
Abstract
Species’ functional traits are closely related to ecosystem processes through evolutionary adaptation, and are thus directly connected to environmental changes. Species’ traits are not commonly used in palaeoecology, even though they offer powerful advantages in understanding the impact of environmental disturbances in a mechanistic way over time. Here we show that functional traits of testate amoebae (TA), a common group of palaeoecological indicators, can serve as an early warning signal of ecosystem disturbance and help determine thresholds of ecosystem resilience to disturbances in peatlands. We analysed TA traits from two Sphagnum-dominated mires, which had experienced different kinds of disturbances in the past 2000 years – fire and peat extraction, respectively. We tested the effect of disturbances on the linkages between TA community structure, functional trait composition and functional diversity using structural equation modelling. We found that traits such as mixotrophy and small hidden apertures (plagiostomic apertures) are strongly connected with disturbance, suggesting that these two traits can be used as palaeoecological proxies of peatland disturbance. We show that TA functional traits may serve as a good proxy of past environmental changes, and further analysis of trait-ecosystem relationships could make them valuable indicators of the contemporary ecosystem state.
Similar content being viewed by others


Introduction
Understanding the response of ecosystems to climate and human driven processes under recent global change conditions is an important challenge for scientists1,2. Patterns of mean annual temperature and precipitation will change3 and climatic extremes may cause environmental stress and disturbance which may lead to shifts in ecosystem structure and function4,5,6. Therefore, it is vital to better understand ecosystem vulnerability and resilience to disturbances, and a long-term view of ecosystem development and dynamics using biotic proxies is especially beneficial7,8. Among endangered ecosystems, peatlands are important carbon (C) sinks9. Water deficit negatively influences their C storage capacity by modifying the hydrophysical properties of peat10, which affects peat-forming processes, potentially leading to increased C emission to the atmosphere11,12.
Anthropogenic activities, such as peat extraction, drainage and burning, are among the most common water disturbances in peatlands leading to substantial shifts in Sphagnum and vascular vegetation cover13,14, which then feedback into C fluxes15. Moreover, intensive drainage promotes prolonged anthropogenically-induced drying and significantly increases peatland vulnerability to fire16. Burning is considered to be a major force affecting various ecosystems17, and is especially important for peatlands, in which fires often cause long-lasting smouldering combustion that results in fuel consumption and substantial CO2 emission9,18. It has been lately shown that even moderate drops in the water table can lead to increased vulnerability of peatlands to wildfire16. As fire activity has significantly increased in recent years in many parts of the world19, studying the vulnerability of peatlands to fire disturbance over long time scales may provide new insights for assessing future peatland responses under global-change scenarios.
Among the different approaches used in palaeoecology, a functional trait-based approach is a useful tool to track ecosystem response to climatic and anthropogenic change20,21. Species’ functional traits have been shown to be closely related to ecosystem processes and functioning through evolutionary adaptation22,23, and they closely reflect species interactions in the communities and their response to changing environmental conditions20. Together with the analysis of functional traits, functional diversity is substantial as it represents the diversity of species functions, and is thought to be a better proxy for ecosystem functioning than the number of species per se21,24 or taxonomical diversity, which refers to the number and the relative abundance of species in the community. Additionally to conventional taxonomical analyses, species’ functional traits and functional diversity may serve as an early signal of ecosystem disturbance, helping determine thresholds of ecosystem resilience to disturbances. Such information might be crucial for nature conservation and biodiversity protection21; however, an important challenge is to choose which traits from which fossil record can be used in both palaeoecology and modern ecology to explicitly link the past with the present and unambiguously infer peatland response to environmental changes25. In the peatland microbial food web, testate amoebae (TA) are crucial for ecosystem functioning26, and they are key players in the elements cycling in soil27. TA are useful indicators of hydrological changes in peatlands28, as due to preservation of their tests it is possible to quantitatively reconstruct past water table depth29,30. TA functional traits have recently been used in ecological and palaeoecological studies of peatlands and have been linked to specific ecosystem functions: in ecological studies strong relationships have been found between moss type and TA size-structure31, and the warming effect is positively correlated to shell-aperture size over body size ratio32. Palaeoecological studies have underlined the crucial role of the shell aperture position for TA adaptation to hydrological conditions33, showed that drying eliminated large TA species from the community34, and that past atmospheric pollution and dust deposition influenced shell size and structure35.
Still, there is a scarcity of studies focusing on the long-term impact of disturbances on testate amoeba trait composition, in particular concerning effects of human-induced disturbances. Therefore, our primary goal is to examine whether the response of TA to various environmental disturbances over the past 2000 years is reflected in their trait composition. We aim to assess differences between TA trait groups in response to two disturbance agents: fire and peat extraction. We also want to define the relationship between the taxonomical and functional diversity for TA. We hypothesised that TA community composition responds to abrupt disturbance. In particular, we expected that certain traits, such as species size or aperture position, reflect shifts in peatland hydrology, based on recent findings studying the effect of water level changes33. Finally, we tested the effect of disturbances on the linkages among the structure of TA communities, the composition of functional traits and functional diversity. With this study we want to provide information about the response of TA traits to disturbances, which could support conventional microscopy analysis of TA communities.
