
Abstract
Megaherbivores (>1000 kg) are critical for ecosystem health and function, but face population collapse and extinction globally. The future of these megaherbivore-impoverished ecosystems is difficult to predict, though many studies have demonstrated increasing representation of C3 woody plants. These studies rely on direct observational data, however, and tools for assessing decadal-scale changes in African ecology without observation are lacking. We use isotopic records of historical common hippopotamus (Hippopotamus amphibius) canines to quantify herbaceous vegetation change in Queen Elizabeth National Park, Uganda following a period of civil unrest and poaching. This poaching event led to population collapse of two threatened African megaherbivore species: hippopotamus and African elephants (Loxodonta africana). Serial carbon isotope ratios (δ13C) in canine enamel from individuals that lived between 1960–2000 indicated substantial increases in C3 herbaceous plants in their diet (<20% C3 in the 1960s to 30–45% C3 in the 80s and 90s), supported by other observational and ecological data. These data indicate megaherbivore loss results in succession of both woody and herbaceous C3 vegetation and further reaching effects, such as decreased grazing capacity and herbivore biodiversity in the area. Given multiple lines of evidence, these individuals appear to accurately capture herbaceous vegetation change in Mweya.
Similar content being viewed by others

Introduction
Megaherbivores exert a profound influence on ecosystem function and structure across ecosystems globally. They provide critical ecosystem services, such as nutrient cycling, soil transformation, increasing net primary productivity, maintenance of open environments and modification of trophic guild structure1,2. Yet, megaherbivores are threatened with extinction due to the effects of habitat loss, range contraction, competition with livestock, climate change, overharvesting and civil unrest3.
In African ecosystems, important threatened megaherbivores include the common hippopotamus (Hippopotamus amphibius) and African elephants (Loxodonta africana). Hippopotamus significantly engineer vegetation and environments near their aquatic habitats by feeding on ground-layer vegetation, transforming grassy areas into short-grass grazing lawns, and building water channels through areas near their habitat3. Hippopotamus feeding habits are unique in that they swing their heads back and forth close the ground, cropping vegetation closely with their lips; this method of feeding precludes them from consuming aquatic or woody vegetation in large numbers4. Likewise, elephants are important influencers of overall savanna habitat structure, function and diversity at multiple trophic levels5,6,7. Elephants browse selectively, and high elephant populations can lead to woodland decline and can result in the extirpation of woody plants in areas of intense elephant browsing pressure5.
These megaherbivores are critical to the health and maintenance of African savanna ecosystems. Savannas can be described as “mixed tree-grass systems characterized by a discontinuous tree canopy in a conspicuous grass layer”8 that can be defined structurally as having 5–80% fractional woody cover, which includes the structural categories grassland, wooded grassland and woodland/bushland/shrubland9,10,11,12,13. Encroachment of woody plants (“bush encroachment”; C3 shrubs and trees) in the grassy layer (C4 grasses) of savannas is an issue of major concern for ecologists, as increased shrub presence alters soil conditions and suppress grass productivity, affecting organisms at many trophic levels14. Studies on encroaching plants in savannas have focused predominately on C3 woody plants, and little attention has been given to the role of herbaceous non-grassy plants (C3 forbs and herbs). Replacement of grasses by herbaceous plants within savannas can be just as deleterious for ecosystem health and biodiversity as succession of woody vegetation, though the effects of herbaceous plant expansion within grassy biomes have understudied consequences15.
