
Abstract
A new, extraordinary palaeontinid Hallakkungis amisanus Nam, Wang & Szwedo, gen. et sp. nov., from the Upper Triassic of the Amisan Formation in Boryeong City, Korea is described. It is the first Palaeontinidae from Korea. The newly described taxon displays a mosaic of characters present in presumed ancestors of this insect family and some highly advanced features.
Similar content being viewed by others

Introduction
The first record of the Late Triassic palaeontinid from Korea presented below comes from the upper deposits of the Nampo Group. Palaeontinidae is an extinct family of cicadomorphan hoppers (Hemiptera: Cicadomorpha), superficially resembling huge moths, which existed since the Triassic to end of the Late Cretaceous in Europe, Asia, and South America. Palaeontinids had large bodies covered with bristles (setae), small heads and broad wings. The host plants of these plant-sucking insects have been assumed to be ginkgophytes based on the geographic distribution of both groups.
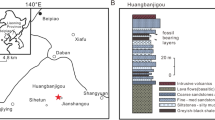
The Nampo Group is a non-marine deposit and mainly distributed in Boryeong City and Cheonyang-gun County, the western part of Chungcheongnam-do Province in Korea. The Nampo Group is distributed in the Chungnam Basin of southwestern part of the Chungcheongnam-do, and consists of a 3,000 m-thick sequence of terrestrial sediments1,2,3. The Chungnam Basin contains several subbasins4,5; the insectiferous locality is located in the Oseosan Subbasin (Fig. 1a–d). The fossil comes from the Upper Triassic, Norian (ca. 227 – ca. 208.5 Mya) Amisan Formation, which is one of five formations in the Nampo Group6.

(a) location of the fossil site (boxed); (b) close-up of the southern Korean Peninsula showing the study area (boxed); (c) geological map of study area; (d) Strato-sedimentological interpretation of the Nampo Group in the Oseosan subbasin. Figures in inserts a and b were made with data from Natural Earth (http://www.naturalearthdata.com); coastline in c adopted from Open Street Map, with cartography map tiles licensed under CC BY-SA (www.openstreetmap.org/copyright). Figure created using ArcGIS 10.1, CorelDraw X7 and Adobe Photoshop CS software packages.
Abundant fossil plants have been found from the Amisan and the Baegunsa formations, and fossil wood from the Jogyeri Formation7,8,9. The Nampo leaf floras based on material from the Amisan and the Baegunsa formations were described, found similar to each other10 and assigned to the Dictyophyllum-Clathropteris type of palaeoflora10. This flora is known from the southern part of China from the Late Triassic to Early Jurassic11, with more occasional occurrences as far north as the Korean Peninsula, and characterized by plants typical of the tropical to subtropical climate10,12.
The specimen under study was collected from the Chungnam Basin of the Boryeong City, South Korea. The Boryeong deposits consist of dark shale, sandstone, siltstone, and conglomerate. The age of this insect fauna is still debatable, but it is generally considered to be Late Triassic or Early Jurassic4,5. We consider it as Late Triassic in age based on the analysis of the fossil conchostracans (Estherites kawasaki) and plants (Equisetites ferganensis).
It is the first record of Palaeontinidae from Korea, for the moment the oldest record of the family and one of the first reports of fossil insects from this locality – only Mesopsyche dobrokhotovae Novokshonov, 1997 (Mecoptera: Mesopsychidae) was listed recently13. There are a few other insect remains representing Hemiptera, Coleoptera, Ephemeroptera, Blattodea, Plecoptera, Grylloblattodea and Mecoptera found there.
Material and Method
This specimen was examined with a dissecting microscope (Nikon SMZ 800) and illustrated with the aid of a drawing tube attached to the microscope. Line drawing of tegmen was compiled using Adobe Photoshop CS graphic software. Fossil photograph was taken using a digital camera (Nikon D700).
