
Abstract
Mountains of Anatolia are one of the main Mediterranean biodiversity hotspots and their richness in endemic species amounts for 30% of the flora. Two main factors may account for this high diversity: the complex orography and its role as refugia during past glaciations. We have investigated seven narrow endemics of Centaurea subsection Phalolepis from Anatolia by means of microsatellites and ecological niche modelling (ENM), in order to analyse genetic polymorphisms and getting insights into their speciation. Despite being narrow endemics, all the studied species show moderate to high SSR genetic diversity. Populations are genetically isolated, but exchange of genes probably occurred at glacial maxima (likely through the Anatolian mountain arches as suggested by the ENM). The lack of correlation between genetic clusters and (morpho) species is interpreted as a result of allopatric diversification on the basis of a shared gene pool. As suggested in a former study in Greece, post-glacial isolation in mountains would be the main driver of diversification in these plants; mountains of Anatolia would have acted as plant refugia, allowing the maintenance of high genetic diversity. Ancient gene flow between taxa that became sympatric during glaciations may also have contributed to the high levels of genetic diversity.
Similar content being viewed by others


Introduction
The Mediterranean Basin is considered one of the most important hotspots of the world1 because of its high concentration of endemic species. Up to 60% of these are specifically narrow endemics2. Southern Turkey stands out especially and shows a wealth of endemic species3. According to some authors4, percentage of endemism reaches 30% of the flora with a maximum concentration in the areas of transition where the floras of different regions meet. Anatolia constitutes a crossroads of three floristic domains: Pontic, Irano-Turanian and Mediterranean5. The Mediterranean floristic domain is confined to the western and southern coastal strips and adjacent mountain ranges, mainly Western and Central Taurus, with some minor enclaves on the coast of the Black Sea3. It is worth noting the definition of a diagonal line, known as the Anatolian Diagonal (Fig. 1), formed by a long mountain range with heights of 3000–4000 m, which has served as a dispersal corridor for a number of plants and animals from northeast Anatolia and the Black Sea region to south Anatolia3. Mountain ranges have acted as refugia for Mediterranean species during glaciations6, being the main specific refugia the Taurus, the whole W Anatolia and the Amanus.

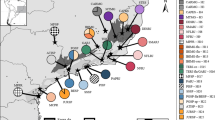
Location of the sampled populations for the seven species studied.
Symbols for the sampled populations: • = C. amaena; ○ = C. antalyensis; ■ = C. cadmea; □ = C. luschaniana; Δ = C. lycia; ▲ = C. lycaonica; ☆ = C. wagenitzii. This figure has been generated with ArcGIS v. 9.3 (ESRI, Redlands, CA, USA) and modified using Adobe Illustrator CS5.1 (Adobe Systems Incorporated, San Jose, CA, USA). Map layers were obtained from the site www.naturalearthdata.com.
In this context of high endemicity, one of the most diverse and most characteristic genera of the Mediterranean flora5 is Centaurea (Compositae-Cardueae) with over 250 species7. Centaurea constitutes an excellent model for analyzing the speciation and diversification processes in the region. The main centres of diversification of Centaurea coincide largely with the main refugia defined in the Mediterranean7,8,9, one of them being the Mediterranean part of the Anatolian Peninsula6. In fact, Centaurea is the third genus with more endemic species of Turkey10 and the number of species is constantly increasing since the publication of Davis Flora of Turkey11. Latest estimations suggest that there are 159 species in the area, 118 of them endemics12. Centaurea is also notable for the frequent introgression and hybridisation9 and references therein, which in some cases has offered a window into the history of the species by revealing gene flow between species not sympatric at present13.
Amidst the different sections of genus Centaurea that show specific diversification in the Mediterranean hotspots, we will focus on sect. Centaurea subsect. Phalolepis. All the nine Anatolian species of subsect. Phalolepis listed in the Flora of Turkey are narrow endemics11. Two of them, C. hieropolitana Boiss. and C. tossiensis Freyn & Sint. ex Freyn, fall outside Phalolepis according to the results of the latest molecular survey of Centaurea and should be included in a new section, sect. Hierapolitanae9. The species that remain in Phalolepis are C. amaena Boiss. & Balansa, C. antalyensis H. Duman & A. Duran, C. aphrodisea Boiss., C. cadmea Boiss., C. dursunbeyensis Uysal & Köse, C. luschaniana Heimerl ex Stapf, C. lycaonica Boiss. & Heldr., C. lycia Boiss., and C. wagenitzii Hub.-Mor.
On morphological grounds, Turkish species of Phalolepis share a very similar morphology, which was reviewed in depth recently14. In sharp contrast with Greece where most of species were at some point subordinated to other widespread taxa15, species of Phalolepis described from Turkey are fairly undisputed. They are usually mountain chasmophites on limestone, with the exception of C. wagenitzii, which grows on serpentines at sea level.
We chose the group of Phalolepis species as a subject for phylogeographic studies for three reasons: firstly, the group is morphologically cohesive and its limits have been recently reviewed and redefined9; secondly, there are no doubts on the taxonomy of the species, thanks to a recent morphological review14; and thirdly, subsection Phalolepis has been subject of a recent study on speciation in Greece15, and a similar study in Turkey would allow a comparison of results of high interest. The study in Greece concluded that it was a case of allopatric speciation by area fragmentation mediated by the special topography of the mountains of Greece, which favored isolation, and pointed out the character of refugium of the region during the Pleistocene glaciations15.
As in the case of Greece, we selected microsatellites, a highly variable type of marker, for analyzing subsect. Phalolepis in Turkey. Microsatellites have the advantages of codominant inheritance, reproducibility and high levels of variability, and are extensively used for analysing genetic structure. Previous studies using microsatellites have resolved the population structure of other species of Centaurea16,17,18,19,20 and have unraveled the speciation process in Phalolepis in the refuge of continental Greece15. Besides population analyses using microsatellites, ecological niche modelling (ENM) has also been employed to get insights into the paleodistribution of species. ENM represents an independent method of biogeographic inference on a given species, thus being an ideal complement to the classical genetic methods21.
Our specific objetives are the following: (1) to investigate population structure by estimating genetic diversity within and between narrow endemics using microsatellites, and compare the results with those in Greece for verifying whether the evolution of the endemicity in Phalolepis follow a similar pattern; (2) to analyse the correlation of morphological species and genetic clusters; (3) to explore the levels of ancient and recent gene flow and verify the extent of introgression; (4) to verify whether the range expansions/contractions of the studied Centaurea endemics is related to the special physiography of Anatolia using ecological niche modelling in Centaurea cadmea, the only species in this study that shows enough occurrences as to build a model.
Results
Genetic diversity
All surveyed loci were polymorphic for all the populations (except 21D9 for LY, LU, LYC1, AMA, and CA3). We detected a total of 97 alleles, ranging between four (locus 21D9) and 29 (locus 12B1); average was 13.9 alleles per locus. At population level, the mean number of alleles per locus (A) ranged between 4.286 in populations LYC1, LYC2, WA2, and CA2 and 7.286 in population LU (Table 1). We detected exclusive alleles for many populations, even though numbers were low (from one to five; Table 1) and frequencies were low too (mean = 0.065). Population LYC2 showed the lowest value of expected heterozygosity (He = 0.491), whereas population WA1 harboured the highest value (He = 0.669; Table 1). The least variable species was C. lycaonica (He = 0.498), whereas the highest genetic diversity was shown by C. antalyensis (He = 0.655; Table 1).