Evaluating the potential short and long-term effects on plant succession within ecosystems after megaherbivore collapse requires historical records of ecology from ecosystems where previous such population collapses have occurred. Comparisons are often drawn between the current rates of extinction and Pleistocene megafaunal extinctions, and though useful, those extinctions resulted in modern ecosystems for which there are no fossil analogs3. Building a historical record of ecology in African savannas to understand megaherbivore collapse and plant succession is difficult, as direct observations over decades have not been made for many savanna ecosystems. We provide a ground-truthed method for reconstructing terrestrial ecology on a decadal scale within African savanna ecosystems using stable carbon isotopes (δ13C) in hippopotamus canine enamel. This paper illuminates long-term ecological effects following population collapse of hippopotamus and elephants in a savanna ecosystem in Queen Elizabeth National Park (QENP), Uganda. A period of civil unrest during the regime of Idi Amin resulted in widespread poaching and near extirpation of elephants from the park during the 1970s. Historical diet records obtained from sequentially-sampled hippopotamus canine enamel for stable carbon isotopes indicate that herbaceous vegetation shifted such that these hippopotamus switched their diets from >80% C4 grass in the 1960s to as low as 55% C4 grass in the 80s and 90s on the Mweya Peninsula, previously one of the most diverse parts of the park. Data that indicates changes in the diets of these individuals accurately reflect both environmental changes and dietary shifts of the population at large, and are supported by other lines of evidence, including gut content studies, vegetation surveys and isotope studies of individuals from discreet time periods16,17,18,19. Though previous studies have used serially sampled hippopotamus enamel to reconstruct environments, none of these studies have had such data placed in the context of recorded environmental change19,20,21. Serial samples of δ13C from hippopotamus canines could serve as a useful measure of ecological change in other savanna parks where hippopotamus are present, and such skeletal collections exist.
The ecological history of Queen Elizabeth National Park
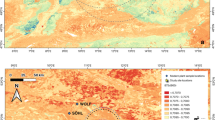
Queen Elizabeth National Park (Fig. 1) is located in western Uganda in the Albertine Rift Valley, along the border of the Democratic Republic of Congo (DRC). The park is 1,979 km2, surrounding Lakes Edward and George. Rainfall varies between 600 and 1400 mm/yr during March-May and in September-November18. The soils in the park are rich in volcanic ash, creating environments with high net primary production and many globally threatened and endemic plant and animal species16,22,23. At the time of census in the mid 1970s, QENP had the densest herbivore biomass (19,928 kg/km−2) on Earth24.

Map of Queen Elizabeth Park, Uganda.
The red circle highlights the Mweya Peninsula. Image created in Illustrator (v. 19.2.0), re-drawn and modified from the image “Uganda_location_map.svg” downloaded from Wikimedia Commons (https://commons.wikimedia.org/wiki/File:Uganda_location_map.svg) and is licensed under the Attribution-ShareAlike 3.0 Germany license. The license terms can be found on the following link: https://creativecommons.org/licenses/by-sa/3.0/de/deed.en.
The effects of three major ecosystem drivers are elephant browsing, hippo grazing, and fire23. In the 1960s and 70s, hippos were observed to feed predominately in grasslands on or near Mweya (i.e., within a few kilometers of Lake Edward)25 that were maintained by elephant browsing and fire5,6,26. The dominant herbaceous vegetation on Mweya during that time was C4 grasses that grow in heavily grazed areas; trees and shrubs occurred in small thickets, clustered around emergent Euphorbia candelabrum25,27.
From 1972 to 1980, during the regime of Idi Amin, management of all national parks essentially ceased, and widespread poaching decimated herbivore populations across the country26,28. Intensification of poaching activities occurred in the late 1970s, resulting in collapsing herbivore populations and genetic bottlenecks28,29. The population of elephants within the park fell from 4,139 to ~150 individuals, and hippo populations fell to ~4,000 from nearly 12,000 individuals28.
Heavy wildlife poaching continued into the mid 1980s28, by which time there was a significant expansion in the size and number of woody vegetation, predominantly Euphorbia candelabrum and Turraea robusta6,17. Hippopotamus range restriction from poaching resulted in a positive-feedback for thicket encroachment: repeated trips onto land led to soil compaction, shunting rainwater into thickets and reducing fire fuel, further suppressing grass regrowth25,30,31.
Stable Isotope ecology in African Mammals
Stable isotope analysis is a powerful tool for understanding aspects of mammalian herbivore ecology in Africa, and is particularly useful for generating ecological records on temporal and spatial scales that are difficult or impossible to observe19,32,33,34. Stable carbon isotope analysis (δ13C) of herbivore tooth enamel can reveal the relative proportions of C3 vs. C4 in the diets of African herbivores, and the relationship between δ13Cdiet and δ13Cenamel values is well-understood in African ungulates35,36.
