
Abstract
It is well established that emotions influence our decisions, yet the neural basis of this biasing effect is not well understood. Here we directly recorded local field potentials from the OrbitoFrontal Cortex (OFC) in five human subjects performing a financial decision-making task. We observed a striking increase in gamma-band (36–50 Hz) oscillatory activity that reflected subjects’ decisions to make riskier choices. Additionally, these gamma rhythms were linked back to mismatched expectations or “luck” occurring in past trials. Specifically, when a subject expected to win but lost, the trial was defined as “unlucky” and when the subject expected to lose but won, the trial was defined as “lucky”. Finally, a fading memory model of luck correlated to an objective measure of emotion, heart rate variability. Our findings suggest OFC may play a pivotal role in processing a subject’s internal (emotional) state during financial decision-making, a particularly interesting result in light of the more recent “cognitive map” theory of OFC function.
Similar content being viewed by others


Introduction
The decisions we make on a daily basis are heavily influenced by our emotions. Phenomena like loss aversion or risk sensitivity can bias decisions away from maximization of expected reward1,2. Emotional influences can also be seen through more superficial biases, like that of an attractive face3. From a behavioral perspective, ways that emotions can bias our decisions have been extensively studied (refs 4, 5, 6 to name a few). However, the specifics of how these biases are represented in the brain are still not well understood. Considerable physiological and anatomical evidence suggests that the OrbitoFrontal Cortex (OFC) is involved in both decision-making and emotional processing.
Activity in the OFC is associated with value encoding of choices7,8, stimuli9, discrepancies between realized and expected results10, and compulsive decision-making11. In addition, lesions of the OFC have been shown to influence a person’s ability to perform reversal learning12,13 and to impair the ability to incorporate emotional cues into his or her decisions14. Other lesion studies, such as those in macaques15, support the idea the OFC helps assign credit for outcomes to previously made decisions9,16,17. Single unit work in rats further indicates a strong link between OFC and stimulus-outcome learning over action-outcome learning18.
While a diverse set of explanations have arisen to explain the extensive body of OFC related work, recently studies have emerged supporting the idea OFC functions as a sort of “cognitive map”19. That is, the OFC maintains or provides key information for a state-space description of a task and reflects the organism’s location within it, particularly aspects that are not reflected in the environment. Single unit recordings20 and other neuropsychological data21 indicate the OFC contains complex information regarding internal choice allowing the disambiguation of states that seem similar externally but result in different outcomes. In this paper, we present data suggesting that OFC activity contains information dissociating between the choices of two equivalent options in the face of identical external stimuli, reflecting a bias stemming from an internal state—possibly an internal emotional state.
We accomplished this by analyzing local field potentials recorded directly from the human OFC during a gambling task as a means of examining relationships between OFC activity, inferred emotional state, and decision-making. The OFC is typically studied using human case studies, functional Magnetic Resonance Imaging (fMRI) in humans, lesion experiments in animal models, or electrophysiology in monkeys or rats. Typical concerns associated with interpreting fMRI studies are the poor temporal resolution and the blood oxygenation level-dependent signal only being a proxy for neural activity22. On the other hand, conclusions drawn from rat and monkey studies suffer from difficulties in translation to humans23. By taking advantage of Stereotactic ElectroEncephaloGraphy (SEEG) recordings in epileptic patients at the Cleveland Clinic24, we were able to examine local field potential activity of the OFC in five humans at high temporal resolution (1 or 2 kHz). With this technique, we can both analyze quick neural changes as well as draw straightforward conclusions for human OFC function. While OFC activity has always been studied in response to a stimulus or reward, we are able to show that gamma rhythms in OFC present before any stimulus is shown may influence behavior. We are further able to link these gamma rhythms back to the subjects’ heart rate variability (HRV)—a proxy to emotion—and mismatched expectations in past trials, supporting the idea the OFC reflects the current state with respect to the task using internal information.