Methods
Study sites and palaeoecological datasets
To investigate the long-term response of testate amoeba communities to disturbances, we used existing palaeoecological data sets from two Sphagnum peatlands located in Poland: Linje13 and Puścizna Krauszowska (PK)36,37. Linje, located in northern Poland (90 m a.s.l., 53°11′15″N, 18°18′34″E), is classified as a poor fen38, covering an area of 5.95 ha39. PK, located in the Orawa-Nowy Targ Basin, Polish Western Carpathians (613 m a.s.l., 49°28′06″N, 19°56′18″E)36, is a bog covering an area of 79 ha. Both peatlands were significantly disturbed by humans and those disturbances resulted in peatland drying. Environmental changes in the Linje mire were linked with the establishment of human settlement and a parallel increase in fire activity that led to substantial vegetation change in the mire area in the 14th century13. In PK, anthropogenically-induced drying appeared due to drainage followed by peat extraction that led to peat layer disruption from the 7th to the beginning of 19th century AD (this layer does not have an absolute chronology due to radiocarbon date inversion)36,37. Two hundred years ago, drainage ditches have been established on both mires, and a growth of local population was recorded in the last 100 years38,40. Such disturbances allow us to cover a large gradient which can better support our research questions and test our hypotheses.
Selected peat cores cover the last 2000 years, and possess TA community data and associated depth to water table (DWT) reconstructions, and macroscopic charcoal, plant macrofossil and pollen data13,36,37 (Supplementary Methods S1). The chronology relies on depth-age models with 20 radiocarbon dates on terrestrial plant macrofossils and 56 lead-210 dates for Linje13, and 12 radiocarbon dates on terrestrial plant macrofossils and 18 lead-210 dates for PK37 (Supplementary Table S2, Supplementary Figure S3).
Selection of testate amoeba functional traits
In order to understand the response of TA communities to the past disturbances on the group level and community level, we exploited: (1) diversity indices of the testate amoeba functional groups within the community; and (2) functional traits of the testate amoeba community.
-
1
Values of diversity indices for testate amoeba functional groups (evenness, richness and compositional change-DCA axis 1) along the sequences were calculated for selected TA groups (Fig. 1). Comparing evenness and richness is useful to reconstruct past trends in diversity and to assess how the diversity is determined by specific groups of taxa or individual species41. This approach was used to define if tendencies in certain groups can explain general changes in TA community, and to assess if particular groups of TA responses are faster than community change. TA groups were selected according to the ecological preferences of each species: preferred hydrological conditions and pH, and to the shell morphology: shell size and position of the shell aperture (acrostomic aperture – terminal, located on top of the shell, plagiostomic aperture – sub-terminal, hidden in the side of the shell, and axial aperture – located in the centre of the shell) (Supplementary Table S4).
Figure 1 
Values of diversity indices (evenness, richness, compositional change-DCA axis 1) of the testate amoeba functional groups within the testate amoeba communities calculated for Linje (a) and Puścizna Krauszowska (b) mires, associated depth to water table reconstructions (cm) and disturbance indicators: charcoal influx (particles/cm2/year, proxy for fire) and pollen indicators linked to human activities, i.e. released by ruderals (%, proxy for direct human impact, including Ambrosia artemisiifolia type, Artemisia, Brassicaceae, Chenopodiacae, Medicago lupulina type, Plantago lanceolata, P. major, P. media, Polygonum aviculare type, Rumex acetosa type, R. acetosella type, R. cf. obtusifolius, Urtica62). Disturbance periods are marked in grey.
-
2
Functional traits of the testate amoeba community were calculated for both profiles. Six traits were chosen for this study following Lamentowicz et al.33: mixotrophy, body size, biovolume, diameter of the shell aperture, position of the shell aperture within the shell and body range (Fig. 2). Descriptions of selected traits, their functions and predicted effects of disturbances on traits are listed in Table 1.
Figure 2 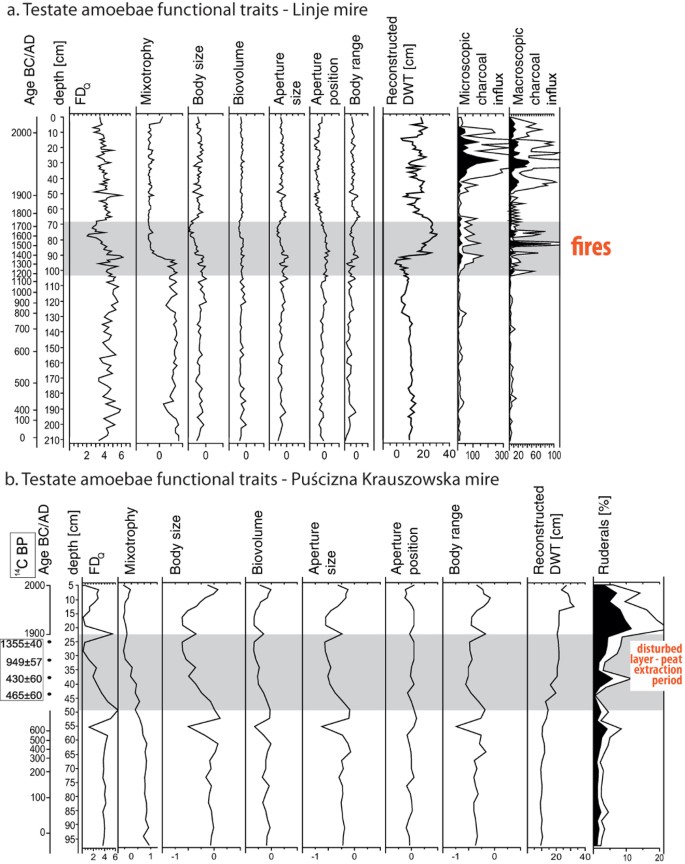
Results of functional traits and functional diversity (FDQ) of the testate amoeba communities calculated for Linje (a) and Puścizna Krauszowska (b) mires, associated depth to water table reconstructions (cm) and disturbance indicators: charcoal influx (particles/cm2/year, proxy for fire) and ruderals (%, proxy for direct human impact). Disturbance periods are marked in grey.
Table 1 Selected functional traits of testate amoebae and related detailed assumptions on how they might respond to local disturbances (drought caused by fire or peat extraction).