Only one loci pair showed evidence of linkage disequilibrium (28A7 vs. 17E3). Many individual loci (42 out of 77 possible cases) exhibited significant deviations from H–W equilibrium expectations (P < 0.05); accordingly, FIS values at population and species level were positive and significantly different from zero (Table 1), which can be attributed to inbreeding and/or to the occurrence of null alleles22. Given that the frequency of null alelles in our dataset is very low [range = 0.0003 (21D9 locus) − 0.178 (12B1 locus); average = 0.095], we do not expect significant biases in our genetic analyses (it has been suggested that biases are negligible when null alleles are present at frequencies below 0.20022). However, there are positive correlations between their frequency and FIS values within (N = 77, R2 = 0.760, P = 0.000) and across populations (N = 7, R2 = 0.573, P = 0.049), indicating that the significant deviations from H–W equilibrium can be partly attributed to the presence of null alleles.
All but one genetic divergence values between populations based on FST were significant (P < 0.05). These values ranged from 0.027 (LU-LYC1) to 0.297 (LYC2-AMA), with a mean of 0.198 (Table 2). The FST values between pairs of species were significant in all the cases, and somewhat smaller than the FST values between populations (Table 3): they ranged from 0.057 (C. lycaonica-C. luschaniana) to 0.217 (C. lycia-C. amaena), with a mean of 0.135. Corrected FST values for null alleles showed very similar to those found using the uncorrected dataset (the differences were only ca. 5–10%), both between populations and between species (Supplementary Tables S1 and S2); the effects of null alleles on the genetic structure of populations, thus, would be negligible.
According to Evanno’s approach, K = 10 was the most likely number of genetic clusters for Structure simulations, as this is the only clear peak. The ln Pr(X|K) also reached a plateau when K = 10 (Supplementary Fig. S1). Although several of the studied populations had their “own” clusters, other populations (LY, LU, LYC1, and WA1) showed a high degree of admixture (Fig. 2). Notably, not all the grouping schemes input in AMOVA were significant. The within-population component accounted for most of the total variation (84.42%), the among-population component contributed much less (15.53%), whereas the among-taxa component (i.e., when the populations were grouped into species) was negligible (0.05%) (Table 4). Even though the percentage of variance explained by the first two components was not very high (52.90%), the PCoA analysis at population level was in close agreement with both the Structure and (particularly) with AMOVA results: populations are relatively isolated among them, with no signs of taxonomically meaningful aggrupation (i.e. all populations belonging to a same species were invariably located in different quadrants; Fig. 3).

Membership proportion of 323 individuals to ten groups (K = 10) according to the Bayesian analysis of population structure carried out with Structure.
For abbreviation of populations, see Table 1.

Principal Coordinate Analysis performed from pairwise genetic distances between populations.
For abbreviation of populations, see Table 1.
The first two assigned barriers in the Barrier analysis (Supplementary Fig. S2), mainly separated the populations located on western Taurus Mountains (that is, those around the Gulf of Antalya). With the third barrier (B = 3), separations in other regions (Pontic Mountains, central Taurus Mountains) started to be evident, although some separations (LU vs. LYC1, LY vs. CA2, and WA1 vs. WA2) did not appear until B = 8. The BayesAss analysis confirmed the absence of recent gene flow between populations: all but one pairwise m values were at least one order of magnitude below 0.114 (this figure separates real migration from noise; Table 5). Regarding historical gene flow, as estimated with the software MIGRATE-N, Nm values were relatively low (from 0.263 to 1.193, averaging 0.466; Table 6). Total immigration rates were highest for 1LYC, 1WA, and 2WA (all with Nm > 5), whereas the populations that showed total emigration rates > 5 were LU, 2WA, and 2CA (Table 6).
Ecological niche modelling
According to the MaxEnt jackknife tests of variable importance, the precipitation variables were more informative for the model than the temperature ones (the more informative variables were, by far, precipitation of the driest month and precipitation seasonality; Supplementary Fig. S3). The present-day distributional predictions for C. cadmea were largely congruent with the known species occurrences, although other areas appear as suitable (shaded in Fig. 4), such as mountains of NW Anatolia and some scattered areas along the western and central Taurus Mountains (Fig. 4A). Projections of the species niche to the LGM climate produced considerably different maps of presence/absence. With the CCSM model almost all the Anatolian Peninsula appears as suitable, leaving only as unsuitable the eastern and southern coastal areas (Fig. 4B). The MIROC model showed that most of the mountainous areas of the Anatolian Peninsula were suitable for C. cadmea at the LGM with the exception of the plateau region of central Anatolia (Fig. 4C). Both LGM projections show a considerable increase of the potential area of C. cadmea compared to the present time, even for the likely more “realistic” MIROC model (with an increase of about four-fold of the suitable area, compared to an increase of ten-fold for the CCSM). The LGM projections should be, however, treated with extreme caution because the uncertainty of projecting the present-day ENM to the LGM should be added to that resulting from the use of a very small number of occurrences.

Potential distribution (shaded areas) in Turkey of Centaurea cadmea obtained with MaxEnt.
(a) At the present time; (b) at the Last Glacial maximum (LGM, ca. 21,000 years BP) using the Community Climate System Model (CCSM80); (c) at the LGM using the Model for Interdisciplinary Research on Climate (MIROC81). Black dots indicate current populations of the species. The reconstructed LGM coastlines are represented in (b) and (c), with current coastlines superimposed as dotted lines. This figure has been generated with ArcGIS v. 9.3 (ESRI, Redlands, CA, USA).
Discussion
Genetic diversity
Genetic theory predicts low levels of genetic diversity for narrow endemic species because of a combination of traits usually associated with rarity: low population sizes, isolation of populations, inbreeding, and ecological specialization23,24. In support of this view, several meta-analyses have demonstrated that there is an association between rarity and low genetic variability25,26,27; such association is even more compelling for those species that are limited to one or very few localities, with dozens or, at most, a few hundreds of individuals (often referred as “extremely narrow endemics”, ENEs28). In the Mediterranean, two biological traits are often present in the endemic flora: production of few and small flowers, and low investment in pollen and seed production29. Both traits might also have a role in the association between rarity and low genetic diversity. Accordingly, examples of Mediterranean narrow endemics showing very low levels of genetic diversity are abundant, e.g. Anchusa crispa30, Aquilegia barbaricina/A. nugorensis/A. nuragica31, Coristospermum huteri28, or Zelkova sicula32.
