
Abstract
Interactions between climate, fire and CO2 are believed to play a crucial role in controlling the distributions of tropical woodlands and savannas, but our understanding of these processes is limited by the paucity of data from undisturbed tropical ecosystems. Here we use a 28,000-year integrated record of vegetation, climate and fire from West Africa to examine the role of these interactions on tropical ecosystem stability. We find that increased aridity between 28–15 kyr B.P. led to the widespread expansion of tropical grasslands, but that frequent fires and low CO2 played a crucial role in stabilizing these ecosystems, even as humidity changed. This resulted in an unstable ecosystem state, which transitioned abruptly from grassland to woodlands as gradual changes in CO2 and fire shifted the balance in favor of woody plants. Since then, high atmospheric CO2 has stabilized tropical forests by promoting woody plant growth, despite increased aridity. Our results indicate that the interactions between climate, CO2 and fire can make tropical ecosystems more resilient to change, but that these systems are dynamically unstable and potentially susceptible to abrupt shifts between woodland and grassland dominated states in the future.
Similar content being viewed by others


Introduction
Savanna-forest complexes occupy nearly 20% of the Earth’s land surface1, are responsible for 30% of the Earth’s terrestrial primary production and play an important role in global carbon budgets2. Anthropogenic climate change is anticipated to have a significant influence on the ecology and distribution of tropical ecosystems, though the magnitude, rate and direction of these changes are uncertain3. These uncertainties reflect the competing influences of moisture availability, CO2 and fire on tropical vegetation4,5,6. For example, although the dominant factor controlling the potential amount of woody cover in savanna ecosystems is precipitation, actual woody cover is controlled by landscape disturbance, much of which is attributable to fire7,8. The establishment of woody plants in fire-prone tropical grasslands depends on both the frequency of fires and growth rates by affecting whether plants can achieve sufficient stem diameters and heights to survive subsequent fires9,10. In ecosystems with frequent fires or slow growth rates, woody plants may be unable to escape this “fire trap” despite increases in moisture availability. Changes in CO2 may alter these relationships by changing the carbon available for woody plant growth9,10, and increased CO2 has been shown to increase growth rates, leading to increased survivability in the presence of fire9,11,12.
To better understand the stability of tropical forest-savanna systems, we use the sediment record from Lake Bosumtwi, Ghana, which is located in the lowland tropical forest, several hundred kilometers south of the modern-day savanna-forest boundary (Fig. 1). We reconstruct changes in vegetation and hydrology using the carbon and hydrogen isotope composition of sedimentary leaf wax n-alkanes (δ13Cwax, δDwax). Changes in δ13C are interpreted as reflecting changes in the relative proportions of plants using the C3 and C4 photosynthetic pathways13. C3 vegetation includes most woody plants and winter season grasses with average δ13C values that are as low as −36‰, whereas most summer or dry season C4 grasses have δ13C values of around −20‰14. We simultaneously reconstruct changes in precipitation using hydrogen isotope analysis of the same compounds (δDwax)15, which allows us to assess the relative phasing of vegetation and precipitation changes independent of any potential complications associated with organic matter transport pathways or residence time.

Modeled variations in vegetation cover computed as proportions of C3 and C4 plants replotted from41. White areas indicate deserts. Circle indicates the location of Lake Bosumtwi, Ghana (6°30′N, 1°25′W). Map produced using ESRI ArcGIS software [v.10.3.1], (http://www.esri.com/software/arcgis).
Changes in fire frequency are estimated from layer counts of distinct charcoal laminae within the annual sediment laminations (Fig. 2). Because the charcoal occurs as layers and is composed mostly of relatively intact fragments, we interpret the charcoal layers as reflecting the seasonal transport of charcoal to the lake from dry season fires with little or no time lag. We then use the frequency of charcoal layers as a proxy for annual fire frequency. Reconstructions of fire frequency are supported by fire reconstructions from concentrations of levoglucosan, a byproduct of cellulose combustion, and sediment particulate charcoal counts in pollen slides. Previous studies have shown that particulate charcoal16 and levoglucosan17 can provide reliable indicators of paleofire in some sedimentary archives and Lake Bosumtwi is an ideal location for preservation of the fire marker levoglucosan because of its permanent bottom water anoxia and well-preserved organic matter18. While all three approaches provide proxies of changes in fire regime, processes such as transport and deposition, fire intensity/temperature and fuel source will affect them differently, leading to differences in fire reconstructions19,20,21,22,23 (Fig. 3). However, the most significant changes are consistent between the fire proxy indicators, indicating that they do preserve consistent large scale variations in the fire regime (SOM).