The type material is deposited in the Department of Earth Science Education, Kongju National University, Korea. The wing venation nomenclature of Palaeontinidae used in this paper is based on the interpretations by Wang B. et al.14 and Nel et al.15.
Systematic Palaeontology
Order HEMIPTERA Linnæus, 1758
Suborder CICADOMORPHA Evans, 1946
Superfamily PALAEONTINOIDEA Handlirsch, 1906
Family PALAEONTINIDAE Handlirsch, 1906
Genus Hallakkungis Nam, Wang & Szwedo, gen. nov.
Type species: Hallakkungis amisanus sp. nov.; here designated.
Diagnosis: Tegmen with costal margin strongly curved at base. Stem of subcosta posterior (ScP) with several branches intersecting costal area and costal cell, basal portion of ScP shifted from common stem radius + media posterior + cubitus anterior (R + MP + CuA) in distance exceeding the length of basal cell. Stem MP bifurcated into branches MP1+2 and MP3+4 earlier than stem ScP + R forking. Stem of CuA straight; branch CuA2 strongly curved mediad in median 1/3 of its length. Crossvein r-mp1 apicad of crossvein mp3+4-cua; crossvein mp3+4-cua connected with stem CuA before CuA forking; crossvein mp3+4-cua forms part of nodal line; apex of clavus obtuse, due to strong curving of the utmost distal part of cubitus posterior (CuP) claval veins postcubitus (Pcu) and analis prima (A1) fused for a short distance as common stalk.
Remarks: Based on some venational characters, e.g. anterior margin indented, costa posterior (CP) present and ScP with several branches, this new genus is similar to Fletcheriana Evans, 1956, which was reported from the Middle Triassic of Australia. However, the genus Fletcheriana Evans, 1956, with Fletcheriana triassica Evans, 1956 (New South Wales, Australia) was transferred to Dunstaniidae16. In the same paper16 the species ‘Fletcheriana’ magna Riek, 1976, from the Triassic of South Africa17 was placed in the family Palaeontinidae. Later, the genus was discussed and some Jurassic species previously ascribed to this genus, were transferred to the other Palaeontinidae genera14. The genus Asiocossus Becker-Migdisova, 1962 from Kirghizstan18 is incompletely preserved (only basal portion of forewing), and the deposit was reevaluated as early Jurassic.
The new genus described above clearly differs from ‘Fletcheriana’ magna by the very strong curving of costal margin at base, distinct shift of basal part of ScP from common stem R + MP + CuA for a distance exceeding the length of basal cell (this portion is not clear in ‘Fletcheriana’ magna); stem MP forked anteriad of stem ScP + R forking (similar pattern, but less anteriad in ‘Fletcheriana’ magna); straight stem CuA (strongly curved in ‘Fletcheriana’ magna); distinct mediad curving of median portion of CuA2 branch (terminal CuA2 almost straight in ‘Fletcheriana’ magna); veinlet mp3+4-cua composed to nodal flexion line (only part close to CuA2 of mp4-cua composed to nodal flexion line in ‘Fletcheriana’ magna; this veinlet meets terminal MP4 in ‘Fletcheriana’ magna not the branch MP3+4); mp3+4-cua fused to stem CuA basad of CuA forking (connected with CuA1 slightly apicad of forking in ‘Fletcheriana’ magna); discal cell about three times as long as wide and narrow (discal cell about twice as wide as long in ‘Fletcheriana’ magna).
Etymology: The generic name is derived from “Hallakkungi” – the Flower Warden God in the Soch’on Flower Garden, from the Korean mythology. Gender: masculine, 3rd declension.
Hallakkungis amisanus Nam, Wang & Szwedo, sp. nov.
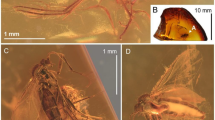
(Fig. 2a,b).

(a) photograph of holotype, No. KNU-2009018, part; (b) line drawing of forewing, No. KNU-2009018, drawn on photograph with Adobe Photoshop CS and adjusted with CorelDraw X7 software packages.