Contrarily to the expected patterns for narrow endemics, Anatolian species of subsect. Phalolepis are not genetically depauperated at all (mean A = 5.643; mean He = 0.580; Table 1). Even the most restricted taxa such as C. antalyensis, C. cadmea subsp. cadmea, and C. lycaonica (all are classified as CR, see below in Material and Methods) have moderate to high levels of genetic diversity, and C. antalyensis, only known from the type locality and having less than 250 individuals, harbors the highest levels of variability in terms of expected heterozygosity (He = 0.655; Table 1) among all studied taxa. Genetic diversity values in Turkish species of subsect. Phalolepis are among the highest within the genus (mean A = 5.160; range 3.100–7.286; mean He = 0.504; range 0.073–0.779; Supplementary Table S3), and are comparable to the values obtained for a series of narrow endemics of the same subsection from the mountains of Greece using the same set of microsatellites (A = 5.363; He = 0.587)15. Comparison with the “reference” values for plants (He = 0.420 for endemic species, He = 0.620 for widespread ones) provided in the meta-analysis of Nybom27 is another proof that our study species conserve considerable microsatellite genetic variability in spite of their rarity.
Our results, as well as the growing number of studies reporting narrow endemic species with unexpectedly high levels of genetic polymorphism33,34 suggest that the geographic range is not always a good predictor of genetic diversity in plant species. It is generally agreed that a complex network of factors shape genetic diversity in plant populations, which are usually classified into two categories35: (1) intrinsic biological properties of the species (i.e., life-history traits and ecological interactions) and (2) extrinsic dynamic processes which affect species (i.e., historical factors that may include occurrence of bottlenecks, divergence events, or Quaternary expansions/retreats). Species of Centaurea share most of their biological traits: they are usually protandrous and usually self-incompatible, and they have entomophilous pollination by a wide range of insects36,37,38. Thereafter, intrinsic factors should be discarded and differences in polymorphism between the studied Centaurea species (Table S3) are more likely extrinsic and attributable to their evolutionary history.
The endurance of glacial-interglacial cycles in mountain glacial refugia in the Pindus Mountains of Greece was the main reason provided for the high levels of polymorphism detected in the Greek taxa of subsect. Phalolepis15. Likewise, the relative environmental stability of another mountainous refugium in the Pre-Pyrenean Mountains has been suggested as an explanatory factor for the moderate microsatellite diversity in C. emigrantis and C. tripontina39. The Pindus and the Pre-Pyrenean regions range among the “phylogeographic” glacial refugia recognized within the Mediterranean Basin6. The southern Anatolian Peninsula and especially the western and central sections of Taurus Mountains is home of several refugia6, and is also characterized by a high concentration of plant diversity and endemism4.
Several factors have contributed to the refugial character of the Taurus: the rugged topography of these mountains with peaks surpassing 3000 m, its closeness to the sea, which would have constituted a continued source of moisture even at the most arid phases of the Pleistocene; and, in contrast to the Pyrenees or the Alps, the lack of major ice-sheets. Only the peaks over 1900 m were glaciated40. This scenario is consistent with the most recent paleoecological reconstructions for the Taurus Mountains based on pollen sites or climatic simulations: the mountains would have been covered by temperate/boreal forests and parklands instead of more arid formations such as desert or steppe41,42,43.
The results in the montane species of Centaurea studied by our team15,39 clearly support this scenario. Centaurea species, like other mountain endemics, would have found abundant favorable pockets throughout the unglaciated, relatively moist Mediterranean mountains, and survived the repeated glacial/interglacial cycles by altitudinal migrations without large geographical displacements. These movements would have favored secondary contacts at the glacials and populations would have admixed, largely blurring the genetic differentiation produced by periods of isolation at the interglacials. This process was favored by the much longer duration of glacials compared to interglacials and has also been proposed for other plants from the Mediterranean mountains13,34,44. Genetic connectivity is also present in Anatolian species of subsect. Phalolepis (FST = 0.198) and the results of ENM constitute another indirect evidence because the suitable habitat projected at the LGM is significantly increased for C. cadmea compared to the present time (Fig. 4). The increase of LGM suitable areas may be common to the other studied Turkish species of subsect. Phalolepis, given that almost all of them have the same ecological requirements, and contacts between populations would have been frequent.
Systematic and evolutionary implications
The results of the genetic study only partially coincide with the morphological classification usually accepted. The Structure analysis defines individual clusters for C. amaena and C. antalyensis, and partly for C. wagenitzii (Fig. 2). By contrast, several other species share a common gene pool (cases of C. lycaonica and C. luschaniana, along with a population of C. lycia). Besides, C. cadmea cannot be defined as a species because each population forms a different gene cluster (Fig. 2), which is very evident in the PCoA (each population falls into a different quadrant; Fig. 3). It is likely that C. cadmea is in active process of speciation and in fact there is a morphological reason to support this hypothesis. The northern populations have been segregated as a different subspecies, C. cadmea subsp. pontica Wagenitz ex Y.B. Köse & Ocak. This case is similar to Greek C. chrysocephala Phitos & Georgiadis, a species that could be in active speciation process in its most isolated population in Meteora Mountains15.
The results of the AMOVA analysis also indicate that the species cannot be defined as independent entities; the among-taxa component is notoriously low (0.05%; Table 4). This difficulty often arises when dealing with taxa that are scarcely divergent genetically45, as in the present case (Table 3). The use of the ITS region and plastid markers in Centaurea has usually resulted in a general lack of resolution: the resulting phylogenies consist usually of large polytomies9. Introgression and incomplete lineage sorting following recent speciation are usually invoked as the major causes of species-level polyphyly46. Speciation of Centaurea subsect. Phalolepis in Anatolia is recent because an age of 1.55 (0.32–3.13) Ma was assigned to the Turkey/Cyprus haplotype9. We will discuss in terms of current and former gene flow the reasons that make us think that cases of shared gene pool are due to introgression in a recently differentiated group.
Recent gene flow is now almost absent and populations are genetically isolated, as deduced from the results of BayesAss (Table 5). The only case of current flow is the connection between one of the populations of C. lycia (LYC1) and C. luschaniana. In this case, Barrier results also indicate that there are no barriers between them, even with B = 8 (Supplementary Fig. S2). This may be due to lack of topographical barriers that do exist, however, with the other population of C. lycia (LYC2; Fig. 1), which grows on the other side of the mountains. The population LYC2 appears to be isolated and free from admixture in Structure (pale blue, Fig. 2) and the Barrier analysis effectively separates it from the rest of populations from B = 5 (Supplementary Fig. S2). Other instances of high admixture (C. lycaonica and population 1 of C. wagenitzii) are evident in Structure (Fig. 2), for which recent gene flow is not a valid explanation as reflected in BayesAss. For these two populations (especially for WA1) we can argue, instead, ancient gene flow as shown by the MIGRATE-N results (Table 6). Although the Nm values suggested by this software are not high at all, at least there is some signal of exchange of genes; however, we cannot assign an exact temporal framework for these old contacts, as they may range from the Holocene (~450 ya) to the end of LGM (~18,500 ya) following our previous approximation15. According to our reconstruction of the ecological niche at the LGM for one of the species studied here (Fig. 4; see below), gene flow would have been much more extensive during the glaciations than at present.