Numerical analyses
Based on the TA functional traits, we calculated the community weighted mean values (CWM) of each standardised trait, which is an index of functional composition expressed as the mean trait value of species present in the community, weighted by their relative abundances42. Rao quadratic entropy (Q) was used as a measure for functional diversity (FDQ value, Fig. 2)43,44. FDQ is unit-less and shows how distinct communities are functionally different, i.e. increasing Rao’s Q values indicate functionally increasingly different communities. FDQ was calculated using the synthetic functional traits (i.e. species coordinates on Principal Coordinate Analysis (PCoA) axis) defined above and the relative abundance of species44. To calculate synthetic functional traits, we created a functional distance matrix by applying Gower’s distance on each pair of species described by their traits, and then computed a PCoA on this basis45. Then, the first three axes of the PCoA (more than 80% of variance) were selected as synthetic functional traits summarising TA functional space, and species coordinates in the three-dimensional space defined by the PCoA were used to calculate FDQ.
To see if functional diversity of TA communities responds similarly to disturbances in both datasets, redundancy analysis (RDA) was performed using six functional traits of the TA community and DWT, macroscopic charcoal influx and pollen indicators linked to human activities, i.e. released by ruderal plants as explanatory variables (Fig. 3, Table 2). We later used structural equation modelling (SEM)46 to test the effect of different disturbances on the linkages among TA community structure, functional trait composition and functional diversity, and on local vegetation composition (plant macro-remains) (Fig. 4). Using a priori knowledge, we developed several hypothetical mechanisms to build a network of causal relationships including predicted effects of disturbances46 (Table 1). We hypothesised within this causal network that environmental disturbances influence the TA community structure both directly and indirectly through its effect on local vegetation composition. We assumed that TA community structure drives its trait composition, and thus its functional diversity (FDQ). We used pollen human indicators as a proxy for peat extraction and macroscopic charcoal influx as a proxy for fire. For each core, we ran a DCA on a TA data set and extracted DCA site scores as a proxy for TA community composition. Similarly, we ran a PCA on CWM traits of TA of each core and extracted PCA site scores as a proxy for trait community composition. This model was tested for fire (n = 146) and human (n = 35) disturbances, respectively. Adequate model fits are indicated by non-significant χ2 tests (P > 0.05), a low Standardised Root Mean Square Residual index (SRMR < 0.1), and a low Root Mean Square Error of Approximation index (RMSEA < 0.1). Detailed pollen data used for both profiles can be found in Kołaczek, et al.36 and Marcisz, et al.13. All numerical analyses were conducted with R 3.0.147 using the packages FD42, sem48 and vegan49.

The models are significant (P = 0.001 for each model, see Table 2). Black points represent individual samples. Green circles indicate samples from pre-disturbance communities; yellow circles indicate samples from post-disturbance communities. Abbreviations used in the figure: Mixo – mixotrophs; BodSiz – body size; BioVol – biovolume; ApeSize – diameter of the shell aperture; ApePos – shell aperture position; BodRange – body range; DWT – reconstructed depth to water table; macroCHAR – macroscopic charcoal influx/accumulation rate; ruderals – pollen human indicators.

Solid lines are significant paths (P < 0.05), while dashed lines represent non-significant paths (P > 0.05). Squared multiple correlations (R2) for the predicted/dependent factors are given on the top of the box of the dependent variable. TA comp = testate amoebae species composition (PCA axis 1); Macro comp = plant macro-remains composition (PCA axis 1); Traits comp = testate amoebae trait composition (PCoA axis 1). These threes variables are synthetic variables (see methods). Models include macroscopic charcoal influx (proxy for local fire) and ruderals (proxy for direct human impact) used as disturbance indicators for fire and direct human impact, respectively.
Results
Diversity indices of the testate amoeba functional groups
The compositional change of TA showed a drastic turnover in both peatlands in response to disturbances; however, this occurred with a delay (Fig. 1). In Linje, the decrease in compositional change started in cal. AD 1390, after local fires appeared on the mire and the values decreased steadily until cal. AD 1700. Disturbed layer in PK also revealed decrease in TA community richness. It should be mentioned that communities in the disturbed layer might have partly been a result of secondary colonisation by TA (mainly dry indicators) after the peat disturbance. Post-disturbance TA communities stabilised in Linje. TA community composition in both peatlands remained quite stable and no community resilience can be observed. Evenness and richness showed parallel trends. In Linje they rose to cal. AD 400, when a short dry shift was recorded. The lowest values were recorded at cal. AD 620, and these rose again along with water table increases. During the fires, evenness and richness reached the highest values at cal. AD 1390, and then decreased. After the period of increased fire activity (from cal. AD 1700), the values were slightly variable, with a general rising trend. A similar situation was observed in PK, where evenness and richness were consistent along the core reflecting minima in disturbed layer. Post-disturbance (since cal. AD 1900) evenness and richness values were low; they started rising when the peatland was regenerating from disturbances (cal. AD 1965).
Compared to the community response, the response of some groups was delayed. The change of shell size in response to disturbance is clearly visible in Linje, where at the beginning of local fire activity richness and evenness of small species increased (cal. AD 1390). Simultaneously, evenness and richness of large TA slightly declined. In general, richness and evenness of large TA followed the community trend, whereas in the group of small TA evenness and richness varied before the fire, and after the fire they show similar tendencies. In PK, evenness and richness of large TA followed the community trend up to the disturbed layer; together with disturbance and post-disturbance evenness and richness of small TA was in line with the community response.
Species with acrostomic apertures were the most abundant in both data sets. In Linje, evenness and richness were very similar to community trends; during fires values decreased, and increased post-disturbance. Similarly, in PK richness and evenness followed community trends. In both peatlands, TA possessing axial apertures were highly variable. In pre-disturbance communities, TA with plagiostomic aperture were either not present in the community (PK), or their abundances were low (Linje).
