termed microbiome, play a major role in the digestion of food and in this way influence the global metabolism of their host9. The intestinal microbiome forms a complex ecosystem of up to 100 trillion microbes and shows substantial diversity of gut communities among individuals10. However, changes in microbiome composition can potentially modulate or alternatively stress host metabolism and may act as sources of inflammation and disease11. Recent findings have identified the intestinal microbiome as a contributor to metabolic diseases such as atherosclerosis, which is associated with lipid accumulation and inflammation in the arterial wall12. Furthermore, a recent report has suggested that the intestinal microbiome may trigger autoimmune response in the eye through activation signals to retina specific T cells13. Given the link between AMD and diet, the composition of the intestinal microbiome may also influence AMD development and progression. This link has recently been substantiated in an animal model, where alterations in the intestinal microbiome have resulted in exacerabation of choroidal neovascularization, a hallmark of neovascular AMD14. Neovascular AMD is caused by choroidal neovascularization, which often results in exudation and hemorrhage, leading to photoreceptor damage and central visual loss.

(a) Representative color photographs and optical coherence tomography (OCT) images of a patient with neovascular age-related macular degeneration (AMD; left) and an age-matched control (right). (b) Illustration of our bioinformatics pipeline for analysing metagenomic data and its relation to neovascular AMD. Sequencing reads from stool probes were generated with high throughput sequencing and subjected to quality control. Species abundances were estimated by alignment of high quality reads to clade-specific marker sequences using the Metagenomic Phylogenetic Analysis (MetaPhlAn) tool. To describe the functional profile of the intestinal microbiome, the HMP (Human Microbiome Project) Unified Metabolic Analysis Network (HUMAnN) was used. The linear discriminant analysis (LDA) effect size (LEfSe) algorithm was applied to identify features significantly different between patients and controls. To provide a global analysis of microbial species and pathway abundance principal component analysis (PCA) using ade4 were performed.
To address this issue, we investigated whether compositional and functional diversity of the intestinal microbiome is associated with AMD development and progression. To this end, we sequenced the intestinal metagenomes of patients with recent onset neovascular AMD and age- and sex-matched controls without AMD. For profiling microbial communities, we used the Metagenomic Phylogenetic Analysis (MetaPhlAn) tool15 and to infer the metabolic potential of the identified microbes, the HMP (Human Microbiome Project16) Unified Metabolic Analysis Network (HUMAnN)17 was applied. The data revealed an enrichment of Oscillibacter, Anaerotruncus, Eubacterium ventriosum and Ruminococcus torques in AMD patients versus Bacteroides eggerthii in controls. Our findings may have important implications for the prevention of this sight threatening disease as the intestinal microbiome may be modified by probiotics and antibiotics.
Results
Taxonomic characterization of the intestinal microbiome
To address whether the intestinal microbiome is associated with neovascular AMD, we sequenced the gut metagenomes of 12 patients with recent onset signs of neovascularization due to AMD and 11 age- and sex-matched controls without any signs of AMD (Table 1). In total, we generated 557 million 100 bp paired-end reads with an average insert size of 350 bp (on average, 24 ± 0.1 (s.d.) million reads per sample).