Lower but still noticeable levels of admixture are evident in almost all species studied (Fig. 2). For these cases, we can also argue ancient gene flow. In none of the cases of admixture, either recent or old, there is morphological evidence of introgression. That is, no intermediate characters have been observed between the species that have been connected. Ancient contacts between non sympatric species in the Mediterranean region have been explained in several cases by altitudinal migrations forced by the glaciations13,47. In our case, the contacts may have occurred by the pathway of the mountains surrounding the Anatolian plateau (Fig. 1). According to the ENM for C. cadmea, its current area is much smaller than the potential distribution of the species in the LGM (Fig. 4C). Centaurea cadmea migration would be possible through the zones defined by the ENM, which coincide with the archs of mountains just described, especially via the Anatolian Diagonal in the east of the plateau. At the LGM, these mountainous archs were probably covered by boreal/temperate forests and parkland instead of xeric grasslands or steppe, as revealed by climate simulations41,42. Fossil pollen records also points towards cold but relatively moist conditions (probably due to the orographic rain) for most of the mountainous southern regions of Anatolia43. A pollen record from Lake Iznik, in the mountains of NW Anatolia, is suggestive of steppe but with a still important fraction of arboreal vegetation (ca. 25%)48. There are no pollen sites for the Pontic Mountains, but there are currently some relict termophilous species in the region, such as Pterocarya fraxinifolia. Along the Black Sea coastline, Carya and Glyptostrobus persisted until the Holocene, and were probably extinct due to human disturbance49. Presence of termophilous species indicates that glacial refugia would have existed in the region. Contrarily, the Anatolian Plateau was dominated by Artemisia-steppe; at the only pollen site from the plateau, only 10% of the pollen is arboreal43. Presence of steppe suggests much drier conditions to those of the surrounding mountain archs. All the species of the study except C. wagenitzii share similar ecological requirements (see Introduction), and it is plausible that the same pathways favorable to C. cadmea have been exploited by other species of the study. The lack of barriers between the most isolated species, C. amaena, and the core of the SW species in the Taurus (Supplementary Fig. S2) reinforces the hypothesis of the existence of migration routes through the mountains of S Turkey. In addition to the ENM, migration routes though the northern mountain arches are also suggested by the lack of barriers between C. amaena and population 3 of C. cadmea (Supplementary Fig. S2) as well as by the relatively high Nm values between populations 2 and 3 of C. cadmea (located in the Pontic Mountains) and some of the studied populations of SW Taurus.
Greece and Turkey models
A comparison between the diversification patterns in Greece and Anatolia is straightforward. The similarities are relevant, with a main factor having shaped genetic diversity and diversification in Centaurea subsect. Phalolepis: the rugged and dissected topography. The Anatolian mountains favored the population isolation of a more widespread ancestor of the species of the group, triggering allopatric speciation. At the same time, the mountains would have constituted refugia during the glaciations, protecting the populations from genetic bottlenecks and promoting contacts between them as shown by the high genetic diversity found. Finally, the mountain arch that encircles the Anatolian Plateau, especially the Anatolian Diagonal at the east, offered a pathway during glacials for the migration of species.
The main difference between Greece and Turkey in the diversification of Centaurea subsect. Phalolepis is the time frame: speciation in Turkey is probably more recent. The main reasons for reaching this conclusion are: (a) Genetic clustering in Greek species using STRUCTURE show only limited level of admixture15, while in Turkey the levels of admixture are much higher (Fig. 2). (b) FST values between species are larger in Greece compared to Turkey (0.182 vs. 0.135). These results support that Greek species would have almost achieved complete isolation, while Anatolian ones would be still in progress. In sum, processes involving diversification in both hotspots are basically the same, with a slightly later temporal displacement in Turkey when compared to the Greek case.
Evolutionary and conservation implications
Mountains elsewhere are often regarded as suitable Quaternary refugia, where both persistence and differentiation of plant lineages would have occurred6; thus, it is not surprising that mountains often harbor high taxonomic richness as well significant rates of endemism50. For the mountains of the Mediterranean basin, it was suggested a dual model to explain the patterns of endemism51: prevalence of paleo-endemics in its western section and dominance of neoendemics in its eastern half; according to these authors, the dominance of differentiation processes over conservative ones in the Eastern Mediterranean basin would have resulted from the relative geologic youth of this sector and, remarkably, the moderate role of Pleistocene glaciations49. The present study as well as our previous one15 in the subsect. Phalolepis of Centaurea (a very young group) are paradigmatic of the role of Mediterranean mountains as places where active speciation processes have taken, and are still taking place. The high levels of genetic diversity and the moderate genetic isolation between species support the role of mountains as glacial refugia and “species pumps”.
In conservation biogeography, one of the most debated questions at present is whether priority should be given to areas of active speciation (“species cradles”) or, on the contrary, to areas that are a sink of ancestral taxa (“species museums”)52. Although with the conservation of “species museums” we are avoiding the loss of relict taxa (that, in most cases, have unique, irreplaceable evolutionary history), conserving species pumps we are preserving those places that contribute most to the growth of the Tree of Life (i.e., we are guaranteeing the maximum levels of both present and future biodiversity). Conserving the eastern Mediterranean mountains in a more or less pristine state is, however, a challenging issue; overgrazing and erosion are rampant, and tourism impacts are quickly increasing53. Regrettably only 1.2% of Turkey’s terrestrial area is strictly protected (much below the international standards), and high mountains, shrublands, and steppe ecosystems are underrepresented53. Therefore, although conserving large landscapes is perhaps the best tool in plant by preserving processes that create and maintain biodiversity54, it seems more realistic to ensure the conservation at population level of those species of conservation interest, such as narrow endemics. Unfortunately, most of the studied species do not enjoy at present of suitable conservation measures (even despite that some of them are classified as CR); for instance, less than 20% of all the populations of the studied species are located within a nature reserve and, to our knowledge, no specific conservation measures, either in-situ or ex-situ, exist.
Methods
Plant material
Most species of subsect. Phalolepis clasically recognized11 are included in the study, with the addition of a couple of taxa described afterwards. We excluded C. hieropolitana and C. tossiensis (see Introduction). Tetraploid C. aphrodisea and C. dursunbeyensis were also excluded due to the difficulties in interpreting microsatellite results in polyploids55. Thereafter, our survey includes seven species: C. amaena (one population), C. antalyensis (one population), C. cadmea (three populations), C. luschaniana (one population), C. lycaonica (one population), C. lycia (two populations) and C. wagenitzii (two populations).
Centaurea amaena grows in rocky places in the Kayseri province and it is restricted to a small area between Yılanlı and Erciyes Mountain. The only two localities together cover an area of around 0.55 km2, and the number of individuals was calculated to be 5672. Its threat category was suggested to be EN (Endangered)56. However, the species displays fairly local distribution in an isolated area open to urbanisation as well as limited number of mature individuals. The species should be categorised as CR (Critically Endangered) according to 2001 IUCN criteria57 [CR B1ab(i,iii,iv) + 2ab(i,iii,iv)].
Centaurea antalyensis is known only from the type locality and the number of mature individuals is below 250. The species was listed as CR (Critically Endangered)58. According to our observations, the species is very local and its distribution area is less than 10 km2, which agrees with the assigned category.