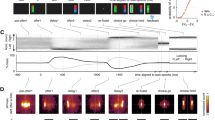
Patients implanted with SEEG for seizure localization had the opportunity to participate in this decision-making study25. All patients in our study signed a Cleveland Clinic Institutional Review Board approved informed consent before participation. Five subjects volunteered who happened to have SEEG electrodes in OFC. The task involved a gambling decision (Fig. 1a). Subjects played a game of high card where they would win virtual money if their card beat the computer’s card. Specifically, in the beginning of each trial, the subject controls a cursor via a planar manipulandum (Interactive Motion Technologies, USA) to a fixation target. Afterwards, subjects are shown their card (2, 4, 6, 8 or 10) that is randomly chosen with equal distribution (known by the subjects). The computer’s card is initially hidden. The screen then shows their two choices, a high bet ($20) and a low bet ($5). The subject has 6 seconds to select one with their cursor. Following selection, the computer’s card, which follows the same distribution, is revealed. The final screen depicts the amount won or lost. See Subjects and Gambling Task in Methods for more details.

Gambling task and behavioral results.
(a) Timeline of the behavioral task. After fixation, subjects were shown their card. Once the bets were shown, subjects selected one of the choices and then were shown the computer’s card following a delay. Feedback was provided afterwards by displaying the amount won or lost. (b) Average bet decisions across cards. Subjects predominantly bet low for 2 and 4 cards and bet high for 8 and 10 cards. There was no predominant strategy for 6 cards, which had a 29% chance of eliciting a high bet. (c) Average normalized reaction times across cards. Subjects reacted slower for cards whose rewards had higher variability. Error bars represent one standard error of the mean.
Results
The optimal strategy for maximizing expected reward in this task is straightforward: bet high when dealt the 8 or 10 card and bet low when dealt the 2 or 4 card. Subject behavior closely followed this strategy (Fig. 1b). In contrast, for the 6 card, both betting options resulted in the same expected reward. The ambiguity of the decision on 6-card trials resulted in subjects switching between both high and low bets and taking longer to decide (Fig. 1c).
For their decisions, subjects on average bet high 29% of the time for these 6-card trials, thus exhibiting risk-averse behavior. Given that a decision must be made despite both decisions having the same expected reward, decisions are likely based on some internal bias. We hypothesize that the choice on each 6-card trial is driven by the subject’s internal state that fluctuates on a trial-by-trial basis and that this internal bias is mediated by the OFC.
Linking OFC to an internal bias that predicts future behavior
We first tested the link between gambling decisions on these 6-card trials (i.e., trials with the greatest uncertainty of a prospective win) and oscillatory power in OFC local field potentials. Oscillatory power was used due to its common association with synchronized activities of the underlying neuronal population encoding behavior26. See Spectral Analysis in Methods for more details concerning the calculation of oscillatory power27. To ensure bias encoding instead of decision encoding, neural oscillations were analyzed before subjects even saw their card at the beginning of each trial. To determine whether any OFC activity correlated to betting decisions, OFC oscillatory power was compared between the set of trials where subjects end up betting high on a 6 card and the set where they end up betting low. The average normalized spectrograms for both high and low bet trials show that high bet trials have higher 40–50 Hz oscillatory power about 1000 milliseconds (ms) preceding the show card epoch (Fig. 2a). To determine statistical significance of this effect, we used a cluster-based nonparametric statistical test28. Clusters are defined as a set of adjacent time-frequency windows whose activity is different between trials where the subjects end up betting high versus low. See Cluster-Based Nonparametric Statistical Test in Methods for more details.

Oscillatory power before the Show Card.
(a) The average spectrograms show differences between high-bet and low-bet conditions on 6-card trials. One significant cluster (p = 0.042) resulted from the cluster-based nonparametric statistical test. The cluster contained frequencies between 36 and 50 Hz at a timing between 1000 and 800 ms before the Show Card. This frequency range matches the traditional lower gamma band. Plots of average oscillatory power (36–50 Hz) over time for 6-card trials resulting in high and low bets show the modulation of the power in the gamma band preceding the Show Card. Time bins with significant differences are marked by the grey bar. Error bars represent one standard error of the mean. The number n denotes the number of trials pooled across patients. (b) The average spectrograms show differences between high-luck and low-luck conditions on all trials. One significant cluster (p = 0.040) resulted from the cluster-based nonparametric statistical test. The cluster is located in the similar time-frequency region as the cluster emerging from the high-bet and low-bet conditions on 6-card trials.
The most significant cluster emerged between 36–50 Hz around 1000 and 800 ms before the card is shown (p = 0.042). It had significantly higher power during 6-card trials where the subjects ended up betting high (Fig. 2a, third panel). Comparing the average gamma power, as defined by this frequency range, shows that the power is significantly higher 1000 to 800 ms before the subject even sees the 6 card on trials where the subject ends up betting high (Fig. 2a, fourth panel). Additionally, we tested if the modulation of OFC oscillatory power in the same time period was merely a result of other effects, including effects from outcome or reward processing in the previous trial. No significant clusters were found (see data in Supplementary Information). Future references to “OFC activity” in the remainder of this report relate to the average power in the cluster defined by 1000–800 ms before the show card and 36–50 Hz (red rectangle in Fig. 2a, third panel).