A common issue in modern ecological systems is generating continuous, decadal-scale historical records of environmental change. Serial samples of biological materials present a time-series of ecological information that can be useful as archives of environment, as each isotope sample represents an independent measure of diet and ecology for that individual21,34,37,38. Hippopotamus canines are excellent long-term ecological archives, since they are ever-growing and include isotopic input spanning ten or more years of enamel growth36,39,40. Therefore, stable isotope analysis of tusks is ideal for quantifying the nocturnal feeding behavior of hippos and thus ground-layer vegetation, and particularly for generating multi-year to decadal-scale ecological records from deceased animals. It is important to note, however, that these records only represent diet of individuals over a discreet time period, rather than of the population. Given the known behavioral characteristics of hippopotamus feeding behavior, we argue that these data can serve as a reflection of group feeding dynamics. Although hippopotamus travel alone to feed, their feeding areas are restricted and generally feed less than 6 km away from their aquatic habitats, often between 0.5–3 km, and tend to feed in the same areas as other hippopotamus2,25.
Hippopotamus ecology and isotopic indicators of environment
The common hippopotamus is a large bodied herbivore found across Africa that lives in varied environments, ranging from arid savanna to forest4,41,42. They are semi-aquatic, spending the day resting in pools, rivers and lakes, and feeding nocturnally on terrestrial vegetation43. Hippos can significantly restructure grazing areas, mowing tall grasses into short, closely-cropped lawns. Early studies of hippo ecology suggested that hippos selectively feed on C4 grasses16,44. However, stable isotope ratios of carbon (13C/12C) in hippopotamus biological tissues have revealed considerable variability in the consumption of C3 plants (herbs and forbs) and C4 plants (tropical grasses < 3000 m), indicating a range in diets from purely C3-based to purely C4-based19,45,46. These data indicate that hippopotamus are more generalist feeders than previously thought, and dietary isotopic data reflect local herbaceous vegetation. Previous analysis of hippopotamus molars (integrating only a few years of growth) between 1970 and 1998 populations reveals statistically significant differences in diet between these two populations (ANOVA, Bonferroni’s test, P < 0.000147). These molar analyses, however, only reveal snapshots of environmental change (i.e., an integrated measure of diet over the years of formation of the tooth) during two discreet time intervals. A more continuous measure of environmental change, such as those obtained by serial sampling of biological tissues, could better resolve when such changes took place.
Materials and Methods
Lower hippopotamus canines were sampled from individuals who died on the Mweya Peninsula within Queen Elizabeth National Park. Date of death was assigned using recorded death dates (if known) or through radiocarbon dating of tusk enamel39. Calendar years for hippopotamus tusk samples were assigned using two methods. The 1960–1970 tusk was dated using bomb-curve radiocarbon dating39. The death years for the other two tusks were known–1991 and 2000. Using an average hippo lower canine growth rate of 4.4 cm/yr from five wild hippo tusks measured with bomb radiocarbon by Uno (2013), the calendar years were estimated for each sample in the profile assuming a constant growth rate.
Samples of enamel were drilled at intervals along the length of the tusk using a diamond-tipped drill bit and Dremel tool. Enamel powders were treated with 2% H2O2 for 30 minutes to remove organics, then washed 3 times with distilled water. Enamel samples were reacted with >100% phosphoric acid in a common acid bath in a dual-inlet Carboflo carbonate device. Stable isotope ratios (13C/12C and 18O/16O) of resulting CO2 were analyzed on an MAT 252 isotope ratio mass spectrometer, and stable isotope ratios are reported as delta (δ) values relative to the international carbon isotope standard, Vienna Pee Dee Belemnite (VPDB), following the standard permil (‰) notation, where δ13C = (Rsample/Rstandard − 1) × 1000. Enamel isotope values were corrected relative to an internal carbonate standard (Carrara Marble) calibrated to V-PBD and two in-house enamel standards (“MHS” and “MRS”). Dietary designations for hippos are given based on estimated dietary intake of C4 plants (lowland tropical grasses; −12.9‰48) and C3 plants (trees, shrubs, herbs, and forbs; −27.9‰48). This is calculated using the isotope enrichment factor (ε*):

where ε∗ is the isotopic enrichment (*denoting compared materials not at equilibrium) between diet and herbivore tooth enamel (14.1‰)35, using the average isotopic value of modern C3 and C4 plants in eastern and central Africa48.