After trimming and filtering (see Methods), we obtained about 20 million non-human high-quality reads per sample for further analysis. On average, 4.6 ± 1.2% (s.d.) of the reads of a sample could be aligned to any marker sequence. This is by a factor of ten less than described in previous metagenomic studies using Illumina reads12,18. This discrepancy is due to the fact that MetaPhlAn maps reads against a catalogue of marker sequences including about 4% of the sequenced microbial genomes15. The majority of mapped reads were bacterial (99.2 ± 1.6% in patients and 99.7 ± 0.5% (s.d.) in controls). The microbiome composition was dominated by the phyla Bacteriodetes (65.3 ± 16.5% in AMD and 74.7 ± 12.6% (s.d.) in controls) and Firmicutes (29.0 ± 14.4% in AMD and 20.4 ± 8.9% (s.d.) in controls), followed by Actinobacteria (2.1 ± 3.2% in AMD and 1.6 ± 3.6% (s.d.) in controls) and Proteobacteria (1.6 ± 1.7% in AMD and 2.4 ± 3.0% (s.d.) in controls). Bacteroidia (65.3 ± 16.5% in AMD and 74.7 ± 12.6% (s.d.) in controls) and Clostridia (27.1 ± 13.4% in AMD and 19.4 ± 8.8% (s.d.) in controls) were shown to be the most abundant classes in our cohort (Fig. 2). This composition is consistent with previous observations12,19,20. The genera Bacteroides (36.5 ± 14.6% in AMD and 40.8 ± 23.1% (s.d.) in controls) and Alistipes (12.6 ± 6.4% in AMD and 13.8 ± 11.9% (s.d.) in controls) dominated the microbiome, followed by Eubacterium with a high intersubject variation (8.9 ± 8.6% in AMD and 5.3 ± 6.9% (s.d.) in controls). A total of 12 species were detected in all 23 samples, making up the core microbiome in our cohort, in agreement with previous observations18. Bacteroides uniformis was the most abundant species with 8.7 ± 6.0% in AMD and 6.1 ± 5.7% (s.d.) in controls, followed by Alistipes onderdonkii (4.4 ± 4.7% in AMD and 7.4 ± 10.5% (s.d.) in controls), Subdoligranulum species (4.8 ± 3.9% in AMD and 4.4 ± 6.1% (s.d.) in controls) and Alistipes putredinis (4.8 ± 4.6% in AMD and 4.1 ± 3.9% (s.d.) in controls).

Mean relative abundance of the major microbial classes among individuals of the cohort. Red are patients (age-related macular degeneration (AMD), n = 12), green are controls (n = 11).
Distinct microbial composition
A principal component analysis with the health status as grouping variable revealed that differences in abundance of microbial species separated the patient group from the control group (Fig. 3a). To identify the taxa differing in relative abundance between AMD and controls, LEfSe was applied on the previously identified microbiome. The genera Anaerotruncus and Oscillibacter (and the family Oscillospiraceae) as well as Ruminococcus torques and Eubacterium ventriosum were enriched in AMD, whereas Bacteroides eggerthii was enriched in controls (p < 0.05, Kruksal-Wallis test; Fig. 3b,c).

(a) Principal component analysis of microbial species abundance using health status as grouping variable. The relation between microbial abundance and health status was assessed using Monte Carlo simulations with 10,000 iterations by which a p value was calculated. (b,c) Identification of taxonomic features relevant in neovascular AMD using LefSe. Cladogram for taxonomic representation of significant differences among AMD and controls. The diameter of each circle is proportional to its abundance (b). Histogram of the LDA scores for differentially abundant taxonomic features among groups. The threshold on the logarithmic LDA (linear discriminant analysis) score for discriminative features was set to 2.0 (C). (d) Box plots representing the mean abundance +/−SD of bacterial genera and species associated with AMD (Kruksal-Wallis test, p ≤ 0.05). Red are patients (AMD, n = 12), green are controls (C, n = 11).
Metabolic functions of altered intestinal microbiome
To describe the metabolic features of the microbiome, we applied HUMAnN on each sample separately based on the taxonomic profiles from MetaPhlAn. Resulting organism-specific gene hits were functionally assigned to pathways using MinPath and their relative abundances were assessed. From the totally 2193 assigned organism-specific pathways, 253 pathways were detected in all samples making up the core metabolic functions in our cohort. A principal component analysis with the health status as grouping variable revealed that the difference in abundance of metabolic pathways separated the patient group from the control group (Fig. 4a). Applying LEfSe, 20 abundant pathways (i.e. occurring in at least 11 samples) were identified that differ between AMD patients and controls (Fig. 4b). Based on the distribution of pathway abundances in each sample with empiric p values > 0.05, we identified four differentially abundant pathways (Fig. 4c) using an empiric null distribution. L-alanine fermentation (GO 0019652), glutamate degradation (GO 0019671) and arginine biosynthesis (GO 0006525) pathways were up-regulated in the intestinal microbiome of AMD patients, whereas fatty acid elongation pathway (GO 00062) was enriched in the microbiome of controls (see Supplementary Fig. S1).