The influence of OFC activity was then assessed for each card type individually by constructing a model of the probability of betting high on a given card. This test was performed to see if this OFC activity improves prediction of betting behavior on any card other than the 6-card trials, and the model will be compared to models constructed in remaining analyses below. Using a logistic function to ensure probability values between 0 and 1, the subject’s betting behavior was modeled as:

where pc(k) is the probability of betting high on card c = {2, 4, 8, 10} for trial k, and OFC(k) is the subject’s OFC activity for trial k. The model parameters ac and γc are fitted using maximum likelihood estimation for each card separately29. The coefficients γc for c = {2, 4, 8, 10} were found to be not significant, indicating that OFC activity only statistically significantly biases behavior during the ambiguous trials on the 6-card. The p-values associated with OFC influence on 2, 4, and 8 cards saturate to 1, and the p-value for 10-card trials is 0.18 (after applying a Bonferroni correction to compensate for the four comparisons).
So far, effects of OFC bias on behavior have only been assessed through bet decisions. However, another key aspect of behavior in this task is reaction time. If OFC activity biases towards the decision made, then a faster reaction time would be expected when the OFC activity is higher; while if the opposite were true, a slower reaction time would be expected. This relationship between normalized reaction times and OFC power was tested for both high and low bet trials for 6 cards using Pearson’s correlation tests (Fig. 3). The correlation was only significant for the case of the high bets. Here, higher OFC activity, which biases towards high bets, is correlated with faster reaction times, as expected. The lack of significance for the low bet cases may suggest that OFC only selectively influences decision-making. This type of selective influence of other brain areas has been noted in other decision modalities. For example, the insular cortex selectively activates for risk-averse choices while the nucleus accumbens selectively activates for risk-seeking choices30.

Reaction time and OFC activity correlations for high (a) and low (b) bets for 6 cards. For high bets, there is a negative and significant correlation between OFC activity and reaction time, meaning that higher activity resulted in faster decisions. No significant correlation was observed in the low bet scenario. The number n denotes the number of trials pooled across patients.
Linking OFC to previous outcomes
The next step was to assess how this OFC activity linked back to outcomes of previous trials, as this may impact how the subject “feels” on a trial-by-trial basis. While not a direct measure of emotional state, we used cumulative past trial outcomes as a proxy. It was also assumed that subjects have an accurate perception of the odds of winning given a specific card. The overall behavior shown in Fig. 1b supports this. Specifically, we constructed a fading memory model of cumulative mismatched expectations that we herein refer to as “luck” L(k) on trial k. The luck variable updates as follows:

where a is a decay factor (0 ≤ a ≤ 1) and e(k) is the mismatched expectations on trial k, that is, the difference between the actual outcome (loss = −1, draw = 0, or win = 1 and expected outcome given the player card c(k) (computed as  ). Note that e(k) enters the model only during trials where expectations are mismatched. Further note how the mismatched expectations, e(k), functionally acts similarly to the error signal in reinforcement learning, as it tracks the mismatch between actual and expected outcome31 However, this analysis differs from traditional reinforcement learning as the changes in behavior are not a function of the subject’s previous actions. This focus on mismatched outcomes is in part motivated by the previously mentioned theory on reversal learning proposed in ref. 10.
). Note that e(k) enters the model only during trials where expectations are mismatched. Further note how the mismatched expectations, e(k), functionally acts similarly to the error signal in reinforcement learning, as it tracks the mismatch between actual and expected outcome31 However, this analysis differs from traditional reinforcement learning as the changes in behavior are not a function of the subject’s previous actions. This focus on mismatched outcomes is in part motivated by the previously mentioned theory on reversal learning proposed in ref. 10.