Results
The 1970s tusk reveals a diet of greater than 80% C4 grass intake from ca. 1960 to 1970 (Table 1, Fig. 2, SI Appendix Table 1). The 1991 hippo profile is significantly more depleted in δ13C than the 1970 profile, indicating a mixed C3/C4 diet (ca. 65% C4) from 1982 to 1991, following collapse of the elephant population in Queen (Fig. 2, SI Appendix Table 1). The third tusk includes the time interval from 1985 to 2000 with an estimated diet ranging from ca. 55 to 70% C4. Both the 1991 and 2000 individuals show local minima in δ13C values for the ca. 1988–1989 interval.

Carbon isotope profiles of the three Queen Elizabth hippo serial samples (1970 tusk in black, 1991 tusk in dark grey, 2000 tusk in light grey) and number of hippos (dotted line) and elephants (solid line) present in the park over time23,56.
The chronolgy of major ecological and political events is listed above the graph.
Hippopotamus canine carbon isotopes reveal a diet shift associated with increased consumption of herbaceous vegetation, which indicates a significant encroachment of C3 herbaceous plants within the Mweya Peninsula. Changes in atmospheric δ13C due to the burning of fossil fuels (the Suess Effect, <1‰) are smaller than changes in carbon isotopes from this dietary shift. In non-quantitative vegetation counts conducted in 1992 and 2009, Plumptre and others (2010) found that Cynodon dactylon (a C4 grass), Commelina diffusa, Commelina africana (C3 herbs), Asystasia gangetica (C3 forb), Cyanotis foecunda (a C3 flowering herb), Achyranthes aspera (C3 herb), Ocimum suaveolens (C3 herb), Oplismenus hirtellus (C4 grass) and two Cyperus species have become dominant, which suggests that hippopotamus have been increasingly relying on these C3 forbs and herbs. Though useful, it should be noted that these vegetation counts are not quantitative, and we present these data here as a validation for using hippopotamus diet change to infer ecological change.
Discussion
Our findings reveal that hippopotamus have more flexible diets than previously thought, and can shift their diets to accommodate increased C3 herbaceous groundcover when preferred grasses are no longer present. Hippopotamus have traditionally been characterized as selective C4-grazers that feed on short grasses and sedges (which comprise 95–99% of their diet), supplemented by forbs2. This dietary classification was established by stomach content analysis by Field (1970), conducted on hippos in QENP during the 1960s, when grasses were abundant. However, modern hippopotamus environments are strikingly different from those of the 1960s, when groundcover vegetation on the Mweya Peninsula consisted predominately of grazing-tolerant C4 grasses25,27. Therefore, carbon isotopic records from serially sampled hippopotamus tusks may provide a much-needed quantitative resource for investigating historical ecological change across tropical grassy biomes in Africa.
The effects of poaching in the park have resulted in ecological restructuring of QENP. Aerial photographs and photomosaic analysis of vegetation types within the park between 1950 and 2006 indicate an increase in woody cover of ~30% across QENP18. Our findings demonstrate that in addition to bush encroachment, elephant poaching results in C3 encroachment in the herbaceous ground layer of savannas. The expansion of non-grassy ground vegetation restricts grazing capacity and decreases herbivore biodiversity and biomass, and these effects are intensified by the loss of hippopotamus-maintained grazing lawns27,49.
The long-term and wide-reaching effects of bush encroachment, such as changes in nutrient composition of soil, and %C and %N depletion in heavily encroached areas50, are further compounded by suppression of C4 grass growth, a critical resource for numerous mesoherbivores in savanna ecosystems. The combined effects of bush encroachment, which has already taken place across Africa14,51,52, and elephant poaching, which is occurring at unprecedented rates53, are likely to further exacerbate habitat degradation and suppress populations of grazing herbivores.
Civil war in eastern Democratic Republic of the Congo (DRC) in 1998 led to heavy poaching in Virunga National Park and other areas near QENP, resulting in herbivore migration to QENP and a partial recovery of herbivore communities, which may explain the delayed later succession of woody plants54. Elephant populations have been increasing in the park since 1990 (from ~500 to almost 3000 in 200523), inhibiting only very recent (i.e., with the last 20 years) woody cover encroachment in the park, but not in the Mweya peninsula23. Although the deleterious effects of human conflict and wildlife poaching as a mechanism for ecosystem destabilization are known, further work is needed on the long-term ecological effects of overharvesting on ground vegetation in savanna ecosystems55.