(a) Principal component analysis of microbial pathway abundance with health status as instrumental variable. The relation between pathway abundance and health status was assessed using Monte Carlo simulations with 10,000 iterations by which a p value was calculated. (b) Metabolic pathways that are differentially abundant among subjects with AMD and controls, ranked according to the effect size (LDA score, linear discriminant analysis score). (c) Box plots representing the mean abundance +/−SD of pathways associated with AMD (empiric p > 0.05). Red is patients (AMD, n = 12), green is controls (C, n = 11).
Discussion
In this study, we identified several compositional and functional variations of the intestinal microbiome that may be related to the development of neovascular AMD. There is significant geographic and ethnic variability in the prevalence of AMD2. Although genetic susceptibility has been shown to play a substantial role in the pathogenesis of AMD and may contribute to the ethnic differences in its prevalence21, this variability may be further influenced by differences in the intestinal microbiome and diet, respectively. The intestinal microbiome consists of a complex community of microbes that impact normal physiology and susceptibility to disease mediated by inflammatory molecules such as lipopolysaccharides and peptidoglycans, which may contribute to metabolic disease22,23. The intestinal microbiome landscape seems to be fairly distinct and temporally stable in each individual. This suggests that a stable equilibrium exists for the microbiome. Furthermore, twins have more comparable microbiome compositions than unrelated individuals, suggesting that genetics may influence the microbiome composition. On the other hand, environmental factors may have a similar impact on microbiome composition, as monozygotic and dizygotic adult twins have equally similar microbiomes11. Recent studies have shown that the European microbiome is dominated by the Bacteroides enterotype, whereas the African microbiome was dominated by the Prevotella enterotype24.
We observed associations between neovascular AMD and microbiome composition at the taxonomical levels of bacterial genera and species. Furthermore, principal component analysis (PCA) on microbial species abundance confirmed a separation of patients and controls by microbiome composition (Fig. 3a). First, the relative abundance of Bacteroides eggerthii was increased in healthy age- and sex-matched controls. Bacteroides genera have been shown to play a major role in carbohydrate fermentation resulting in a pool of volatile fatty acids that are reabsorbed through the intestinal mucosa, providing a significant source of energy to the host25. Bacteroides can produce polysaccharide A (PSA), which may be involved in ligand-receptor interaction, modulating the immune response to pathogens. It has been indeed shown that PSA derived from Bacteroides spp. protect from experimental autoimmune encephalitis26. Secondly, the relative abundances of species such as Ruminococcus torques, a gram positive bacterium with mucin degradation capacities and Oscillibacter, which has been associated with high fat diet27, were increased in AMD patients. An increased Oscillibacter population has been shown to result in an increased gut permeability presumably mediated by a reduction in the mRNA expression tight junctions including ZO-128. An increased relative abundance of Anaerotruncus species has been associated with aging and age-associated inflammation with elevated pro-inflammatory chemokines in a mouse model29. A similar finding has been reported for Eubacterium ventriosum spp. in humans which has been associated with elevated IL-6 and IL-8 cytokine levels30.
High-fat diets have been shown to exacerbate choroidal neovascularization in a mouse model by increasing the relative abundance of Firmicutes. These findings were associated with heightened intestinal permeability and chronic low-grade inflammation with elevated production of IL-6, IL-1b, TNF-a, and VEGF-A, cytokines that have been associated with the development of neovascular AMD14. A similar shift of relative abundance in Firmicutes at the expense of Bacteroidetes was observed in our cohort, with a relative abundance of Firmicutes of 29.0% in AMD and 20.4% in the control group.
Functional annotation analyses indicated that specific genes involved in individual metabolic pathways are enriched or decreased in patients with neovascular AMD. Although these genes allow to identify associated pathways and therefore can predict functional capabilities, quantification of messenger RNA and metabolic profiling to confirm functional differences are needed to further confirm these associations.