We hypothesize that luck is linked to OFC activity on a trial-by-trial basis. To test this hypothesis, we varied the decay factor a between 0 and 1 in 0.01 increments and computed the Pearson’s correlation coefficient between luck and OFC activity. The optimal decay factor was a = 0.97 (ρ = 0.13, p = 0.043) after a Bonferroni correction). This optimal decay factor was used for the remainder analyses. To further examine the correlation between OFC activity and luck, we separated the trials between high-luck and low-luck conditions (defined as the bottom third and top third of the values taken by luck variable for all patients) and computed the average normalized spectrograms for both conditions. High-luck trials showed higher OFC oscillatory power than low-luck trials (Fig. 2b, first and second panels). Interestingly, the cluster-based nonparametric statistical test identified a significant cluster (p = 0.040) in the time-frequency vicinity of the cluster identified when separating trials based on high-bet and low-bets conditions on 6-card trials (Fig. 2b, third panel).
Now, knowing the link between luck and OFC activity, we tested the hypothesis that luck predicted betting decisions by modeling the probability to bet high on 6-card trials as a function of luck:

The fitted model showed that luck significantly influenced betting decisions (β6 = 0.20, p = 0.028). This indicates positive luck biases subjects to bet high on 6 cards.
By linking OFC activity, luck, and decision-making together, a possible confound is introduced. Does OFC activity actually influence future decisions or does it merely encode luck? To assess this confound, another model was used that simultaneously accounts for both the influence of luck and OFC activity on 6-card betting decisions:

After including both luck and OFC as covariates to explain the variability in 6-card bets around the baseline, only the OFC activity remains statistically significant in the model (γ6 = 0.85, p = 0.004), while luck is no longer significant (β6 = 0.17, p = 0.065). The point of the final GLM that we construct in equation (3) is to show the data suggests OFC activity carries much of the same information about betting behavior as luck and that both are redundant in the probability model. The redundancy does not refute the hypothesis that the OFC activity encodes the emotional state that then drives betting behavior, and is not surprising since we already showed that OFC activity and luck are correlated.
Linking previous outcomes to emotional state
In order to clarify luck’s link to the subject’s emotional state, we compared the signal L to each subject’s HRV signal, which is an objective measure of a person’s emotional state32. Specifically, it measures the effects of both parasympathetic and sympathetic systems on the subject32. HRV for each trial was calculated from the variance of the interbeat intervals preceding that trial and was measured through concurrently recorded ElectroCardioGrams (ECG)33,34. HRV was found to be significantly correlated with luck (ρ = −0.09, p = 0.039). This result suggests that luck is an appropriate proxy to emotional processing. See Heart Rate Variability in Methods for further details on the calculation and analysis.
Discussion
In this study, we find that OFC activity encodes an internal bias that is predictive of betting behavior on ambiguous trials where simple reason (e.g., maximizing expected reward) cannot drive the decision; and, this activity also correlates with reaction times of bets when subjects bet “high” on these trials. Then, we show that this OFC “bias” signal correlates to previous outcomes of mismatched expectations which are taken as a proxy to the subject’s emotional state. Finally, we show that cumulative mismatched expectations correlate to HRV, which is an objective measure of emotional state.
As mentioned above, we observe OFC activity that precedes any task information biases future decisions. Looking at the OFC as a biasing agent helps shed new light on the exact role of the OFC in the decision-making process. Two popular theories for the role of the OFC are response inhibition and the emotion marker theory. For the former theory, lesions interfere with reversal learning as well as go/no-go regulation35. Additionally, patients with obsessive-compulsive disorder have been linked to having a dysfunctional OFC11. However, response inhibition can be produced in some scenarios even with OFC damage, so this likely is not its fundamental role36.
For the emotional marker theory, OFC lesions are associated with an inability to link past emotions to future decisions37. Specifically, subjects with lesions performing the Iowa gambling task still had somatic markers associated with making risky choices but never ended up being dissuaded against them14. More recent work, however, has shown that this influence is only prevalent in situations with model-based learning as opposed to more heuristically-derived choices37.
Both of these theories make sense in the new context of bias encoding as a result of mismatched expectations. A dysfunctional OFC can no longer regulate bias correctly. This results in dysfunctional biasing of behavior that could manifest as failure to adapt and failure to control compulsions. These deficits can be linked to both ideas of emotional processing and response inhibition. It also answers key concerns for both theories. This biasing occurs as a result of mismatches between realizations and expectations from previous trials, which can only be present when there is an expectation in place. This could explain why learning and response inhibition are possible in situations that lack prior training, despite OFC lesions.