Centaurea cadmea is represented by two subspecies in Turkey. Both of them have fairly local distribution limited to one or two small populations. Centaurea cadmea subsp. cadmea is known from a few localities in Denizli, growing on calcareous rocky slopes. The number of individuals is less than 250 and this subspecies is under high risk due to overgrazing. It was evaluated as LR (Lower Risk)56. However, it should be categorized as Critically Endangered (CR) because of its local distribution [B1ab(i,iv,v)]. Centaurea cadmea subsp. pontica is present in the Zonguldak and Bartın provinces, where it is common. It grows on rocky slopes overlooking deep valleys, at 275–700 m. Its threat category should be VU following 2001 IUCN criteria57 [B2ab(i,iii,iv)].
Centaurea luschaniana is a local endemic species that is restricted to a limited area between Elmalı and Korkuteli. This species is known from 4 or 5 populations on the line Korkuteli-Elmalı. Its threat category was evaluated before as LR (Lower Risk)56. According to our observations, the populations of this species are in good health. However, some of them may become under intense pressure due to farming activities of nearby villages. Therefore, we suggest that the species should be categorised as VU (Vulnerable) following 2001 IUCN criteria57 [VU B1ab(ii,iv)].
Centaurea lycaonica is a local endemic species in Konya province of Central Anatolia. It was classified as Critically Endangered (CR)56. There are four–five very close populations that are threatened due to extreme livestock grazing and gold mining59. Further alteration of the habitat of C. lycaonica would result in the extinction of the species. Thereafter, the species is correctly categorized as CR (Critically Endangered) based on 2001 IUCN criteria57 [B1ab(i,ii,iii,v) + 2ab(ii,iv,v)].
Centaurea lycia is a regional endemic species growing on rocky cliffs in Antalya province. There are six populations, which appear in good condition in terms of mature individuals (K. Ertuğrul, pers. obs.). Its threat category was evaluated as LR (Lower Risk)56. However, the species is exposed to heavy tourism activities as well as suffering the negative impacts of urbanisation (K. Ertuğrul pers. obs.). Therefore, we suggest that the species should be categorised as Vulnerable (VU) following 2001 IUCN criteria57 [VU B1ab(ii,iii,iv)].
Centaurea wagenitzii is known only from the type locality, with an estimated area of occupancy of less than 5 km2. Its threat category was evaluated before as EN (Endangered)56 under the criterion B2a. According to our observations, the population comprises less than 250 individuals, and it should be classified as Critically Endangered (CR) based on the 2001 IUCN criteria57 [B2ab(i,ii,iv,v)].
Voucher information is provided in Supplementary Table S4, and a map with the location of the collected populations is shown in Fig. 1.
DNA isolation and microsatellite loci
We extracted genomic DNA from dried leaves using the CTAB method60. For verifying cross-amplification of the seven Turkey-endemic species, we carried out a preliminary test using 16 SSR markers developed for other species of Centaurea (C. corymbosa61, C. diffusa and C. stoebe62). We amplified polymorphic bands from seven microsatellites (CD37, 42CM27, 12B1, 13D10, 17E3, 21D9, 28A7) in all the studied species, as explained in López-Vinyallonga et al.15. We used at least 30 individuals per population when possible, accounting for a total of 323 individuals from 11 populations. Genotyping was performed as described in López-Vinyallonga et al.15.
Genetic analysis
Software GenAlEx v. 6.1 63 and Genetix v. 4.05 64 were used for estimating genetic diversity parameters at both species and population levels: (a) mean number of alleles per locus (A); (b) number of private alleles (PA); (c) percentage of polymorphic loci (P95); (d) observed heterozygosity (Ho); (e) unbiased expected heterozygosity (He); and (f) inbreeding coefficient (FIS) by the method of Weir & Cockerham65. GenePop v. 4.0.1066 was used for calculating possible deviations from Hardy-Weinberg (H–W) equilibrium and for checking genotypic linkage disequilibrium between pairs of loci at population level and across all populations; for both calculations the software uses a Fisher’s exact test following the Markov chain (MC) algorithm67. Frequency of null alleles was calculated using FreeNA68, which was also used to estimate FST values between pairs of populations and species (with and without the ENA correction for null alleles68).
The spatial genetic structure was assessed through four different methods. First, Structure v. 2.3.469, a widely-employed clustering software that is a based on a Bayesian algorithm, was used. On the basis of preliminary runs, K was run from 1 to 12 (20 iterations per K) assuming an admixture model with correlated allele frequencies, and with a priori grouping of individuals into populations (but not into species). The length of burn-in period and the MCMC replications were set to 105 and 106, respectively. The most likely value of K was determined both by choosing the smallest K after the log probability of data [ln Pr(X|K)] values reached a plateau69 and by the ΔK statistic of Evanno et al.70 with the aid of Structure Harvester71. Second, a molecular variance analysis (AMOVA) was run with the aid of GenAlEx v. 6.1, establishing two hierarchical levels: (i) among taxonomic groups (species), (ii) among populations, and within populations. Third, a Principal Coordinates Analysis (PCoA) at population level was carried out with the same software (GenAlEx). Fourth, putative genetic barriers between populations were detected with the software Barrier v. 2.272; significance of barriers was tested by bootstraping 1000 DA73 matrices that were previosuly obtained with Microsatellite Analyzer (MSA) v. 4.05 software74.
Gene flow was estimated with two time-frameworks. First, we estimated recent (i.e. within the recent 2–3 generations) migration rates between individual populations with the sofware BayesAss v. 1.375. As program settings, the default values were used (MCMC iterations, 3 × 106; length of the burn-in, 999,999; sampling frequency, 2000; delta value, 0.15). Second, historical mutation-scaled migration rates (M = m/μ, where m is migration rate and μ is mutation rate per generation) were estimated using MIGRATE-N v. 3.6.476. Ten replicates were run under a Brownian motion model, assuming constant mutation rate for all loci. With a Bayesian approach, a long chain with 20,000 genealogies to sample was run, with a sampling increment of 100 (thus, totalling 2,000,000 genealogies for each replicate); the burn-in was set to 20,000. A static heating scheme was chosen (temperatures were specified to 1.00, 1.50, 3 and 1 × 106), with uniform prior distribution both for Θ and M (min: 0; max: 100; delta: 10). The effective number of migrants per generation (Nm) among populations was estimated using the formula 4Nm = ΘM 76. We could not estimate migration rate (m) values, provided that those obtained from MIGRATE-N are mutation-scaled (M), and mutation rates (μ) for microsatellites for the genus Centaurea are not available. Total immigration and emigration rates for each population were obtained by summing values of Nm. Analyses were carried out at the CIPRES bioinformatic facility77.