However, PCA on gene and pathway abundance revealed a separation of patients and controls by functional features (Fig. 4a). Our data revealed a decline in bacteria responsible for fatty acid elongation and an increase in L-alanine fermentation, glutamate degradation and arginine biosynthesis in patients with neovascular AMD. In the last decade, it has become evident that nutritional factors may influence the progression of AMD. A recent study has observed a decrease of some n-3 very long chain polyunsaturated fatty acids in the retina of eyes with early and intermediate AMD31. As such bioavailability of long chain polyunsaturated fatty acids is likely to have an impact on retinal physiology and may contribute to AMD. Glutamate is the main excitatory neurotransmitter in the retina and decreased availability of glutamate has been shown to result in deficient neurotransmission in the retina32. Furthermore, in patients with reduced ornithin aminotransferase activity increased levels of arginine are associated with progressive chorioretinal atrophy, suggesting an important role for arginine in the development of retinal degeneration33.
The best-validated supplementation therapy for AMD is the age-related eye disease study (AREDS) formulation, which contains antioxidant vitamin C and vitamin E, beta-carotene, copper and zinc. This combination has been shown to reduce risk toward progression of advanced AMD34. The bioavailability of several of these components may be influenced by the microbiome composition. This is supported by the large effect of the microbiome on mammalian plasma biochemistry which has been demonstrated by comparing plasma extracts from germ-free mice with samples from conventional mice35. Importantly, generation of certain antioxidants such as indole-3-propionic acid was shown to be completely dependent on the presence of intestinal microflora. Furthermore, it has been suggested that zinc absorption may be influenced by zinc competition between microbiome in the gastrointestinal tract of a host36.
This study cannot provide evidence for direct causal effects of an altered intestinal microbiome on the development of neovascular AMD. Study limitations are the small sample size which may not represent the entire AMD population, and furthermore, there are possible confounders due to genetic background and nutritional habit of patients and controls. However, if a specific taxonomic group or a metagenomic cluster annotated to a specific metabolic pathway is ultimately shown to be causally related to neovascular AMD, then long-term dietary interventions may allow modulation of an individual’s microbiome to avoid disease. The interplay between microbiome and the innate immune system, e.g. the complement system may provide a link between host genome and microbiome and remains an area of active investigation.
Our study suggests that the intestinal microbiome may play an important role in the development of neovascular AMD and may constitute a link between nutrition and development of this common disease.
Methods
Study design and recruitment
Participants (n = 23) were recruited from the Department of Ophthalmology of the University Hospital Bern (Inselspital), Switzerland. This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the Canton of Bern (ClinicalTrials.gov: NCT02438111). After receiving oral and written information, all participants gave written informed consent to participate in the study. All participants were subjected to an ophthalmic examination including standard fundus color photography and optical coherence tomography. Patients (n = 12) had clinically confirmed recent onset of neovascular AMD and were 50 years of age or older. The control group (n = 11) was selected to represent an age- and sex-matched group with no signs of AMD. We tested for differences in a range of demographic values between the two groups using either Welch’s t test (for age and BMI) or Fisher’s exact test (for gender and smoking) in R v.3.2.137. Further exclusion criteria for both groups were problems with the bowel movement, chronic inflammatory and gastrointestinal disease (including previous surgery in the gastrointestinal tract), recent history of use of systemic antibiotics within the last three months and opacities of ocular media.
Sequencing and data quality control
Stool samples were brought chilled to the study center where they were immediately processed and frozen at −20 °C. Metagenomic DNA was isolated from 0.25 g of stool sample using the PowerSoil® DNA Isolation kit (Mo Bio Laboratories, Carlsbad, USA) according to the manufacturer’s protocol. The DNA was sent to BGI Europe (Copenhagen N, Denmark) for paired-end shotgun sequencing with up to ten samples pooled in one lane using the Illumina HiSeq 2000 platform. For library preparation, purified DNA was sheared by nebulization. The resulting overhangs were converted into blunt ends using T4 DNA polymerase, Klenow Fragment and T4 Polynucleotide kinase. After adding an ‘A’ base to the 3′ end of the blunt phosphorylated DNA fragments, adapter ligation was performed. Too short fragments were removed with Ampure beads and the resulting library was used for sequencing following standard pipelines of the Illumina platform, generating 100 bp paired-end reads. The reads were quality filtered with Trimmomatic v.0.32 as follows38: Illumina adapter sequences were removed and read ends were clipped off if the average base quality in a 4 bp window slid along the length of the read dropped below 15. We retained only reads of at least 80 bp after trimming. To remove sequences of human origin, all reads were mapped to the human reference genome hg19 using Bowtie2 v.2.2.439 and only the unmapped reads were used for further analysis.