While the results described here can be put in the context of prior theories of OFC function, they fit most cleanly into the recent description of the OFC as containing a cognitive map19. The bias signal observed in the OFC aligns with current ideas that the OFC reflects the location of the organism within task relevant state-space. It seems to reflect an internal state, whether tied to memory or emotion, that helps explain differences in behavioral responses that are not reducible to difference in trial-specific external stimuli20,21.
The effects of the external environment on decision-making can be clearly seen in the predictable behavior of subjects when given low (2-card, 4-card) or high (8-card, 10-card) cards. However, after those effects are explained away, the question of why subjects sometimes bet high and sometimes bet low on the intermediate 6-card remains. The same stimulus leads to a differing response across trials. In this case, we proposed the existence of a hidden state, perhaps emotional, existing before the trial that would bias the subject toward betting high or low in ambiguous situations. When we treat a portion of the power spectrum of the OFC as an estimate of the hidden state, the predictions of betting behavior are improved. Interesting parallels exist with free-choice trials (similar to 6-card trials) in odor-discrimination tasks where ventral tegmental area (VTA) dopamine neurons no longer fire before the reward is acquired when the OFC has been lesioned20. In both cases, behavior is driven by an internal state rather than by an external cue. This state-space interpretation of OFC function has clear parallels with advances in control theory frequently employed in modern engineering, perhaps indicating a convergence between neuroscience and engineering theory.
Methods
Subjects
At the Cleveland Clinic, patients with medically intractable epilepsy may undergo SEEG recording in order to localize the seizure focus. In this study, aside from the behavioral experiments, no alterations were made to the patient’s clinical care including the placement of the electrodes24. All experimental protocols were approved by the Cleveland Clinic Institutional Review Board. Methods were carried out in accordance with the approved guidelines by the Cleveland Clinic Institutional Review Board. Subjects enrolled voluntarily and gave informed consent under criteria approved by the Cleveland Clinic Institutional Review Board. A total of five subjects volunteered to perform the task that had SEEG electrodes in the OFC. Details on these recordings and eventual resections of these five patients are noted in Table 1.
Subjects were implanted with 10 to 14 depth electrodes. Implantation was performed using robot-assisted surgery along with co-registered fMRI and angiograms to ensure safe implantation23. Once inserted, SEEG electrophysiological data were acquired using a Nihon Kohden 1200 EEG diagnostic and monitoring system (Nihon Kohden America, USA) at a sampling rate of 1 or 2 kHz. Behavioral data were simultaneously acquired through the MonkeyLogic Matlab toolbox27.
There are standard concerns in analyzing data from epileptic patients. First, patients are often on medication, which might affect the neurophysiology of the brain. For clinical purposes, patients were kept off of their anti-seizure medication for their entire stay at Cleveland Clinic, so these effects would be minimized. Secondly, actual seizures might impact the neurophysiology around the seizure focus. Human epilepsy recordings are taken to localize the seizure focus, so overlap is expected between seizure focus and areas recorded. We only had one subject whose seizure focus overlapped with OFC, as indicated in Table 1.
Gambling Task
The gambling task (Fig. 1a) is based on a simple game of high card where subjects would win virtual money if their card beat the computer’s card. Specifically, in the beginning of each trial, the subject controls a cursor via a planar manipulandum to a fixation target. During fixation, subjects must center the cursor in less than 8 seconds. Once centered, the subject is shown his card (2, 4, 6, 8 or 10) for a duration of 2 seconds. The card is randomly chosen with equal distribution. The computer’s card is initially hidden. The screen then shows the two possible choices: a high bet ($20) or a low bet ($5). The subject has 6 seconds to select one with his cursor. Following selection, the computer’s card, which follows the same distribution, is revealed. If the computer’s card is larger than the player’s card, then the subject loses the amount he bets. If the computer’s card is smaller than the player’s card, then the subject wins the amount he bet. If the two cards are equal, then no virtual money is won or lost. The final screen depicts the amount won or lost.
Spectral Analysis
Oscillatory power was calculated using multitapers from the Chronux toolbox27. Three orthogonal tapers were used with a 300 ms window sliding at 50 ms steps. Frequencies under 10 Hz were dropped because of the Rayleigh criterion and analyzed upwards to 100 Hz. Afterwards, each frequency bin’s power was normalized based on the power across the entire recording session by fitting the log of the power in each frequency bin to a standard normal distribution. The mean and standard deviation used for the normalization were computed from the power between the 1th and 95th percentiles of the data set. This calculation was performed for every electrode’s recording with the final normalized power being averaged across all OFC electrodes. In addition, we removed artifacts by identifying time points in the spectrograms for which the median of the absolute value of the power across all frequencies is larger than 2.5. Finally, in order to remove the effect of 60 Hz power-line noise, we ignored the frequency bins between 56.66 Hz and 63.33 Hz in all analyses.