Diagnosis: Forewing elongately triangular, costal margin blade-like, distinct ambient vein and narrow appendix present; corrugations exceeding to narrow appendix and apical portions of apical cells. Stem CP faint, costal area widest at base, with intersecting branchings of ScP more distinct. Stem ScP emitting six braches intersecting costal area and costal cell, these branches dispersed in increasing distance each other. Veinlet mp3-4-cua long, slightly sigmoid, connecting branch MP3+4 just after its separation from stem MP to stem CuA at ½ of stem CuA length. Discal cell elongately almond-shaped, with acute apical angle, about 3 times as long as broad at widest point.
Etymology: The specific epithet is derived from the Amisan Formation, in which the fossil has been found.
Holotype: Single right forewing (tegmen), No. KNU-2009018. Deposited in Department of Earth Science Education, Kongju National University, Gongju, Korea.
Type locality, formation and age: Boryeong City, South Korea (N36°21′, E126°40′); Amisan Formation, Late Triassic.
Description: Right tegmen, 46 mm long, 21 mm wide. Anterior margin strongly curved at base, with distinct nodal incision basad of half of anterior margin length. Anteroapical angle acute, posteroapical angle of 121°; posteroapical margin straight; postclaval margin straight, curved towards apical angle; posteroclaval margin straight. Apex of clavus not reaching 1/3 of total length of forewing. Posteroapical margin corrugated, corrugations on appendix and at basal portions of apical cells. Costal margin (costa anterior; CA) strongly curved at base, blade-like, arcuate to nodal incision, arcuate apicad of nodal incision towards the anteroapical angle. Vein CP obscure, slightly curved and ending at the level of nodal indentation. Stem ScP distinctly separated from common stem R + MP + CuA at base, fused with stem R distinctly apicad of basal cell apex. Costal area and costal cell intersected by six branchings of ScP, with spaces between branchings sequentially increasing; apical portions of these branchings more distinct on costal area. Stem R + MP + CuA thick, stems R, MP and CuA leaving basal cell separately. Stem R strongly curved anteriad at base, forked basad of nodal line incision, slightly posteriad of stem MP forking; branch RA forked basad of nodal line incision, terminal ScP short, branch RA1 reaching anterior forewing margin at about half of post-incision portion length, branch RA2 longer, reaching anterior margin distinctly basad of anteroapical angle. Stem MP curved at base, forked basad of stem R forking, apicad of stem CuA forking; branch MP1+2 shorter than branch MP3+4; forking of branch MP1+2 merely apicad of nodal incision level, slightly earlier than forking of MP3+4; forking MP3+4 more apicad than nodal incision; terminals MP1, MP2, MP3 and MP4 slightly curved, reaching margin in median portion of posteroapical margin of forewing. Stem CuA leaving basal cell thick, straight, forked at basal ¼ of forewing length, basad of claval apex; branch CuA1 curved anteriad, reaching the posteroapical margin before posteroapical angle; branch CuA2 straight at basal 1/3, then strongly curved mediad, apical 1/3 thinner, slightly wavy, reaching posterior margin beside the posteroapical angle. Claval vein CuP thicker at base, thinner in apical portion, distinctly curved posteriad at claval apex, forming obtuse claval apex. Claval veins Pcu and A1 fused in apical ¼ of clavus. Posteroclaval margin distinct, not strongly separated from postclaval margin. Crossvein r-mp1 short, distinctly apical of nodal line; crossveins mp3+4-cua long, sigmoid, included to nodal (flexion) line, connecting branch MP3+4 slightly after stem MP forking with stem CuA at half of its length after separation from basal cell. Nodal line distinct, from nodal line incision at anterior margin, through terminal ScP, stem R forking, cutting branches MP1+2 and MP3+4 slightly after the stem MP forking, then, followed with crossvein mp3+4-cua for a long interval and after separating from mp3+4-cua crossvein, fused with basal 1/3 of branch CuA2 finally, separating from CuA2 and reaching CuP at level of claval apex.