Taxonomical analysis
The bioinformatics pipeline used for analysing the metagenomic data is illustrated in Fig. 1b. For microbial profiling, the high-quality non-human reads were mapped against a set of clade-specific markers (spanning bacteria and archaea at the species level or higher) using the Metagenomic Phylogenetic Analysis (MetaPhlAn) tool v.2.0 and the marker database v.2015 using default settings. In order to provide the relative abundance of each taxonomic unit, Bowtie2 v.2.2.4 was used for alignment (option “very-sensitive”) followed by normalization of the total number of reads in each clade by the nucleotide length of its marker.
The R package ade440 performing PCA was used to provide the global analysis of microbial species abundances between AMD and controls as described in http://www.sthda.com/english/wiki/ade4-and-factoextra-principal-component-analysis-r-software-and-data-mining. PCA was performed with scaled values on relative abundances of microbial species identified by MetaPhlAn (Fig. 1b). A visualization of the individual samples grouped by the classes (case and control) is provided in Fig. 3a. Monte Carlo simulations with 10,000 iterations were performed on the between-groups inertia percentage by which a p value was calculated in Fig. 3a.
To identify taxa with significantly different relative abundances in AMD and controls, relative abundances from the different samples were merged and the linear discriminant analysis effect size (LEfSe) algorithm41 was applied with default settings (p < 0.05 based on Kruksal-Wallis test and LDA score ≥ 2).
Functional profiling
To describe the metabolic potential of the identified microbes, the HMP (Human Microbiome Project)16 Unified Metabolic Analysis Network (HUMAnN2 v.0.2.1) was used with default settings17. HUMAnN investigates the presence/absence and the relative abundance of gene families and pathways in each sample to provide a functional interpretation of the metagenomic sequences. HUMAnN was run for each sample separately using the taxonomic profiles from MetaPhlAn. For nucleotide-level searches, Bowtie2 v.2.2.4 was used to map reads to the functionally annotated pangenome database ChocoPhlAn. All unmapped reads were used for translated searches against the universal protein reference database UniRef5042 using Diamond v. 0.7.943. The resulting organism-specific gene hits were assigned to pathways using MinPath v.1.244, finally providing a set of pathways and their relative abundances. To provide an empiric null distribution, 10,000 permutations were performed, randomly assigning the pathways to the two groups, and for each permutation the difference of the means was calculated, providing an empiric p value for each feature (Fig. 4c).
The R package ade440 performing PCAwas used to provide the global analysis of pathway abundance between AMD and controls as described in http://www.sthda.com/english/wiki/ade4-and-factoextra-principal-component-analysis-r-software-and-data-mining. PCA was performed with scaled values on relative abundance of genes and pathways identified by HUMAnN (Fig. 1b). A visualization of the individuals grouped by case and control is provided in Fig. 4a. Monte Carlo simulations with 10,000 iterations were performed on the between-groups inertia percentage by which a p value was calculated in Fig. 4a.
To identify differences between AMD and controls, LEfSe was applied to all pathways that had been found in at least 11 of the 23 samples using default settings. A difference was considered to be statistically significant if p < 0.05 (Kruksal-Wallis test) and LDA score ≥ 2.
Additional Information
Accession codes: Gut microbiome sequencing reads have been deposited in the European Nucleotide Archive under accession number PRJEB13835.
How to cite this article: Zinkernagel, M. S. et al. Association of the Intestinal Microbiome with the Development of Neovascular Age-Related Macular Degeneration. Sci. Rep. 7, 40826; doi: 10.1038/srep40826 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Friedman, D. S. et al. Prevalence of age-related macular degeneration in the United States. Archives of ophthalmology 122, 564–572, doi: 10.1001/archopht.122.4.564 (2004).
Wong, W. L. et al. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. Lancet Glob Health 2, e106–116, doi: 10.1016/S2214-109X(13)70145-1 (2014).
Age-Related Eye Disease Study Research, G. et al. The relationship of dietary carotenoid and vitamin A, E, and C intake with age-related macular degeneration in a case-control study: AREDS Report No. 22. Archives of ophthalmology 125, 1225–1232, doi: 10.1001/archopht.125.9.1225 (2007).