Due to our hypothesis that there exists an internal bias before any stimulus is shown, we focused our. analysis on a time window preceding the Show Card epoch.
Cluster-Based Nonparametric Statistical Test
We used a cluster-based nonparametric statistical test to leverage the dependency between adjacent time-frequency windows in order to avoid over-penalizing with multiple comparison corrections28. For each window in the spectrogram, a null distribution was created by shuffling the condition labels 5000 times between trials within each subject. Within each shuffle, we computed a t-statistic and a p-value for each window of the newly labeled spectrograms (independent two-sample t-test with both tails, unequal sample sizes, and unequal variances). Clusters were formed by grouping windows with significant p-values (<0.05) that were adjacent in either time or frequency. The cluster-level test statistic was calculated by taking the sum of absolute values of the t-statistics for each window in the cluster. This prioritizes clusters that both have strong differences as well as large sizes. A null distribution of cluster statistics was created using the same process but with the 5000 spectrograms obtained from the originally shuffled labels. The observed cluster statistic was then compared against this null distribution of cluster statistics in order to obtain the final p-value of the test. Data from all patients were pooled together but the labels were permuted within each subject only.
Correlating OFC Activity and Reaction Time
We normalized reaction times for each patient separately by fitting the log of the reaction times to a standard normal distribution. We computed a Pearson’s correlation coefficient between the OFC activity and the normalized reaction time for all high-bet trials on 6 cards and all low-bet trials on 6 cards, respectively. The data from all patients was pooled together.
Correlating OFC Activity and Luck
To test whether OFC activity is linked to luck, we computed a Pearson’s correlation coefficient between the OFC activity and the fading memory luck variable. In order to avoid the correlation being biased toward stronger values by the earliest trials for which the luck is dominated by a few trials’ worth of outcome-prediction error (initial transient effects), the first 10 trials for each subject were ignored. Data from all patients were pooled together.
Other models of luck were considered. However, the model discussed in this paper was selected based on its alignment with past literature on the subject38,39,40, its intuitive nature, and the fact that it provides the highest correlation with OFC activity.
Generalized Linear Model Regression of Betting Behavior
We estimated the coefficients for each generalized linear regression of the responses (high and low bets) on the predictors (OFC activity and/or luck variable) using a binomial distribution and a logistic link function. When needed, we estimated these regressions for each card separately. The data from all patients was pooled together.
Correlating Luck and Heart Rate Variability
We had concurrently recorded ECG for 4 of the 5 subjects. The R peaks in the ECG signal were detected using the Pan-Tompkins algorithm33. HRV was calculated by taking the variance of interbeat intervals measured as the time between R peaks. The HRV for each trial was taken over a 5-minute window preceding the show card epoch. This window length is the established standard for HRV recording34. The HRV was normalized within each subject by dividing by the mean HRV across all trials and then by taking the logarithm as HRV distributions are often skewed right. We computed a Pearson’s correlation coefficient between the normalized HRV and the fading memory luck variable. As in the case of the correlation between luck and OFC activity, the first 10 trials for each subject were ignored to avoid initial transient effects. Data from all patients were pooled together.
Limitations of our Study
Finally, we discuss the limitations of this study.
We had relatively few patients performing our decision-making task. This is because not all patients consented and/or met the criteria of our study. The small sample size of the study population is further limited by the fact that each patient had electrodes implanted in different brain regions. In order to compensate for this small sample size, we pooled the trials over the subjects. In addition, we studied the sensitivity of the observed effects to the inclusion of each patient in the analysis: rerunning the cluster analysis leaving each subject out one at a time produces a cluster in the same time-frequency regions. This sensitivity analysis suggests that the effect truly emerges from all patients and not only from one patient (see Supplementary Information).
Additional Information
How to cite this article: Sacré, P. et al. Lucky Rhythms in Orbitofrontal Cortex Bias Gambling Decisions in Humans. Sci. Rep. 6, 36206; doi: 10.1038/srep36206 (2016).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Change history
16 February 2017
A correction has been published and is appended to both the HTML and PDF versions of this paper. The error has been fixed in the paper.