Ecological niche modelling
Ecological niche modelling (ENM) was performed to evaluate the potential distribution of the Turkey species of Centaurea under present climatic conditions and to project it to the Last Glacial Maximum (LGM, ca. 21,000 yr BP). The ENM was only performed with two of the study species (C. cadmea and C. luschaniana) because for the other taxa the number of wild occurrences was not enough to get reliable models (<5)78. We employed the maximum entropy algorithm, as implemented in MaxEnt v. 3.3 79. The current distribution information for both Centaurea species was obtained from presence records included in the Global Biodiversity Information Facility (www.gbif.org), from literature12,14, and from the sampling sites of this study. In total, after removing duplicate records within each pixel (2.5 arc-min, ca. 5 km), we obtained 8 and 5 presence records for C. cadmea and C. luschaniana, respectively. A set of 19 bioclimatic variables at 2.5 arc-min resolution covering the distribution range (and neighboring areas) for both species under current conditions (1950–2000) were downloaded from the WorldClim website (www.worldclim.org). After a correlation analysis in a random sample of 10,000 points within the study area, we selected a smaller set of nine (relatively) uncorrelated (r ≥ |0.9|) variables: isothermality (bio3), temperature seasonality (bio4), mean temperature of the wettest quarter (bio8), mean temperature of the warmest quarter (bio10), mean temperature of the coldest quarter (bio11), annual precipitation (bio12), precipitation of the driest month (bio14), precipitation seasonality (bio15), and precipitation of the coldest quarter (bio19). The selection of variables from pairs or groups of highly correlated ones was done on the basis of their relative contribution to the model (percent contribution, jackknife tests of variable importance). The distribution model under current conditions was projected to the LGM using palaeoclimatic layers simulated by both the Community Climate System Model (CCSM)80 and the Model for Interdisciplinary Research on Climate (MIROC)81. Replicate runs (10) of MaxEnt (using the “bootstrap” method) were performed to ensure reliable results.
Model testing when the number of occurrences is small (<25) can be problematic, because the “training” and “test” datasets would be too low78. To overcome this, we used a methodology based on a jackknife (or “leave-one-out”) procedure78 to test the model. With this procedure the model is built (or “trained”) using n − 1 occurrences, and tested using the discarded locality (that is, by evaluating the ability to predict the single locality excluded from the training dataset). Thus, eight and five predictions were obtained for C. cadmea and C. luschaniana, respectively. We used the “lowest presence threshold” (LPT, also commonly referred as “minimum training presence” in the MaxEnt terminology) as the cut-off value to decide whether the discarded locality is “suitable” or “unsuitable”. The LPT is considered as more conservative than other approaches (such as the 10% fixed threshold) since identifies the minimum predicted area possible whilst maintaining zero omission error in the training data set78. Performance of models for both C. cadmea and C. luschaniana was evaluated through success rate (q, which is the proportion of right predictions) and statistical significance (a P-value computed across the set of jackknife predictions), which was done using the software provided by Pearson et al.78. The jackknife analysis indicated that the model was successful at predicting known occurrences as suitable areas for C. cadmea (success rate = 75%, P < 0.001) but not for C. luschaniana (success rate = 40%, P = 0.041). Thus, definitive ENM models (that is, using all occurrence points) for the present and LGM were only built for C. cadmea. Before it, however, we carried out tuning experiments varying the level of regularization, in order to improve the performance of the model82. After discarding regularization multipliers >2 (as they diminished AUC values considerably according to exploratory runs), leave-one-out models were built with up to eight regularization multipliers (0.25, 0.50, 0.75, 1.00, 1.25, 1.50, 1.75, and 2.00). Based on the optimal combination of AUC values and success rates (q)83, the best model was that using a regularization multiplier of 0.5 (Supplementary Fig. S4).
To convert continuous suitability values obtained for each pixel to presence/absence (that is, a binary map), we chose the minimum training presence value as the threshold. Finally, a jackknife analysis was used to evaluate the relative importance of the nine bioclimatic variables employed. All ENM predictions were visualized in ArcGIS v. 9.3 (ESRI, Redlands, CA, USA), with the aid of Hawth’s Analysis Tools84.
Additional Information
How to cite this article: López-Pujol, J. et al. Speciation and genetic diversity in Centaurea subsect. Phalolepis in Anatolia. Sci. Rep. 6, 37818; doi: 10.1038/srep37818 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000).
Thompson, J. D., Lavergne, S., Affre, L., Gaudeul, M. & Debussche, M. Ecological differentiation of Mediterranean endemic plants. Taxon 54, 967–976 (2005).
Davis, P. H. Distribution patterns in Anatolia with particular references to endemism in Plant life of south-west Asia (eds. Davis, P. H., Harper, P. C. & Hedge, I. C. ) 15–27 (Botany Society of Edinburgh, 1971).
Médail, F. & Quézel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 84, 112–127 (1997).
Takhtajan, A. Floristic regions of the world (University of California Press, 1986).
Médail, F. & Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeog. 36, 1333–1345 (2009).
Susanna, A. & Garcia-Jacas, N. Tribe Cardueae in The families and genera of vascular plants (eds. Kadereit, J. W. & Jeffrey, C. ) 123–147 (Springer, 2007).
Garcia-Jacas, N. et al. Centaurea revisited: A molecular survey of the Jacea group. Ann. Bot. 98, 741–753 (2006).
Hilpold, A. et al. Phylogeny of the Centaurea group (Centaurea, Compositae) – geography is a better predictor than morphology. Mol. Phylogenet. Evol. 77, 195–215 (2014).
Türe, C. & Böcük, H. Distribution patterns of threatened endemic plants in Turkey: A quantitative approach for conservation. J. Nat. Conserv. 18, 296–303 (2010).
Wagenitz, G. Centaurea L. in Flora of Turkey and the East Aegean Islands, Vol. 5 (ed. Davis, P. H. ) 465–585 (Edinburgh University Press, 1975).
Uysal, T. Centaurea L. In Türkiye bitkileri listesi (Damarlı bitkiler) [A check list of the flora of Turkey (Vascular plants)] (eds. Güner, A., Aslan, S., Ekim, T., Vural, M. & Babaç, M. T. ) 127–140 (Nezahat Gökyiğit Botanik Bahçesi ve Flora Araştırmaları Derneği Yayını, 2012) [in Turkish].
Garcia-Jacas, N. et al. The polyploid series of Centaurea toletana: glacial migrations and introgression revealed by nrDNA and cpDNA sequence analyses. Mol. Phylogenet. Evol. 52, 377–394 (2009).
Köse, Y. B., Alan, S. & Yücel, E. Comparative investigation of the morphological characteristics of species belonging to the Centaurea L. section Phalolepis (Cass.) DC. Biol. Divers. Conserv. 3, 10–22 (2010).
López-Vinyallonga, S., López-Pujol, J., Constantinidis, T., Susanna A. & Garcia-Jacas, N. Mountains and refuges: genetic structure and evolutionary history in closely related, endemic Centaurea in continental Greece. Mol. Phylogenet. Evol. 92, 243–254 (2015).
Fréville, H., Justy, F. & Olivieri, I. Comparative allozyme and microsatellite population structure in a narrow endemic plant species, Centaurea corymbosa Pourret (Asteraceae). Mol. Ecol. 10, 879–889 (2001).
Mameli, G., Filigheddu, R., Binelli, G. & Meloni, M. The genetic structure of the remnant populations of Centaurea horrida in Sardinia and associated islands. Ann. Bot. 101, 633–640 (2008).
Guarino, C., Salerno, G. & Cipriani, G. Effects of fragmentation phenomena on the genetic structure and gene flow in Centaurea cineraria group (Asteraceae) in the Mediterranean Basin. Plant Biosyst. 147, 996–1005 (2013).