Ersoy, L. et al. Nutritional risk factors for age-related macular degeneration. BioMed research international 2014, 413150, doi: 10.1155/2014/413150 (2014).
SanGiovanni, J. P. et al. The relationship of dietary lipid intake and age-related macular degeneration in a case-control study: AREDS Report No. 20. Archives of ophthalmology 125, 671–679, doi: 10.1001/archopht.125.5.671 (2007).
Seddon, J. M. Multivitamin-multimineral supplements and eye disease: age-related macular degeneration and cataract. The American journal of clinical nutrition 85, 304S–307S (2007).
Seddon, J. M. et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group. Jama 272, 1413–1420 (1994).
Chew, E. Y. et al. Effect of Omega-3 Fatty Acids, Lutein/Zeaxanthin, or Other Nutrient Supplementation on Cognitive Function: The AREDS2 Randomized Clinical Trial. Jama 314, 791–801, doi: 10.1001/jama.2015.9677 (2015).
Kirschner, R., Hsu, T., Tuan, N. N., Chen, C. L. & Huang, S. L. Characterization of Fungal and Bacterial Components in Gut/Fecal Microbiome. Curr Drug Metab 16, 272–283 (2015).
Eckburg, P. B. et al. Diversity of the human intestinal microbial flora. Science 308, 1635–1638, doi: 10.1126/science.1110591 (2005).
Turnbaugh, P. J. et al. A core gut microbiome in obese and lean twins. Nature 457, 480–484, doi: 10.1038/nature07540 (2009).
Karlsson, F. H. et al. Symptomatic atherosclerosis is associated with an altered gut metagenome. Nature communications 3, 1245, doi: 10.1038/ncomms2266 (2012).
Horai, R. et al. Microbiota-Dependent Activation of an Autoreactive T Cell Receptor Provokes Autoimmunity in an Immunologically Privileged Site. Immunity 43, 343–353, doi: 10.1016/j.immuni.2015.07.014 (2015).
Andriessen, E. M. et al. Gut microbiota influences pathological angiogenesis in obesity-driven choroidal neovascularization. EMBO molecular medicine, doi: 10.15252/emmm.201606531 (2016).
Segata, N. et al. Metagenomic microbial community profiling using unique clade-specific marker genes. Nat Methods 9, 811–814, doi: 10.1038/nmeth.2066 (2012).
Human Microbiome Project, C. A framework for human microbiome research. Nature 486, 215–221, doi: 10.1038/nature11209 (2012).
Abubucker, S. et al. Metabolic reconstruction for metagenomic data and its application to the human microbiome. PLoS Comput Biol 8, e1002358, doi: 10.1371/journal.pcbi.1002358 (2012).
Qin, J. et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464, 59–65, doi: 10.1038/nature08821 (2010).
Arumugam, M. et al. Enterotypes of the human gut microbiome. Nature 473, 174–180, doi: 10.1038/nature09944 (2011).
Tap, J. et al. Towards the human intestinal microbiota phylogenetic core. Environ Microbiol 11, 2574–2584, doi: 10.1111/j.1462-2920.2009.01982.x (2009).
Seddon, J. M., Santangelo, S. L., Book, K., Chong, S. & Cote, J. A genomewide scan for age-related macular degeneration provides evidence for linkage to several chromosomal regions. Am J Hum Genet 73, 780–790, doi: 10.1086/378505 (2003).
Cani, P. D. et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56, 1761–1772, doi: 10.2337/db06-1491 (2007).
Backhed, F., Ley, R. E., Sonnenburg, J. L., Peterson, D. A. & Gordon, J. I. Host-bacterial mutualism in the human intestine. Science 307, 1915–1920, doi: 10.1126/science.1104816 (2005).
De Filippo, C. et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA 107, 14691–14696, doi: 10.1073/pnas.1005963107 (2010).
Hooper, L. V., Midtvedt, T. & Gordon, J. I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu Rev Nutr 22, 283–307, doi: 10.1146/annurev.nutr.22.011602.092259 (2002).
Ochoa-Reparaz, J. et al. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol 3, 487–495, doi: 10.1038/mi.2010.29 (2010).