Discussion
Hallakkungis amisanus gen. et sp. nov. is the first palaeontinid described from the Korea. Regarding its strongly triangular shape of the forewing, the new taxon resembles more the Early Cretaceous representatives of the family19. The more triangular shape of the forewing is an important character of the Palaeontinidae from the Late Jurassic to the Early Cretaceous20,21,22, but the venation pattern is clearly different. In the Jurassic and Cretaceous Palaeontinidae with triangular wings the hind wing is usually diminished due to particular kind of flight, discussed in ref. 23, but the question of a similar tendency in Hallakungis gen. nov. hind wings and flight performance remains open. The new genus Hallakkungis gen. nov. presents number of unique features, some of them shared also with presumed ancestors of Palaeontinidae, i.e. representatives of the family Dunstaniidae14. The strong curve of the costal margin at the base is one of peculiar features of the newly described genus. Such trend in forewing shape is observed in Dunstaniidae (e.g. Fletcheriana triassica), also in not related to Palaeontinoidea representatives of superfamily Pereborioidea – families Perboriidae and Ignotalidae24,25. The very late separation of RA and RP only just before the node, is similar to the situation in the Permian Prosbolidae. Such a late separation is common in Triassic palaeontinoids, and it is probably a plesiomorphic condition, but in Hallakungis gen. nov., it is unusually late. Another feature worth of mention is the basal separation of the ScP – such situation is present in majority of Jurassic Palaeontinidae, much less common in Dunstaniidae. The feature resembling the situation common with Dunstaniidae is the connection of crossvein mp3+4-cua to the stem CuA. In the vast majority of Palaeontinidae it is connected near, at the point or the distad of point of forking of stem CuA, while in Hallakkungis gen. nov. this point is distinctly more basal, at half of the common stem CuA length. In the other Palaeontinidae, this crossvein meets the terminal MP4, while in Hallakungis gen. nov., it is placed more basad, and meeting the branch MP3+4 in proximity of stem M forking, which is unusual. The exceptional feature of the newly described genus Hallakkungis is that the vast portion of crossvein mp3+4-cua is involved in forming the nodal line – while nodal line is more proximal then crossvein between in any other palaeontinoids. So, the evolutionary tendency of shifting to more apical position of this connection point is observed among the representatives of Palaeontinidae. It is interesting that the claval veins Pcu and A1 are fused in apical ¼ of the clavus length. This is a very ancient feature of early Cicadomorpha, present among the Permian representatives of the suborder, but also in the Triassic Dunstaniidae16. It seems that line of claval margin is rather continued on postclaval margin (the specimen is damaged at this area suggesting the incision); the incisions suggested at this point in the Dunstaniidae17, seems to be an artifact of preservation. However, the obtuse angle of the clavus is a unique feature of the Hallakkungis gen. nov. Another remarkable feature of this fossil is the strong curving in median 1/3 of the branch CuA2, such a character is not known among the other Palaeontinidae. It is interesting, that the corrugation at margin of the forewing is found also in some Early Cretaceous Palaeontinidae21, but such a feature is present also in taxa not related to the Palaeontinioidea representatives of the Cicadoidea: Cicadidae and Cercopoidea: Cercopidae26,27.
The newly described above genus and species presents a mixture of plesiomorphic, ancestral features shared with the Triassic Dunstaniidae, apomorphic, derived features found in the Jurassic and Cretaceous palaeontinids, and strongly autapomorphic features. General trends could be observed: the narrowing of the costal lobe, the progressively proximal separation of RA and RP, the development of a long basal crossvein between MP and CuA, so that the basal part of the wing comes to be supported by a three pronged fork of ScP + RA, RP and MP, rather than ScP + RA + RP, MP and CuA. The abovementioned features places this specimen among the unique taxa. Hallakkungis gen. nov., is an important link to understanding the evolutionary trends, tendencies and traits of early Palaeontinidae.