Marrs, R. A., Sforza, R. & Hufbauer, R. A. Evidence for multiple introductions of Centaurea stoebe micranthos (spotted knapweed, Asteraceae) to North America. Mol. Ecol. 17, 4197–4208 (2008).
Marrs, R. A., Sforza, R. & Hufbauer, R. A. When invasion increases population genetic structure: a study with Centaurea diffusa. Biol. Invasions 10, 561–572 (2008).
Huang, P. & Schaal, B. A. Association between the geographic distribution during the last glacial maximum of Asian wild rice, Oryza rufipogon (Poaceae), and its current genetic variation. Am. J. Bot. 99, 1866–1874 (2012).
Dakin, E. E. & Avise, J. C. Microsatellite null alleles in parentage analysis. Heredity 93, 504–509 (2004).
Babbel, G. R. & Selander, R. K. Genetic variability in edaphically restricted and widespread plant species. Evolution 28, 619–630 (1974).
Frankham, R. Genetics and extinction. Biol. Conserv. 126, 131–140 (2005).
Hamrick, J. L. & Godt, M. J. W. Allozyme diversity in plant species in Plant population genetics, breeding and genetic resources (eds. Brown, A. H. D., Clegg, M. T., Kahler, A. L. & Weir, B. S. ) 43–63 (Sinauer Associates, 1990).
Gitzendanner, M. A. & Soltis, P. S. Patterns of genetic variation in rare and widespread plant congeners. Am. J. Bot. 87, 783–792 (2000).
Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 13, 1143–1155 (2004).
López-Pujol, J., Martinell, M. C., Massó, S., Blanché, C. & Sáez, L. The ‘paradigm of extremes’: Extremely low genetic diversity in an extremely narrow endemic species, Coristospermum huteri (Umbelliferae). Plant Syst. Evol. 299, 439–446 (2013).
Lavergne, S., Thompson, J. D., Garnier, E. & Debussche, M. The biology and ecology of narrow endemic and widespread plants: a comparative study of trait variation in 20 congeneric pairs. Oikos 107, 505–518 (2004).
Quilichini, A., Debussche, M. & Thompson, J. D. Geographic differentiation of morphological traits and isozymes in the Mediterranean island endemic Anchusa crispa: implications for the conservation of a protected species. Biol. Conserv. 118, 651–660 (2004).
Garrido, J. L., Fenu, G., Mattana, E. & Bacchetta, G. Spatial genetic structure of Aquilegia taxa endemic to the island of Sardinia. Ann. Bot. 109, 953–964 (2012).
Fineschi, S., Cozzolino, S., Migliaccio, M. & Vendramin, G. G. Genetic variation of relic tree species: the case of Mediterranean Zelkova abelicea (Lam.) Boisser and Z. sicula Di Pasquale, Garfì and Quézel (Ulmaceae). For. Ecol. Manag. 197, 273–278 (2004).
Fernández-Mazuecos, M., Jiménez-Mejías, P., Rotllan-Puig, X. & Vargas, P. Narrow endemics to Mediterranean islands: moderate genetic diversity but narrow climatic niche of the ancient, critically endangered Naufraga (Apiaceae). Perspect. Plant Ecol. Evol. Syst. 16, 190–202 (2014).
Jiménez-Mejías, P., Fernández-Mazuecos, M., Amat, M. E. & Vargas, P. Narrow endemics in European mountains: high genetic diversity within the monospecific genus Pseudomisopates (Plantaginaceae) despite isolation since the late Pleistocene. J. Biogeogr. 42, 1455–1468 (2015).
Gray, A. Genetic diversity and its conservation in natural populations of plants. Biodivers. Lett. 3, 71–80 (1996).
Marsden-Jones, E. M. & Turrill, W. B. British Knapweeds: a study in synthetic taxonomy (The Ray Society, 1954).
Jeffrey, C. Evolution of Compositae flowers in Systematics, evolution, and biogeography of Compositae (eds. Funk, V. A., Susanna, A., Stuessy, T. F. & Bayer, R. J. ) 293–313 (IAPT, 2009).
Pisanu, S., Filigheddu, R. & Farris, E. The conservation status of an endemic species of northern Sardinia: Centaurea horrida Badarò (Asteraceae). Plant Biosyst. 143, 275–282 (2009).
López-Alvarado, J. Centaurea L. section Phrygia Pers.: Phylogeny and biogeography (PhD Thesis, Università degli Studi di Sassari, 2012).
Sarıkaya M. A., Çiner, A. & Zreda, M. Quaternary glaciations of Turkey in Quaternary glaciations—Extent and chronology. A closer look (eds. Ehlers, J., Gibbard, P. L. & Hughes, P. D. ) 393–403 (Elsevier, 2011).
Prentice, I. C., Harrison, S. P. & Bartlein, P. J. Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytol. 189, 988–998 (2011).
Kageyama, M. et al. Mid-Holocene and last glacial maximum climate simulations with the IPSL model: part II: model-data comparisons. Clim. Dyn. 40, 2469–2495 (2013).
Şenkul, Ç. & Doğan, U. Vegetation and climate of Anatolia and adjacent regions during the Last Glacial period. Quatern. Int. 302, 110–122 (2013).
Nieto Feliner, G. Patterns and processes in plant phylogeography in the Mediterranean Basin. A review. Perspect. Plant Ecol. Evol. Syst. 16, 265–278 (2014).
Duminil, J. & di Michele, M. Plant species delimitation: a comparison of 838 morphological and molecular markers. Plant Biosyst. 143, 528–542 (2009).
Funk, D. J. & Omland, K. E. Species-level paraphyly and polyphyly: Frequency, causes, and consequences, with insights from animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 34, 397–423 (2003).
Gutiérrez Larena, B., Fuertes Aguilar, J. & Nieto Feliner, G. Glacial-induced altitudinal migrations in Armeria (Plumbaginaceae) inferred from patterns of chloroplast DNA haplotype sharing. Mol. Ecol. 11, 1965–1974 (2002).
Miebach, A., Niestrath, P., Roeser, P. & Litt, T. Impacts of climate and humans on the vegetation in NW Turkey: palynological insights from Lake Iznik since the Last Glacial. Clim. Past Discuss. 11, 5157–5201 (2015).
Biltekin, D. et al. Anatolia: A long-time plant refuge area documented by pollen records over the last 23 million years. Rev. Palaeobot. Palynol. 215, 1–22 (2015).
Körner, C. & Ohsawa, M. Mountain systems in Ecosystems and human well-being, current state and trends. Findings of the Condition and Trends Working Group of the Millennium Ecosystem Assessment, Vol. 1 (eds. Hassan, R., Scholes, R. & Ash, N. ) 681–716 (Island Press, 2005).
Verlaque, R., Médail, F., Quézel, P. & Babinot, J. F. Endémisme végétal et paléogéographie dans le bassin Méditerranéen. Geobios 21, 159–166 (1997).
Mace, G. M., Gittleman, J. L. & Purvis A. Preserving the tree of life. Science 300, 1707–1709 (2003).