In Linje, evenness and richness of TA preferring dry and intermediate water tables were variable pre-disturbance (up to cal. AD 1390), but during local fires indices values were more stable and began to change post fire. Evenness and richness of wet indicators were more stable post-disturbance, having the biggest variations during and after disturbances. In PK, the highest pre-disturbance (cal. AD 0–150) richness and evenness were observed for TA preferring intermediate and low water tables. Evenness and richness of all groups (especially dry TA) decrease in the layer disturbed by peat extraction.
Compositional change and changes in evenness and richness of TA preferring lower pH were consistent with the general community trends in both peatlands. Species preferring higher pH were less abundant and in PK compositional change was observed in the post-disturbance period. In Linje, the group of TA preferring high pH shifted during disturbances and stabilised post-disturbances.
Functional traits of the testate amoebae and community patterns
The analysis of TA functional traits revealed similarities in community response to human induced disturbances: fire and peat extraction. In both datasets, FDQ values decreased after disturbances started (stronger decreases are noted for PK), indicating that TA communities were functionally more similar during disturbances. In general, this shift was mainly caused by a decline in mixotrophs and body size (Fig. 2). Both peatlands were in stable hydrological conditions in the past (until cal. AD 1390 in Linje and cal. AD 650 in PK) and during that time TA traits were only slightly variable. In Linje mire, a slight decrease in water table in cal. AD 400 was simultaneous with a drop in mixotrophs and body range increase. At the beginning of fire activity in Linje mire (cal. AD 1225) and at the bottom of the disturbed layer in PK, trait values changed. In particular, we found a sharp decrease in mixotrophs in both mires, although this was more pronounced in Linje. Simultaneously, shell size and aperture size dropped. However, the biovolume remained quite stable. In PK, the number of mixotrophs decreased together with body size, aperture size and biovolume at the bottom of the disturbed layer. The response of TA communities in PK was not as rapid as in Linje; however, it is not possible to quantify this precisely, as the disturbed layers possess a disturbed chronology and they were not taken into account for depth-age modelling37. In both peatlands in the disturbed layer, large taxa and mixotrophs were eliminated from the community, while the number of small taxa increased. Post-disturbances in both peatlands, body size and aperture size increased, together with a slight increase in biovolume.
We used RDA ordination to explore the relationships between TA traits, DWT, ruderals and macroscopic charcoal influx, to define possible similarities in TA trait composition in peatlands that have been under different stressors. RDA models are highly significant (Fig. 3, Table 2), and they show comparable patterns of TA trait composition in both profiles, connected to hydrological (dry-to-wet) gradient. Mixotrophs are connected to pre-disturbance communities and high water table (Linje and PK), and negatively connected to macroscopic charcoal (Linje).
SEMs revealed that functional diversity of TA was significantly influenced by both fire and human disturbance (peat extraction) (Fig. 4). The effect of disturbance factors on TA is, however, different for both datasets. Fire disturbance explained only 10% (R2 = 0.1) of TA functional diversity, while human disturbances explained 52% (R2 = 0.52) of TA functional diversity (Fig. 4). In Linje, SEMs indicate that fire indirectly influenced TA functional diversity through its effect on vegetation composition (path = 0.53), which was then positively linked to TA community composition (0.68). TA community composition then significantly influenced the CWM trait composition (0.50) and functional diversity of TA (−0.31). In PK, human disturbances negatively correlated to macrofossil composition (−0.63) and positively to TA composition (0.35). The model also showed that plant macrofossil composition was a driver of TA composition (−0.48). Shifts in TA composition then strongly drove TA trait composition (0.83) and functional diversity (−0.72).
Discussion
The analysis of TA functional traits revealed similar tendencies in both peatlands (Fig. 2), comparable with changes observed in the compositional change, evenness and richness indices along the studied datasets (Fig. 1). We found the largest variations of traits and indices together with or after human disturbance. Compositional change of TA community was recorded in both peatlands when disturbance started and TA communities were functionally more similar under the influence of stressors (Figs 1 and 2). Post-disturbances, TA communities remained stabilised. Compared to community patterns, we observed a delay in the response of some functional groups of TA (e.g. large TA or TA preferring pH > 4 in Linje, in case of which compositional change was observed when fires terminated, Fig. 1). However, in disturbed layers, the drop in the water table is simultaneous with a decrease in body size and aperture size, with no significant variations in biovolume (Fig. 2).
Analysis of TA functional groups revealed that richness and abundances of small TA and species preferring dry conditions were higher just after the influence of local fire, but decreases in the abundance of large TA and those preferring wet conditions were recorded with a short delay. This delay in response of wet indicators was also recorded in the cross-correlation analysis, i.e. of macroscopic charcoal influx with wet indicator species Heleopera petricola13, and led to the coexistence of wet and dry communities at the beginning of the local fire activity period. However, a clear coexistence of wet and dry communities was not recorded in PK, where the most abundant taxa were TA preferring intermediate water tables. This suggests lower water table seasonal variability and gradual water table lowering in the disturbed layer or, possibly, a combination of pre-disturbance communities established in this layer and those that colonised after the disturbance. In wet conditions, the co-occurrence of small and large TA is possible because they occupy different trophic niches with different functions in the microbial food web. Large TA are usually top predators feeding mostly on rotifers, nematodes, ciliates or other (smaller) TA, whereas small TA feed on small organisms that can be consumed through small apertures, mostly algae, fungi and bacteria32,50. In PK, high numbers of small taxa were recorded in the peat layers, where increased frequencies of fungi were recorded (personal communication with M. Karpińska-Kołaczek, May 2016). The coexistence of small and large TA taxa was already recorded in observational and experimental studies, and this was linked to peatland micro-topography34,39,51 and thus different habitats. TA preferring dry conditions mostly live on elevated hummocks, whereas TA preferring wet conditions live in pools. Fire on the peatland surface results in the removal of dry hummocks, whereas fire in the surrounding forest leads to deforestation, runoff changes and temporal peatland surface flooding52, as well as to increased dust deposition on the peatland surface35. Peatland flooding after fire is suitable for TA preferring wet conditions, but TA preferring dry conditions can still survive temporal increases in wetness. After the runoff changes and water loss is observed on the surface of the mire, microhabitats suitable for TA preferring wet conditions rapidly shrink, whereas those appropriate for TA preferring dry conditions are enlarged.