References
Kacelnik, A. & Bateson, M. Risk-sensitivity: crossroads for theories of decision-making. Trends in Cognitive Sciences 1(8), 304–309 (1997).
Tom, S. M., Fox, C. R., Trepel, C. & Poldrack, R. A. The Neural Basis of Loss Aversion in Decision-Making Under Risk. Science 315(5811), 515–518 (2007).
Kim, H., Choi, M. & Jang, I. Lateral OFC Activity predicts decision bias due to first impressions during ultimatum games. Journal of Cognitive Neuroscience 24(2), 428–439 (2011).
Pfister, H. R. & Böhm, G. The multiplicity of emotions: A framework of emotional functions in decision making. Judgment and decision making 3(1), 5–17 (2008).
Damasio, A. R. Descartes’ Error: Emotion, Reason, and the Human Brain (Grosset/Putnam, New York, NY, 1994).
Lerner, J. & Keltner, D. Beyond valence: Toward a model of emotion-specific influences on judgement and choice. Cognition and Emotion 14(4), 473–493 (2000).
Padoa-Schioppa, C. & Assad, J. A. Neurons in the orbitofrontal cortex encode economic value. Nature 441(7090), 223–226 (2006).
Levy, D. J. & Glimcher, P. W. The root of all value: a neural common currency for choice. Current Opinion in Neurobiology 22(6), 1027–1038 (2012).
Camille, N., Tsuchida, A. & Fellows, L. K. Double dissociation of stimulus-value and action-value learning in humans with orbitofrontal or anterior cingulate cortex damage. The Journal of Neuroscience 31(42), 15048–15052 (2011).
Schoenbaum, G., Saddoris, M. P. & Stalnaker, T. A. Reconciling the roles of orbitofrontal cortex in reversal learning and the encoding of outcome expectancies. Annals of the New York Academy of Sciences 1121, 320–335 (2007).
Chamberlain, S. R. et al. Orbitofrontal dysfunction in patients with obsessive-compulsive disorder and their unaffected relatives. Science 321(5887), 421–422 (2008).
Bohn, I., Giertler, C. & Hauber, W. Orbital prefrontal cortex and guidance of instrumental behavior in rats under reversal conditions. Behavioral Brain Research 143(1), 49–56 (2003).
Kim, J. & Ragozzino, M. E. The involvement of the orbitofrontal cortex in learning under changing task contingencies. Neurobiology of Learning and Memory 83, 125–133 (2005).
Bechara, A. The role of emotion in decision-making: evidence from neurological patients with orbitofrontal damage. Brain and Cognition 55(1), 30–40 (2004).
Walton, M. E., Behrens, T. E. J., Buckley, M. J., Rudebeck, P. H. & Rushworth, M. F. S. Separable Learning Systems in the Macaque Brain and the Role of Orbitofrontal Cortex in Contingent Learning. Neuron 65(6), 927–939 (2010).
Wilson, R. C., Takahashi, Y. K., Schoenbaum, G. & Niv, Y. Orbitofrontal cortex as a Cognitive Map of Task Space. Neuron 81(2), 267–279 (2014).
Camille, N., Griffiths, C. A., Vo, K., Fellows, L. K. & Kable, J. W. Ventromedial frontal lobe damage disrupts value maximization in humans. The Journal of Neuroscience 31(20), 7527–7532 (2011).
Luk, C. H. & Wallis, J. D. Choice Coding in Frontal Cortex during Stimulus-Guided or Action-Guided Decision-Making. The Journal of Neuroscience 33(5), 1864–1871 (2013).
Stalnaker, T. A., Cooch, N. K. & Schoenbaum, G. What the orbitofrontal cortex does not do. Nature Neuroscience 18, 620–627 (2015).
Takahashi, Y. K. et al. Expectancy-related changes in firing of dopamine neurons depend on orbitofrontal cortex. Nature Neuroscience 14, 1590–1597 (2011).
Kennerley, S. W. & Walton, M. E. Decision making and reward in frontal cortex: complementary evidence from neurophysiological and neuropsychological studies. Behavioral Neuroscience 125(3), 297–317 (2011).
Logothetis, N. K. What we can do and what we cannot do with fMRI. Nature Reviews 453(7197), 869–878 (2008).
Öngür, D. & Price, J. L. The organization of Networks within the Orbital and Medial Prefrontal Cortex of Rats, Monkeys and Humans. Cerebral Cortex 10(3), 206–219 (2000).