This new fossil genus of Palaeontinidae comes from the deposits of the Late Triassic Daedong flora, which is a typical representative of the Dictyophyllum-Clathropteris flora of Asia. It contains a number of plant taxa typical of the Triassic or to be relicts of the Permian floras10. The great reconstruction of the face of the Earth and of the organic world (the appearance of a great number of new plant forms) started in the Middle Triassic, and has been completed to the end of the Triassic. This distributional pattern of plants remained relatively stable during the rest of Mesophytic. Hence, this could be the source of evolutionary success of phytophagous, phloem-feeding Palaeontinidae.
The new discovery presented above is also the first record of the family Palaeontinidae from the Upper Triassic of Korea. We anticipate discovery of more well-preserved specimens from South Korea that will allow us to carry out more morphological and taxonomic studies, as well as palaeoecological, palaeobiogeograpical and evolutionary analyses of the Palaeontinidae and its relatives.
Additional Information
How to cite this article: Nam, K. S. et al. An extraordinary palaeontinid from the Triassic of Korea and its significance. Sci. Rep. 7, 40691; doi: 10.1038/srep40691 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Reedman, A. & Um, S. H. Geology of Korea. (Korea Institute of Energy and Resources, 1975).
Chang, K. H. Aspect of geologic history of Korea. J. Geol. Soc. Korea 31, 72–90 (1995).
Kim, J. H. Mesozoic tectonics in Korea. J. Southeast. Asian Earth. Sci, 13, 251–265 (1996).
Egawa, K. & Lee, Y. I. Jurassic sunorogenic basin filling in western Korea: sedimentary response to inception of the western Circum-Pacific orogeny. Basin Res. 21, 407–431 (2009).
Egawa, K. & Lee, Y. I. Thermal maturity assessment of the Upper Triassic to Lower Jurassic Nampo Group, mid-west Korea: Reconstruction of thermal history. Island Arc 17, 109–128 (2008).
Suh, H. G. et al. Four sheets of geological maps of the Seongju area (1:10,000) and the explanatory text. Korea Inst. Energy Resour . 1–42 (in Korean with English abstract) (1980).
Kim, J. H. New fossil plants from the Nampo Group (Lower Mesozoic). Korea Geosci. J. 5, 173–180 (2001).
Kim, J. H., Kim, H. S., Lee, B. J., Kim, J. M. & Lee, H. K. A new species of Leptostrobus from Upper Triassic Amisan Formation of the Nampo Group in Korea. Jour. Korean Earth Sci. Soc. 23, 30–37 (2002).
Kim, K. H. et al. Coniferous fossil woods from the Jogyeri Formation Upper Triassic of the Nampo Group, Korea. IAWA Journal 26, 253–265 (2005).
Dobruskina, I. A. Triassic Floras of Eurasia. Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommissionen. 10 (Springer Verlag, Wien New York, 1994).
Li, X. X. & Zhou, Z. Y. New advances of Mesozoic non-marine bio-stratigraphic researches in China. Adv. stud. Biostrat. China, 1979, 100–110 (in Chinese) (Nanjing Institute of Geology and Palaeontology, Academia Sinica, 1979).
Kimura, T. & Kim, B. K. Geological age of the Daedong flora in the Korean Peninsula and its phytogeographical significance in Asia. Proc. Jpn. Acad. 60, 337–340 (1984).
Nam, K. S. & Kim, J. H. Occurrence of the fossil Mesopsyche dobrokhotovae in the Late Triassic Amisan Formation, Nampo Group, Korea and its geological implication. J. Korean Earth Sci. Soc. 35, 161−167 (2014).