Conclusions
Megaherbivores are key ecological engineers that affect change at nearly every trophic level in African savannas, consuming and spreading nutrients on land and transforming soil properties and landscapes that increase environmental heterogeneity. Dietary stable isotope data from serially sampled hippopotamus canines from the Mweya Peninsula of QENP provide long-term ecological records that reflect increases in C3-vegetation, not only in woody layers as a result of elephant culling, but also in herbaceous ground layers. These isotopic data are supported by other lines of evidence for increasing C3 plant representation in the area. These data are critical for wildlife management within African savanna parks, as increasing C3 vegetation in both woody and herbaceous layers of savanna parks decreases grazing capacity for grass-feeding herbivores. The outcome of this ecological change is that grazers have been pushed to other parts of the national park, and even outside the park itself, posing serious threats for conservation in Africa. Our data suggest that hippopotamus canine isotopes could be used in savanna parks to build decadal records of ecological change in places where direct observational data are lacking.
Additional Information
How to cite this article: Chritz, K. L. et al. Hippopotamus (H. amphibius) diet change indicates herbaceous plant encroachment following megaherbivore population collapse. Sci. Rep. 6, 32807; doi: 10.1038/srep32807 (2016).
References
Asner, G. P. & Levick, S. R. Landscape-scale effects of herbivores on treefall in African savannas. Ecology Letters 15, 1211–1217 (2012).
Owen-Smith, R. N. Megaherbivores: the influence of very large body size on ecology. (Cambridge, 1992).
Ripple, W. J. et al. Collapse of the world’s largest herbivores. Science Advances 1, e1400103–e1400103 (2015).
Eltringham, S. K. The Hippos: Natural History and Conservation. (Academic Press, 1999).
Dublin, H. T., Sinclair, A. & McGlade, J. Elephants and Fire as Causes of Multiple Stable States in the Serengeti-Mara Woodlands. Journal of Animal Ecology 59, 1147–1164 (1990).
Field, C. R. Elephant ecology in the Queen Elizabeth National Park, Uganda. African Journal of Ecology 9, 99–123 (1971).
Valeix, M., Fritz, H., Sabatier, R. & Murindagomo, F. Elephant-induced structural changes in the vegetation and habitat selection by large herbivores in an African savanna. Biological Conservation 144, 902–912 (2011).
Ratnam, J. et al. When is a ‘forest’ a savanna, and why does it matter? Global Ecology and Biogeography 20, 653–660 (2011).
Cerling, T. E. et al. Woody cover and hominin environments in the past 6 million years. Nature 476, 51–56 (2011).
Sankaran, M. et al. Determinants of woody cover in African savannas. Nature 438, 846–849 (2005).
Murphy, B. & Bowman, D. M. What controls the distribution of tropical forest and savanna? Ecology Letters 7, 748–758 (2012).
Good, S. P. & Caylor, K. K. Climatological determinants of woody cover in Africa. Proceedings of the National Academy of Sciences 108, 4902 (2011).
Lehmann, C. E. R., Bond, W. J., Archibald, S. & Hoffmann, W. A. Deciphering the distribution of the savanna biome. New Phytologist 191, 197–209 (2011).
Roques, K. G., O’connor, T. G. & Watkinson, A. R. Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall and density dependence. Journal of Applied Ecology 38, 268–280 (2001).
Bond, W. J. & Parr, C. L. Beyond the forest edge: Ecology, diversity and conservation of the grassy biomes. Biological Conservation 143, 2395–2404 (2010).
Field, C. R. A study of the feeding habits of the hippopotamus (Hippopotamus amphibius Linn.) in the Queen Elizabeth National Park, Uganda, with some management implications. Zoologica Africana 5, 71–86 (1970).
Lock, J. Vegetation change in Queen Elizabeth National Park, Uganda: 1970–1988. African Journal of Ecology 31, 106–117 (1993).
Plumptre, A. J. et al. The effects of environmental and anthropogenic changes on the savannas of the Queen Elizabeth and Virunga National parks in Long Term changes in Africa’s Rift Valley: impacts on biodiversity and ecosystems (ed. Plumptre, A. J. ) 88–105 (Nova Publishers, 2010).
Cerling, T. E. et al. Stable isotope ecology of the common hippopotamus. Journal of Zoology 276, 204–212 (2008).