During the post-disturbance period in PK, large TA reappeared in low abundances and their evenness and richness decreased. In Linje, however, large TA got richer in the final stage of disturbances, but when fire activity decreased their evenness and richness decreased again. When disturbances were terminated, larger TA were not as abundant as before the disturbances and failed to dominate the community. This may be due to mire size, as small mires (e.g. Linje) possess low levels of variety and numbers of microhabitats. Therefore, when TA species become extinct, individuals from other habitats cannot colonise quickly. In contrast, in larger mires with variable microhabitats present and bigger and more diverse species pool for potential immigrants, such as PK, the possibility for faster colonisation from other microhabitats present on the peatland surface is higher. Another important factor is species size. Large species are heavier than smaller ones53,54; therefore, it is unlikely that they will colonise suitable habitats quickly. Wanner and Xylander55 observed that small Euglyphid TA were present in all succession stages, whereas large taxa occurred in later succession stages. In Sphagnum peatlands the most common Euglyphids are Euglypha sp., Assulina sp., Trinema sp. and Corythion sp.56. Tests of Euglypha sp. are built of easily dissolvable silica plates56; therefore, in palaeoecological reconstructions they are most often found only in the recent top peat layers52. Small Trinema sp. and Corythion sp. taxa were present in Linje and PK TA communities during and after disturbances, which suggests that small species do indeed colonise available niches rapidly and that they are not discouraged by tough environmental conditions.
Structural equation modelling provides a synthesis of the relationships between different proxies33. Using SEM, Lamentowicz et al.33 revealed that TA aperture position closely reflects water table variations. Similarly, we show that TA trait composition is in general affected by disturbances; however, this was more evident when peat extraction appeared (Fig. 4). In the case of fire, the composition of plant macrofossils on the mire surface was influenced first – a decrease in the abundance of Sphagnum and increase in abundance of Eriophorum vaginatum was recorded, and later TA were affected (see Figs 3 and 4 in Marcisz et al.13). Exploration of TA traits enabled us to confirm the previous findings of Fournier et al.34, who recorded that drying caused a decrease in species size and biovolume, and of Laggoun-Défarge et al.53, who found that larger species were more abundant in unexploited (undisturbed) sites on peatland than in cut-over parts.
Moreover, our findings show that mixotrophy and plagiostomic aperture traits were connected with local disturbances and, hence, we suggest that these traits could be considered disturbance indicators in palaeoecology. Mixotrophy was connected with pre-disturbance communities, where in periods of high water tables high numbers of mixotrophs (mainly Hyalosphenia papilio, Archerella flavum, Heleopera sphagni and Amphitrema wrightianum) were recorded in both peatlands (Fig. 3). Mixotrophs were lost during disturbances and they dropped rapidly in Linje. This underlines how sensitive mixotrophs are to environmental stressors including anthropogenically-induced drying39, and this is in accordance with previous research that proved their sensitivity to dust deposition35,57. Rapid drops in the number of dominant mixotrophs (H. papilio, A. flavum and A. wrightianum) in Linje can be connected not only with water table lowering, but also with dust deposition during local fires (see Figs 3 and 4 in Marcisz et al.13). Both factors may have led to a limiting of resources for mixotrophs which, consequently, were eliminated from the community. Moreover, mixotrophs are also influenced by light intensity, and light limitation causes a drop in their abundance39,58. The shade can be of different origin, it can be connected with air circulation, atmospheric dust deposition, more intensive growth of dwarf shrubs (such as Ledum palustre) in dry periods and/or local afforestation59. Secondary forest succession usually occurs along with water table decreases in peatlands. As it was lately shown, mixotrophs play a crucial role in C fixation and their functioning may be important for peatlands60. Therefore more studies on mixotrophs are needed, principally to separate the effect of light from the influence of drought.
Plagiostomic apertures were clearly connected with disturbances and the post-disturbance period (Fig. 1). Small species possessing plagiostomic apertures (Trinema sp. and Corythion sp.) were present in very low numbers during the short drought episode cal. AD 700 in Linje and were not present in PK before disturbances. They became more abundant when disturbances occurred and were established in TA communities post-disturbance. Plagiostomic apertures, small and hidden in the shell side56, are an adaptation trait for surviving tough conditions, e.g. periods of rapid water losses61, very low water tables, deforestation and mineral matter input57 or increased dust deposition on the peatland surface13,35. This is in line with the findings of Lamentowicz et al.33, who showed a link between the aperture type and TA adaptation to hydrological conditions. Similarly, in an experimental study in the tropical montane rainforest, Krashevska et al.58 observed that TA species with less protected terminally located apertures were significantly affected by lower water content.