Lam, Y. Y. et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS One 7, e34233, doi: 10.1371/journal.pone.0034233 (2012).
Cani, P. D. et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57, 1470–1481, doi: 10.2337/db07-1403 (2008).
Conley, M. N. et al. Aging and serum MCP-1 are associated with gut microbiome composition in a murine model. PeerJ 4, e1854, doi: 10.7717/peerj.1854 (2016).
Biagi, E. et al. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS One 5, e10667, doi: 10.1371/journal.pone.0010667 (2010).
Liu, A., Chang, J., Lin, Y., Shen, Z. & Bernstein, P. S. Long-chain and very long-chain polyunsaturated fatty acids in ocular aging and age-related macular degeneration. J Lipid Res 51, 3217–3229, doi: 10.1194/jlr.M007518 (2010).
Bui, B. V. et al. Glutamate metabolic pathways and retinal function. Journal of neurochemistry 111, 589–599, doi: 10.1111/j.1471-4159.2009.06354.x (2009).
Simell, O. & Takki, K. Raised plasma-ornithine and gyrate atrophy of the choroid and retina. Lancet 1, 1031–1033 (1973).
Chew, E. Y., Lindblad, A. S. & Clemons, T. & Age-Related Eye Disease Study Research, G. Summary results and recommendations from the age-related eye disease study. Archives of ophthalmology 127, 1678–1679, doi: 10.1001/archophthalmol.2009.312 (2009).
Wikoff, W. R. et al. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc Natl Acad Sci USA 106, 3698–3703, doi: 10.1073/pnas.0812874106 (2009).
Gielda, L. M. & DiRita, V. J. Zinc competition among the intestinal microbiota. MBio 3, e00171–00112, doi: 10.1128/mBio.00171-12 (2012).
Team, R. D. C. R: A language and environment for statistical computing. R Foundation for Statistical Computing (2008).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120, doi: 10.1093/bioinformatics/btu170 (2014).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat Methods 9, 357–359, doi: 10.1038/nmeth.1923 (2012).
Stéphane Dray, A.-B. e. D. The ade4 Package: Implementing the Duality Diagram for Ecologists. Journal of Statistical Software 22 (2007).
Segata, N. et al. Metagenomic biomarker discovery and explanation. Genome Biol 12, R60, doi: 10.1186/gb-2011-12-6-r60 (2011).
Wu, C. H. et al. The Universal Protein Resource (UniProt): an expanding universe of protein information. Nucleic acids research 34, D187–191, doi: 10.1093/nar/gkj161 (2006).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using DIAMOND. Nat Methods 12, 59–60, doi: 10.1038/nmeth.3176 (2015).
Ye, Y. & Doak, T. G. A parsimony approach to biological pathway reconstruction/inference for genomes and metagenomes. PLoS Comput Biol 5, e1000465, doi: 10.1371/journal.pcbi.1000465 (2009).
Author information
Authors and Affiliations
Contributions
M.S.Z., D.C.Z., G.M.F. and S.W. conceived and designed the project. A.B.L. and C.R.L. performed the experiments. D.C.Z. and I.K. analysed the sequence data. All authors contributed writing and editing the manuscript. M.S.Z. and D.C.Z. contributed equally to the study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zinkernagel, M., Zysset-Burri, D., Keller, I. et al. Association of the Intestinal Microbiome with the Development of Neovascular Age-Related Macular Degeneration. Sci Rep 7, 40826 (2017). https://doi.org/10.1038/srep40826
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40826
This article is cited by
-
What is the impact of microbiota on dry eye: a literature review of the gut-eye axis
BMC Ophthalmology (2024)
-
Housing RCS rats under specific pathogen-free conditions mildly ameliorates retinal degeneration and alters intestine microbiota
Scientific Reports (2024)
-
Long-term intake of Lactobacillus helveticus enhances bioavailability of omega-3 fatty acids in the mouse retina
npj Biofilms and Microbiomes (2024)
-
Clinical trials targeting the gut-microbiome to effect ocular health: a systematic review
Eye (2023)
-
Combination of Lactobacillus fermentum NS9 and aronia anthocyanidin extract alleviates sodium iodate-induced retina degeneration
Scientific Reports (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