Wang, B., Zhang, H. C. & Szwedo, J. Jurassic Palaeontinidae from China and the higher systematics of Palaeontinoidea (Insecta: Hemiptera: Cicadomorpha). Palaeontology 52, 53–64 (2009).
Nel, A. et al. Traits and evolution of wing venation pattern in paraneopteran insects. J. Morphol. 273, 480−506 (2012).
Shcherbakov, D. E. Sistema i filogeniya permskikh Cicadomorpha (Cimicida, Cicadina), Paleontol. Zh. 2, 89–101. Translated into English as: Shcherbakov, D.E. Systematics and phylogeny of the Permian Cicadomorpha (Cimicida, Cicadina). Paleontol. J. 2, 87–97 (1984).
Riek, E. F. A new collection of insects from the Upper Triassic of South Africa. Ann. Natal. Mus. 22, 791–820 (1976).
Becker-Migdisova, E. E. Nekotorye novye poluzhestokrylye i senoyedy [Some new hemipterans and psocodeans]. Paleont. Zh. 1, 89–104 (1962).
Menon, F., Heads, S. W. & Martill, D. M. New Palaeontinidae (Insecta: Cicadomorpha) from the Lower Cretaceous Crato Formation of Brazil. Cret. Res. 26, 837–844 (2005).
Ueda, K. A new Palaeontinid species from the Lower Cretaceous of Brazil (Homoptera: Palaeontinidae). Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. 16, 99–104 (1997).
Whalley, P. E. S. & Jarzembowski, E. A. Fossil insects from the lithographic limestone of Montsech (late Jurassic-early Cretaceous), Lerida Province, Spain. Bull. Br. Mus. Nat. Hist. (Geol.) 38, 381–412 (1985).
Ren, D., Yin, J. C. & Dou, W. X. Late Jurassic palaeontinids (Homoptera: Auchenorrhyncha) from Hebei and Liaoning Provinces in China. Entomol. Sinic. 5, 222–232 (1998).
Wootton, R. J. Reconstructing insect flight performance from fossil evidence. Acta zool. cracov. 46 (suppl. Fossil Insects), 89–99 (2003).
Riek, E. F. Fossil insects from the Upper Permian of Natal, South Africa. Ann. Natal Mus. 21, 513–532 (1973).
Pinto, I. D. & de Ornellas, L. P. Permian insects from the Paraná Basin, South Brazil. III – Homoptera – 1 - Pereboridae. In: Sanguinetti, Y. T. (ed.) Anais do 2° Congr Latino-Amer Paleontol, 26 a 30 do Abril 1981, Porto Alegre 1, 207–213 (1981).
Distant, W. L. Homoptera, Fam. & Cicadidae, Subfam. Gaeaninae. In: Wytsman, P. (ed.) Genera Insectorum 158, 1–38, pls 1–3 (1914).
Lallemand, V. Homoptera, Fam. Cercopidae . In: Wytsman, P. (ed.) Genera Insectorum 143, 1–167 (1912).
Acknowledgements
This research is supported by the Beijing Talented Scholar Program Foundation (No. 2011D002022000006), National Natural Science Foundation of China (No. 31301907), Innovation Team Project of Beijing Academy of Science and Technology (IG201101N). This paper results from the scientific exchange programme between Chinese Academy of Sciences and Polish Academy of Sciences awarded to JS.
Author information
Authors and Affiliations
Contributions
Material was collected and preliminarily assigned by K.S.N. and J.H.K., geological data on the site and specimen were provided by K.S.N. and J.H.K., specimen was identified, examined and analyzed by Y.W. and J.S.; Y.W. and J.S. wrote the paper; K.S.N., Y.W. and J.S. prepared figures; D.R. revised the manuscript. The authors declare that they have no conflict of interest.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Nam, K., Wang, Y., Ren, D. et al. An extraordinary palaeontinid from the Triassic of Korea and its significance. Sci Rep 7, 40691 (2017). https://doi.org/10.1038/srep40691
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep40691
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