Conclusions
Under global change conditions, ecosystems are exposed to drastic shifts; hence, recognition of useful indicators of disturbances might be essential to assess the state of ecosystem functioning. We revealed that testate amoebae morphological traits were linked to environmental conditions over a long temporal perspective: (1) testate amoebae preferring dry conditions tended to prefer lower pH, possessed small shells and hidden plagiostomic apertures which are an adaptation to low water content, whereas (2) testate amoebae preferring wet conditions tended to prefer higher pH and possessed larger shells with acrostomic aperture. We underline the sensitivity of mixotrophs to abrupt disturbances. We show that species possessing plagiostomic apertures were indicators of disturbances and that their appearance during dry phases allowed their constant presence in post-disturbance communities. Moreover, disturbances caused substantial body size decreases. The above-mentioned changes in TA communities show that human impact and fires had long-term effects on peatland communities. Our results suggest a low ecosystem resilience of peatlands that – once disturbed – may not be self-restored rapidly within a few decades. Changes in trait composition affected by fire and/or peat extraction are seen in post-disturbance communities, i.e. through the establishment of new traits (plagiostomic aperture) or a lack of previously abundant traits (mixotrophy).
Our results show that testate amoebae functional traits may serve as a valuable proxy that supports conventional reconstructions of the past environmental changes in peatlands, and further analysis of trait-ecosystem relationships could make them valuable indicators of the contemporary ecosystem state. Moreover, the trait approach for testate amoebae requires more attention and more studies focused on the complex evolutionary links between testate amoebae morphology and ecosystem function.
Additional Information
How to cite this article: Marcisz, K. et al. A novel testate amoebae trait-based approach to infer environmental disturbance in Sphagnum peatlands. Sci. Rep. 6, 33907; doi: 10.1038/srep33907 (2016).
References
Sutherland, W. J. et al. Identification of 100 fundamental ecological questions. Journal of Ecology 101, 58–67, doi: 10.1111/1365-2745.12025 (2013).
Seddon, A. W. R. et al. Looking forward through the past: identification of 50 priority research questions in palaeoecology. Journal of Ecology 102, 256–267, doi: 10.1111/1365-2745.12195 (2014).
IPCC. IPCC, 2013: Climate Change 2013. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 1535 (Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2013).
Morris, J. L., McLauchlan, K. K. & Higuera, P. E. Sensitivity and complacency of sedimentary biogeochemical records to climate-mediated forest disturbances. Earth-Science Reviews 148, 121–133 (2015).
Colombaroli, D. et al. Response of broadleaved evergreen Mediterranean forest vegetation to fire disturbance during the Holocene: insights from the peri-Adriatic region. Journal of Biogeography 36, 314–326 (2009).
Dieleman, C. M., Branfireun, B. A., McLaughlin, J. & Lindo, Z. Climate change drives a shift in peatland ecosystem plant community: Implications for ecosystem function and stability. Glob Chang Biol 21, 388–395, doi: 10.1111/gcb.12643 (2015).
Seddon, A. W. R., Froyd, C. A., Leng, M. J., Milne, G. A. & Willis, K. J. Ecosystem Resilience and Threshold Response in the Galapagos Coastal Zone. PloS one 6, doi: 10.1371/journal.pone.0022376 (2011).
Marchetto, A., Colombaroli, D. & Tinner, W. Diatom response to mid-Holocene climate change in Lago di Massaciuccoli (Tuscany, Italy). Journal Of Paleolimnology 40, 235–245, doi: 10.1007/s10933-007-9154-6 (2008).
Parish, F. et al. Assessment on peatlands, biodiversity and climate change: main report. Global Environment Centre, Kuala Lumpur and Wetlands International, Wageningen 179 pp (2008).
Sherwood, J. H. et al. Effect of drainage and wildfire on peat hydrophysical properties. Hydrol. Process. 27, 1886–1874, doi: 10.1002/hyp.9820 (2013).
Fenner, N. & Freeman, C. Drought-induced carbon loss in peatlands. Nature Geoscience 4, 895–900, doi: 10.1038/ngeo1323 (2011).
Turetsky, M. R., Donahue, W. F. & Benscoter, B. W. Experimental drying intensifies burning and carbon losses in a northern peatland. Nat Commun 2, 514 (2011).
Marcisz, K. et al. Long-term hydrological dynamics and fire history over the last 2000 years in CE Europe reconstructed from a high-resolution peat archive. Quaternary Science Reviews 112, 138–152, doi: 10.1016/j.quascirev.2015.01.019 (2015).
Turner, T. E., Swindles, G. T. & Roucoux, K. H. Late Holocene ecohydrological and carbon dynamics of a UK raised bog: impact of human activity and climate change. Quaternary Science Reviews 84, 65–85, doi: 10.1016/j.quascirev.2013.10.030 (2014).
Robroek, B. J. M. et al. Peatland vascular plant functional types affect methane dynamics by altering microbial community structure. Journal of Ecology 103, 925–934, doi: 10.1111/1365-2745.12413 (2015).
Kettridge, N. et al. Moderate drop in water table increases peatland vulnerability to post-fire regime shift. Scientific Reports 5, 8063, doi: 10.1038/srep08063 (2015).
Krebs, P., Pezzatti, G. B., Mazzoleni, S., Talbot, L. M. & Conedera, M. Fire regime: history and definition of a key concept in disturbance ecology. Theory Biosci. 129, 53–69, doi: 10.1007/s12064-010-0082-z (2010).
Turetsky, M. R. et al. Global vulnerability of peatlands to fire and carbon loss. Nature Geosci 8, 11–14, doi: 10.1038/ngeo2325 (2015).
Soja, A. J. et al. Climate induced boreal forest change: predictions versus current observations. Glob. Planet. Chang. 56, 274–296 (2007).
Cadotte, M. W., Arnillas, C. A., Livingstone, S. W. & Yasui, S. E. Predicting communities from functional traits. Trends Ecol Evol. 30, 510–511 (2015).