Johnson, M. et al. Performing behavioral tasks in subjects with intracranial electrodes. Journal of Visualized Experiments (92), e51947 (2014).
Cossu, M. et al. Stereoelectroencephalography in the presurgical evaluation of focal epilepsy: a retrospective analysis of 215 procedures. Neurosurgery 57(4), 706–718 (2005).
Ward, L. M. Synchronous neural oscillations and cognitive processes. Trends in Cognitive Sciences 7(12), 553–559 (2003).
Bokil, H., Andrews, P., Kulkarni, J., Mehta, S. & Mitra, P. Chronux: a platform for analyzing neural signals. Journal of Neuroscience Methods 191(1), 146–151 (2010).
Maris, E. & Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods 164(1), 177–190 (2007).
McCullagh, P. & Nelder, A. Generalized linear models, 2nd ed. (Chapman & Hall, London, UK, 1989).
Kuhnen, C. M. & Knutson, B. The neural basis of financial risk taking. Neuron 47(5), 763–770 (2005).
Holroyd, C. M. & Coles, M. G. H. The neural basis of human error processing: reinforcement learning, dopamine, and the error-related negativity. Psychological Review 109(4), 679–709 (2002).
Appelhans, B. M. & Luecken, L. J. Heart rate variability as an index of regulated emotional responding. Review of General Psychology 10(3), 229–240 (2006).
Pan, J. & Tompkins, W. J. A real-time QRS detection algorithm. IEEE Transactions on Biomedical Engineering BME-32(3), 230–236 (1985).
Gritti, I. et al. Heart Rate Variability, Standard of Measurement, Physiological Interpretation and Clinical Use in Mountain Marathon Runners during Sleep and after Acclimatization at 3480 m. Journal of Behavioral and Brain Science 3(1), 26–48 (2013).
Horn, N., Dolan, M., Elliott, R., Deakin, J. & Woodruff, P. Response inhibition and impulsivity: an fMRI study. Neuropsychologia 41(14), 1959–1966 (2003).
Chudasama, Y., Kralik, J. D. & Murray, E. A. Rhesus monkeys with orbital prefrontal cortex lesions can learn to inhibit prepotent responses in the reversed rewards contingency task. Cerebral Cortex 17(5), 1154–1159 (2007).
McDannald, M. et al. Model-based learning and the contribution of the orbitofrontal cortex to the model-free world. European Journal of Neuroscience 35(7), 991–996 (2013).
O’Doherty, J. P., Dayan, P., Friston, K., Critchley, H. & Dolan, R. J. Temporal Difference Models and Reward-Related Learning in the Human Brain. Neuron 38(2), 329–337 (2003).
O’Doherty, J. P. et al. Dissociable Roles of Ventral and Dorsal Striatum in Instrumental Conditioning. Science 304(5669), 452–454 (2004).
O’Doherty, J. P., Hampton, A. & Kim, H. Model-Based fMRI and Its Application to Reward Learning and Decision Making. Annals of the New York Academy of Sciences 1104(1), 35–53 (2007).
Acknowledgements
This is work was supported by a National Science Foundation grant (EFRI-MC3: #1137237) awarded to S.V.S., J.A.G., J.B. and J.T.G.
Author information
Authors and Affiliations
Contributions
J.G.-M. surgically implanted electrodes in subjects for clinical purposes. J.T.G., J.B., H.-J.P., M.A.J. and S.T. took part in the data collection process. J.J. and J.T.G. identified anatomical regions sampled. P.S., M.S.D.K. and K.K. performed the data analyses. S.V.S., J.T.G. and V.S.C. supervised P.S., M.S.D.K. and K.K. contributed to data analyses and organization of the manuscript. P.S., K.K., M.S.D.K. and S.V.S. wrote the manuscript text and prepared all figures. P.S. and M.S.D.K. contributed equally to this study. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sacré, P., Kerr, M., Kahn, K. et al. Lucky Rhythms in Orbitofrontal Cortex Bias Gambling Decisions in Humans. Sci Rep 6, 36206 (2016). https://doi.org/10.1038/srep36206
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep36206
This article is cited by
-
An exploratory data analysis method for identifying brain regions and frequencies of interest from large-scale neural recordings
Journal of Computational Neuroscience (2019)
-
The influences and neural correlates of past and present during gambling in humans
Scientific Reports (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