Harris, J. M., Cerling, T. E., Leakey, M. G. & Passey, B. H. Stable isotope ecology of fossil hippopotamids from the Lake Turkana Basin of East Africa. Journal of Zoology 275, 323–331 (2008).
Souron, A., Balasse, M. & Boisserie, J. R. Intra-tooth isotopic profiles of canines from extant Hippopotamus amphibius and late Pliocene hippopotamids (Shungura Formation, Ethiopia): insights into the seasonality of diet and climate. Palaeogeography, Palaeoclimatology, Palaeogeography 342–343, 97–110 (2012).
Plumptre, A. J. et al. The biodiversity of the Albertine Rift. Biological Conservation 134, 178–194 (2007).
Plumptre, A. J. et al. The Impact of Fire and Large Mammals on the Ecology of Queen Elizabeth National Park. (Wildlife Conservation Society and Woods Hole Research Centre, 2010).
Coe, M. J., Cumming, D. H. & Phillipson, J. Biomass and production of large African herbivores in relation to rainfall and primary production. Oecologia 22, 341–354 (1976).
Lock, J. M. The effects of hippopotamus grazing on grasslands. The Journal of Ecology 60, 445–467 (1972).
Aleper, D. & Moe, S. R. The African savannah elephant population in Kidepo Valley National Park, Uganda: changes in size and structure from 1967 to 2000. African Journal of Ecology 44, 157–164 (2006).
Strugnell, R. G. & Pigott, C. D. Biomass, shoot-production and grazing of two grasslands in the Rwenzori National Park, Uganda. The Journal of Ecology 66, 73–96 (1978).
Muwanika, V. B. et al. A recent bottleneck in the warthog and elephant populations of Queen Elizabeth National Park, revealed by a comparative study of four mammalian species in Uganda national parks. Animal Conservation 6, 237–245 (2003).
Eltringham, S. K. & Malpas, R. C. The decline in elephant numbers in Rwenzori and Kabalega Falls National Parks, Uganda. African Journal of Ecology 18, 73–86 (1980).
Thornton, D. D. The effect of the complete removal of hippopotamus on grassland in the Queen Elizabeth National Park, Uganda. African Journal of Ecology 9, 47–55 (1971).
Eltringham, S. K. Changes in the large mammal community of Mweya Peninsula, Rwenzori National Park, Uganda, following removal of hippopotamus. Journal of Applied Ecology 11, 855 (1974).
Cerling, T. E., Chritz, K. L., Jablonski, N. G., Leakey, M. G. & Manthi, F. K. Diet of Theropithecus from 4 to 1 Ma in Kenya. Proceedings of the National Academy of Sciences 110, 10507–10512 (2013).
Lee-Thorp, J. A. & Van der Merwe, N. J. Carbon isotope analysis of fossil bone apatite. South African Journal of Science 83, 712–715 (1987).
Blumenthal, S. A., Chritz, K. L., Rothberg, J. & Cerling, T. E. Detecting intraannual dietary variability in wild mountain gorillas by stable isotope analysis of feces. Proceedings of the National Academy of Sciences 109, 21277–21282 (2012).
Cerling, T. E. & Harris, J. M. Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia 120, 347–363 (1999).
Passey, B. H., Cerling, T. E. & Schuster, G. T. Inverse methods for estimating primary input signals from time-averaged isotope profiles. Geochimica et Cosmochimica Acta 69, 4101–4116 (2005).
Cerling, T. E. et al. Stable isotopes in elephant hair document migration patterns and diet changes. Proceedings of the National Academy of Sciences 103, 371–373 (2005).
Ayliffe, L. K. et al. Turnover of carbon isotopes in tail hair and breath CO2 of horses fed an isotopically varied diet. Oecologia 139, 11–22 (2004).
Uno, K. T., Quade, J. & Fisher, D. C. Bomb-curve radiocarbon measurement of recent biologic tissues and applications to wildlife forensics and stable isotope (paleo)ecology. Proceedings of the National Academy of Sciences 110, 11736–11741 (2013).
Laws, R. Dentition and ageing of the hippopotamus. African Journal of Ecology 6, 19–52 (1968).
Klingel, H. Hippopotamus in Mammals of Africa (eds. Kingdon, J. & Hoffmann, M. ) 68–78 (2013).