Cadotte, M. W., Carscadden, K. & Mirotchnick, N. Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology 48, 1079–1087, doi: 10.1111/j.1365-2664.2011.02048.x (2011).
de Vries, F. T. et al. Abiotic drivers and plant traits explain landscape-scale patterns in soil microbial communities. Ecology Letters 15, 1230–1239, doi: 10.1111/j.1461-0248.2012.01844.x (2012).
Vandewalle, M. et al. Functional traits as indicators of biodiversity response to land use changes across ecosystems and organisms. Biodiversity and Conservation 19, 2921–2947, doi: 10.1007/s10531-010-9798-9 (2010).
Díaz, S. & Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution 16, 646–655, doi: 10.1016/S0169-5347(01)02283-2 (2001).
Jackson, S. T. & Blois, J. L. Community ecology in a changing environment: Perspectives from the Quaternary. Proceedings of the National Academy of Sciences of the United States of America 112, 4915–4921, doi: 10.1073/pnas.1403664111 (2015).
Jassey, V. E. J. et al. Above- and belowground linkages in Sphagnum peatland: climate warming affects plant-microbial interactions. Global Change Biol 19, 811–823 (2013).
Wilkinson, D. A. & Mitchell, E. A. D. Testate amoebae and nutrient cycling with particular reference to soils. Geomicrobiology Journal 27, 520–533 (2010).
Mitchell, E. A. D., Charman, D. J. & Warner, B. G. Testate amoebae analysis in ecological and paleoecological studies of wetlands: past, present and future. Biodiversity and Conservation 17, 2115–2137 (2008).
Payne, R. J. et al. Significance testing testate amoeba water table reconstructions. Quaternary Sci. Rev. 138, 131–135, doi: 10.1016/j.quascirev.2016.01.030 (2016).
Mitchell, E. A. D., Lamentowicz, M., Payne, R. J. & Mazei, Y. Effect of taxonomic resolution on ecological and palaeoecological inference - a test using testate amoeba water table depth transfer functions. Quaternary Sci Rev 91, 62–69, doi: 10.1016/j.quascirev.2014.03.006 (2014).
Jassey, V. E. J. et al. Plant functional diversity drives niche-size-structure of dominant microbial consumers along a poor to extremely rich fen gradient. Journal of Ecology 102, 1150–1162 (2014).
Jassey, V. E. J., Shimano, S., Dupuy, C., Toussaint, M. L. & Gilbert, D. Characterizing the Feeding Habits of the Testate Amoebae Hyalosphenia papilio and Nebela tincta along a Narrow “Fen-Bog” Gradient Using Digestive Vacuole Content and 13C and 15N Isotopic Analyses. Protist 163, 451–464 (2012).
Lamentowicz, M. et al. Climate change over the last 4000 years in a Baltic bog in northern Poland revealed by a trait-based approach, biotic proxies, and stable isotopes. Palaeogeography Palaeoclimatology Palaeoecology 418, 261–277 (2015).
Fournier, B., Lara, E., Jassey, V. E. J. & Mitchell, E. A. D. Functional traits as a new approach for interpreting testate amoeba palaeo-records in peatlands and assessing the causes and consequences of past changes in species composition. The Holocene 25, 1375–1383, doi: 10.1177/0959683615585842 (2015).
Fiałkiewicz-Kozieł, B. et al. Peatland microbial communities as indicators of the extreme atmospheric dust deposition. Water Air and Soil Pollution 226, 97 (2015).
Kołaczek, P., Fiałkiewicz-Kozieł, B., Karpińska-Kołaczek, M. & Gałka, M. The last two millennia of vegetation development and human activity in the Orawa-Nowy Targ Basin (south-eastern Poland). Acta Palaeobotanica 50, 133–148 (2010).
Fiałkiewicz-Kozieł, B., Kołaczek, P., Michczyński, A. & Piotrowska, N. The construction of a reliable absolute chronology for the last two millennia in an anthropogenically disturbed peat bog: Limitations and advantages of using a radio-isotopic proxy and age-depth modelling. Quaternary Geochronology 25, 83–95 (2015).
Słowińska, S., Słowiński, M. & Lamentowicz, M. Relationships between Local Climate and Hydrology in Sphagnum Mire: Implications for Palaeohydrological Studies and Ecosystem Management. Polish Journal of Environmental Studies 19, 779–787 (2010).
Marcisz, K. et al. Seasonal changes in Sphagnum peatland testate amoeba communities along a hydrological gradient. European Journal of Protistology 50, 445–455, doi: 10.1016/j.ejop.2014.07.001 (2014).
Łajczak, A. Torfowiska Kotliny OrawskoNowotarskiej. Rozwój, antropogeniczna degradacja, renaturyzacja i wybrane problemy ochrony (W. Szafer Institute of Botany Polish Academy of Sciences, Kraków, 2006).
Colombaroli, D., Beckmann, M., Van der Knaap, W. O., Curdy, P. & Tinner, W. Changes in biodiversity and vegetation composition in the central Swiss Alps during the transition from pristine forest to first farming. Diversity Distrib. 19, 157–170, doi: 10.1111/j.1472-4642.2012.00930.x (2013).
FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0–12 (2014).
Ricotta, C. A note on functional diversity measures. Basic and Applied Ecology 6, 479–486 (2005).
Pillar, V. D. et al. Functional redundancy and stability in plant communities. Journal of Vegetation Science 24, 963–974, doi: 10.1111/jvs.12047 (2013).
Villeger, S., Mason, N. W. H. & Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89, 2290–2301 (2008).
Grace, J. B., Adler, P. B., Harpole, W. S., Borer, E. T. & Seabloom, E. W. Causal networks clarify productivity-richness interrelations, bivariate plots do not. Funct. Ecol. 28, 787–798 (2014).
R: A Language and Environment for Statistical Computing. (Vienna: R Foundation for Statistical Computing, 2011).