Şekercioğlu, Ç. H. et al. Turkey’s globally important biodiversity in crisis. Biol. Conserv. 144, 2752–2769 (2011).
Davis, E. B. et al. The California Hotspots Project: Identifying regions of rapid diversification of mammals. Mol. Ecol. 17, 120–138 (2008).
Andreakis, N., Kooistra, W. H. C. F. & Procaccini, G. High genetic diversity and connectivity in the polyploid invasive seaweed Asparagopsis taxiformis (Bonnemaisoniales) in the Mediterranean, explored with microsatellite alleles and multilocus genotypes. Mol. Ecol. 18, 212–226 (2009).
Ekim, T. et al. (egrelti ve tohumlu bitkiler) [Red data book of Turkish plants (Pteridophyta and Spermatophyta)] (Turkish Association for the Conservation of Nature-Van Centennial University, 2000) [in Turkish].
IUCN. IUCN Red List Categories and Criteria: Version 3.1. (IUCN Species Survival Commission, 2001).
Duran, A. & Duman, H. Two new species of Centaurea (Asteraceae) from Turkey. Ann. Bot. Fennici 39, 43–48 (2002).
Uysal, T., Özel, E., Bozkurt, M. & Ertuğrul, K. Genetic diversity in threatened populations of the endemic species Centaurea lycaonica Boiss. & Heldr. (Asteraceae). Res. J. Biol. 2, 110–116 (2012).
Doyle, J. J. & Dickson, E. E. Preservation of plant samples for DNA restriction endonuclease analysis. Taxon 36, 715–722 (1987).
Fréville, H., Imbert, E., Justy, F., Vitalis, R. & Olivieri, I. Isolation and characterization of microsatellites in the endemic species Centaurea corymbosa Pourret (Asteraceae) and other related species. Mol. Ecol. 9, 1671–1672 (2000).
Marrs, R. A., Hufbauer, R. A., Bogdanowicz, S. M. & Sforza, R. Nine polymorphic microsatellite markers in Centaurea stoebe L. [subspecies C. s. stoebe and C. s. micranthos (S. G. Gmelin ex Gugler) Hayek] and C. diffusa Lam. (Asteraceae). Mol. Ecol. Notes 6, 897–899 (2006).
Peakall, R. & Smouse, P. E. GenAlEx 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6, 288–295 (2006).
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N. & Bonhomme, F. GENETIX 4. 05, Logiciel sous Windows™ pour la génétique des populations (Laboratoire Génome, Populations, Interactions, CNRS UMR 5000, Université de Montpellier II, 1996–2004).
Weir, B. S. & Cockerham, C. C. Estimating F-statistics for the analysis of population structure. Evolution 38, 1358–1370 (1984).
Raymond, M. & Rousset, F. GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J. Hered. 86, 248–249 (1995).
Raymond, M. & Rousset, F. An exact test for population differentiation. Evolution 49, 1283–1286 (1995).
Chapuis, M.-P. & Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 24, 621–631 (2007).
Pritchard, J. K., Wen, X. & Falush, D. Documentation for Structure software: version 2. 3. (Department of Human Genetics, University of Chicago). Available at: http://pritch.bsd.uchicago.edu/structure_software/release_versions/v2.3.4/structure_doc.pdf (Accessed 24th August 2015), (2010).
Evanno, G., Regnaut, S. & Goudet, J. Detecting the number of clusters of individuals using the software Structure: a simulation study. Mol. Ecol. 14, 2611–2620 (2005).
Earl, D. A. & vonHoldt, B. M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resources 4, 359–361 (2012).
Manni, F., Guérard, E. & Heyer, E. Geographic patterns of (genetic, morphologic, linguistic) variation: how barriers can be detected by Monmonier’s algorithm. Hum. Biol. 76, 173–190 (2004).
Nei, M., Tajima, F. & Tateno, Y. Accuracy of estimated phylogenetic trees from molecular data. J. Mol. Evol. 19, 153–170 (1983).
Dieringer, D. & Schlötterer, C. Microsatellite analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 3, 167–169 (2003).
Wilson, G. A. & Rannala, B. Bayesian inference of recent migration rates using multilocus genotypes. Genetics 163, 1177–1191 (2003).
Beerli, P. & Felsenstein, J. Maximum likelihood estimation of a migration matrix and effective population sizes in n subpopulations by using a coalescent approach. Proc. Natl. Acad. Sci. USA 98, 4563–4568 (2001).
Miller, M. A., Pfeiffer, W. & Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees in Proceedings of the Gateway Computing Environments Workshop (GCE), 14 Nov. 2010, 1–8 (2010).
Pearson, R. G., Raxworthy, C. J., Nakamura, M. & Peterson, A. T. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J. Biogeog. 34, 102–117 (2007).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259 (2006).
Collins, W. D. et al. The community climate system model version 3 (CCSM3). J. Clim. 19, 2122–2143 (2006).
Hasumi, H. & Emori, S. K-1 Coupled GCM (MIROC) Description. K-1 Technical Report 1 (Center for Climate System Research. University of Tokyo, 2004).
Radosavljevic, A. & Anderson, R. P, Making better Maxent models of species distributions: complexity, overfitting and evaluation. J. Biogeogr. 41, 629–643 (2014).
Zhao, J.-L., Gugger, P. F., Xia, Y.-M. & Li, Q.-J. Ecological divergence of two closely related Roscoea species associated with late Quaternary climate change. J. Biogeogr. 43, 1990–2001 (2016).
Beyer, H. L. Hawth’s analysis tools for ArcGIS, Available at: http://www.spatialecology.com/htools (Accessed: 5th August 2015) (2004).
Acknowledgements
This work was supported by the Spanish Ministerio de Ciencia e Innovación [project CGL2010/18631] and Generalitat de Catalunya [Ajuts a Grups de Recerca Consolidats 2014-SGR514-GREB].
Author information
Authors and Affiliations
Contributions
A.S., K.E., T.U. and O.T. collected samples; S.L.-V., A.G. and N.G.-J. conducted the experiments; J.L.-P., S.L.-V. and N.G.-J. analyzed the data; A.S. and N.G.-J. designed the study; J.L.-P., A.S. and N.G.-J. led the writing of the manuscript; K.E., T.U. contributed to the writing. All authors read and reviewed the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
López-Pujol, J., López-Vinyallonga, S., Susanna, A. et al. Speciation and genetic diversity in Centaurea subsect. Phalolepis in Anatolia. Sci Rep 6, 37818 (2016). https://doi.org/10.1038/srep37818
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep37818
This article is cited by
-
The Centaurea alba complex in the Iberian Peninsula: gene flow, introgression, and blurred genetic boundaries
Plant Systematics and Evolution (2020)
-
Do climate-driven altitudinal range shifts explain the intraspecific diversification of a narrow ranging montane mammal, Taurus ground squirrels?
Mammal Research (2018)
-
Assessing the Relevance of Herbarium Collections as Tools for Conservation Biology
The Botanical Review (2017)
-
Revisiting the provenance delineation of a widespread shrub, Frangula alnus—the role of spatial, temporal and environmental patterns
Tree Genetics & Genomes (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