Klingel, H. Das flusspferde in Wilde Schweine und Flusspferde (eds. Macdonald, A. A. & Gansloßer, U. ) 353–370 (2008).
Vesey-FitzGerald, D. F. Grazing succession among East African game animals. Journal of Mammalogy 41, 161 (1960).
Field, C. R. The food habits of wild ungulates in Uganda by analyses of stomach contents. African Journal of Ecology 10, 17–42 (1972).
Boisserie, J. R. et al. Diets of modern and late Miocene hippopotamids: evidence from carbon isotope composition and micro-wear of tooth enamel. Palaeogeography, Palaeoclimatology, Palaeoecology 221, 153–174 (2005).
Cerling, T. E. et al. Dietary changes of large herbivores in the Turkana Basin, Kenya from 4 to 1 Ma. Proceedings of the National Academy of Sciences 112, 11467–11472 (2015).
Cerling, T. E. et al. Stable isotope ecology of the common hippopotamus. Journal of Zoology 276, 204–212 (2008).
Cerling, T. E. 14.12 Stable Isotope Evidence for Hominin Environments in Africa in Treatise on Geochemistry 14 (eds Cerling, T. E. ) 157–167 (Elsevier Ltd., 2014).
J T Verweij, R., Verrelst, J., Loth, P. E., M A Heitkönig, I. & M H Brunsting, A. Grazing lawns contribute to the subsistence of mesoherbivores on dystrophic savannas. Oikos 114, 108–116 (2006).
Hudak, A. T., Wessman, C. A. & Seastedt, T. R. Woody overstorey effects on soil carbon and nitrogen pools in South African savanna. Austral Ecology 28, 173–181 (2003).
Wiegand, K., Ward, D. & Saltz, D. Multi‐scale patterns and bush encroachment in an arid savanna with a shallow soil layer. Journal of Vegetation Science 16, 311–320 (2005).
Ward, D. Do we understand the causes of bush encroachment in African savannas? African Journal of Range and Forage Science 22, 101–105 (2005).
Wittemyer, G. et al. Illegal killing for ivory drives global decline in African elephants. Proceedings of the National Academy of Sciences 111, 13117–13121 (2014).
Plumptre, A. J., Kujirakwinja, D., Treves, A., Owiunji, I. & Rainer, H. Transboundary conservation in the greater Virunga landscape: its importance for landscape species. Biological Conservation 134, 279–287 (2007).
Dudley, J. P., Ginsberg, J. R., Plumptre, A. J., Hart, J. A. & Campos, L. C. Effects of War and Civil Strife on Wildlife and Wildlife Habitats. Conservation Biology 16, 319–329 (2002).
Olivier, R. Elephant Conservation Plan for Uganda (Uganda National Parks, 1991).
Acknowledgements
We wish to thank M. Dearing, F. Brown, and two anonymous reviewers who provided constructive comments on earlier drafts of this manuscript. We would also like to thank the Uganda National Counsel for Science and Technology for granting permission to conduct this research and export samples. We thank the members of the Research Monitoring and Conservation Unit at the Uganda Wildlife Authority who granted us permission to conduct this research, Aggrey Rwetsiba and Fred Kisame, and also Margaret Driciru, acting QENP park warden, and Nelson Guma, Conservation Area Manager. This study was funded by grants from the Geological Society of America (#9169111), the Global Change and Sustainability Center, NSF GRFP funding to KLC, the National Geographic Society (YEG #9349-13), a Leakey Foundation grant to SAB, the Wenner-Gren Foundation (#8694), NSF Doctoral Dissertation Improvement #1260535 Grant to SAB and an NSF grant #0621542 to TEC.
Author information
Authors and Affiliations
Contributions
K.L.C., S.A.B. and T.E.C. designed research; K.L.C., S.A.B. and T.E.C. collected data; H.K. provided samples; K.L.C. and S.A.B. performed analysis; K.L.C., S.A.B. and T.E.C. wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chritz, K., Blumenthal, S., Cerling, T. et al. Hippopotamus (H. amphibius) diet change indicates herbaceous plant encroachment following megaherbivore population collapse. Sci Rep 6, 32807 (2016). https://doi.org/10.1038/srep32807
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32807
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}