Grace, J. B., Anderson, T. M., Olff, H. & Scheiner, S. M. On the specification of structural equation models for ecological systems. Ecological Monographs 80, 67–87 (2010).
Oksanen, J. et al. vegan: Community Ecology Package (2010).
Gilbert, D., Amblard, C., Bourdier, G., Francez, A.-J. & Mitchell, E. A. D. Le régime alimentaire des thécamoebiens. L’Année Biologique 39, 57–68 (2000).
Tsyganov, A. N., Aerts, R., Nijs, I., Cornelissen, J. H. C. & Beyens, L. Sphagnum-dwelling Testate Amoebae in Subarctic Bogs are More Sensitive to Soil Warming in the Growing Season than in Winter: the Results of Eight-year Field Climate Manipulations. Protist 163, 400–414 (2012).
Sillasoo, Ü. et al. Peat multi-proxy data from Männikjärve bog as indicators of late Holocene climate changes in Estonia. Boreas 36, 20–37 (2007).
Laggoun-Défarge, F. et al. Cut-over peatland regeneration assessment using organic matter and microbial indicators (bacteria and testate amoebae). Journal of Applied Ecology 45, 716–727, doi: 10.1111/j.1365-2664.2007.01436.x (2008).
Wilkinson, D. M., Koumoutsaris, S., Mitchell, E. A. D. & Bey, I. Modelling the effect of size on the aerial dispersal of microorganisms. Journal of Biogeography 39, 89–97 (2012).
Wanner, M. & Xylander, W. E. R. Biodiversity development of terrestrial testate amoebae: is there any succession at all? Biol. Fertil. Soils 41, 428–438 (2005).
Mazei, Y. & Tsyganov, A. N. Freshwater testate amoebae (KMK, 2006).
Lamentowicz, M. et al. Multiproxy study of anthropogenic and climatic changes in the last two millennia from a small mire in central Poland. Hydrobiologia 631, 213–230, doi: 10.1007/s10750-009-9812-y (2009).
Krashevska, V., Sandmann, D., Maraun, M. & Scheu, S. Consequences of exclusion of precipitation on microorganisms and microbial consumers in montane tropical rainforests. Oecologia 170, 1067–1076, doi: 10.1007/s00442-012-2360-6 (2012).
Payne, R. J. et al. Tree encroachment may lead to functionally-significant changes in peatland testate amoeba communities. Soil Biology & Biochemistry 98, 18–21, doi: 10.1016/j.soilbio.2016.04.002 (2016).
Jassey, V. E. J. et al. An unexpected role for mixotrophs in the response of peatland carbon cycling to climate warming. Scientific Reports 5, 16931, doi: 10.1038/srep16931 (2015).
Chardez, D. & Lambert, J. Thécamoebiens indicateurs biologiques (Protozoa Rhizopoda testacea). Bulletin des recherches agronomiques de Gembloux 16, 181–204 (1981).
Poska, A., Saarse, L. & Veski, S. Reflections of pre- and early-agrarian human impact in the pollen diagrams of Estonia. Palaeogeography Palaeoclimatology Palaeoecology 209, 37–50 (2004).
Acknowledgements
This research was supported by grant PSPB-013/2010 from Switzerland through the Swiss Contribution to the enlarged European Union (CLIMPEAT, www.climpeat.pl). We acknowledge the support of the Scientific Exchange Programme from the Swiss Contribution to the New Member States of the European Union (Sciex-NMSch) – SCIEX Scholarship Fund, project RE-FIRE 12.286. The research was supported by the National Science Centre (Poland) – grants 2015/17/B/ST10/01656 and UMO-2014/13/B/ST10/02091, and a subsidy for young researcher from the Faculty of Geographical and Geological Sciences, Adam Mickiewicz University in Poznań – 0800000000/505/060/STAT_MN (funds for K. Marcisz, 2016). This study is a contribution to the Interdisciplinary Environmental Life Sciences – PhD Programme: EU Human Capital Programme “AMU: A Unique Graduate = Possibilities. Improving the didactic potential of Adam Mickiewicz University through innovative education in English, interdisciplinary approaches, e-learning, and investments in staff”, a project co-funded by the European Union within the European Social Fund POKL.04.01.01–00–019/10 528.81.206 poz 310.
Author information
Authors and Affiliations
Contributions
Testate amoebae were analysed by K.M. with the assistance of M.L. Charcoal was analysed by K.M. with the assistance of W.T. and D.C. Pollen was analysed by P.K. (Linje and Puścizna Krauszowska) and M.K.-K. (Puścizna Krauszowska), plant macrofossils by M.S. (Linje) and M.G. (Puścizna Krauszowska). Age-depth models (Suppl. material) were prepared by P.K. Statistical analysis of functional traits were performed by K.M. with the help and assistance of D.C., V.E.J.J. and M.L. Structural equation modelling was done by V.E.J.J. Figures and tables were prepared by K.M. and V.E.J.J. (Figure 4, SEM). K.M. interpreted the data with the assistance of M.L., D.C., V.E.J.J. and W.T. K.M. wrote the paper to which all authors contributed with discussions and/or text.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Marcisz, K., Colombaroli, D., Jassey, V. et al. A novel testate amoebae trait-based approach to infer environmental disturbance in Sphagnum peatlands. Sci Rep 6, 33907 (2016). https://doi.org/10.1038/srep33907
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep33907
This article is cited by
-
A review of functional approaches for the study of freshwater communities with a focus on zooplankton
Hydrobiologia (2023)
-
Small-scale Variation of Testate Amoeba Assemblages: the Effect of Site Heterogeneity and Empty Shell Inclusion
Microbial Ecology (2019)
-
Testate amoebae community analysis as a tool to assess biological impacts of peatland use
Wetlands Ecology and Management (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