Photomicrographs showing changes in the frequency of charcoal laminations in transmitted light sediment thin section from (A) the late Holocene (~1500 yr BP) when discrete bands of charcoal are absent, (B) the deglacial transition (~14,600 yr BP) when charcoal frequency has started to decline, (C) the glacial (~18,200 yr BP) when charcoal bands occur almost annually. Varves are evident in the millimeter to sub-millimeter scale couplets of light and dark laminae. The charcoal appears as distinct black layers in the boundaries of some of the annual laminae.

(A) Temporal variations in the climate-fire-CO2-vegetation system of tropical West Africa over the last 28,000 years BP reconstructed from Lake Bosumtwi. Atmospheric CO2 from the EPICA (triangles)42 and Taylor Dome ice cores43 (squares). Computed changes in Northern Hemisphere summer (JJA) insolation at 6.5°N44 (grey line). Reconstructed precipitation changes from the δD values of C31 n-alkanes (blue circles) from Lake Bosumtwi15. Reconstructed changes in fire from a 20-year moving average of the frequency of charcoal layers in thin sections (brown), concentrations of the cellulose combustion byproduct levoglucosan (ng/cm2/yr) orange circles) and particulate charcoal influx (particles/cm2/yr; pink diamonds). Relative proportion of C3 and C4 plants from the δ13C values of C31 n-alkanes. (B) Expanded view of the transition from dry, frequently burned, grassland dominated ecosystems to more humid, low fire and CO2 dominated forested ecosystems during the last deglaciation. The blue dashed line indicates the onset of humid conditions, the green dashed line indicates the midpoint of the abrupt deglacial vegetation shift.
Results
Evidence for a nonlinear tropical vegetation response to changes in hydroclimate
The Lake Bosumtwi record shows that the climate, fire and ecological regimes of tropical West Africa changed significantly over the past 28,000 years. During the early part of the record (Fig. 3A; 15,000–28,000 yr BP), δDwax values were positive, indicating more arid conditions, fire was more frequent and δ13Cwax values were higher (−17.3 to −20.6‰) indicating that the landscape was dominated by drought-tolerant C4 summer grasses with only minor contributions from C3 woody plants that burned almost annually. In comparison with the rest of the record, δ13Cwax values show little variability, despite significant variations in δDwax and fire at this time. After ca. 15,100 yr BP (Fig. 3B, blue dashed line), more depleted δDwax values indicate a gradual return to more humid conditions. These are accompanied by a decline in the frequency of fires, which disappear almost entirely for much of the most humid portion of the Holocene (i.e., 10,000–3,200 yr BP). However, in contrast to the gradual nature of these changes in precipitation and fire, the δ13Cwax record suggests that the vegetation response was abrupt, with a ~25% change in the proportion of C3 woody plants in <150 years. The rapid expansion of woody plants also occurs ca. 300 years after the onset of wetter conditions (at 14,800 yr BP), at the time when fire frequency had declined from nearly annual to one fire every five years (Fig. 3B, green dashed line). Over much of the remainder of the record, the landscape around Lake Bosumtwi was dominated by a variable mix of C3 and C4 plants suggestive of an open woodland or forest environment similar to today, though the data also suggest a trend towards an increasing proportion of C3 plants over the past 14,500 years. Coherent variations in δ13Cwax and δDwax, during this period indicate that the dominant control over changes in the relative proportions of C3/C4 plants was precipitation, as expected in the absence of fire disturbance. After ca. 3200 yr BP, δDwax values suggest that conditions became significantly more arid, and were accompanied by an increase in fire. Nevertheless, fire frequency during the late Holocene was much lower than during the late Pleistocene (27 to 15 ka), and the magnitude of the fire response recorded in the levoglucosan and particulate charcoal records differ, suggesting a distinctly different fire regime. Unlike the earlier part of the record, severe late Holocene aridity (∆δDwax = ~17‰) was associated with only minor changes in δ13Cwax, (∆δ13Cwax = ~2–3‰) indicating that woody plant-dominated ecosystems persisted despite these environmental changes.
The changing relationships between δ13Cwax and δDwax suggest that reconstructed vegetation changes over the past 28,000 years cannot be explained solely by precipitation but instead reflect the competing influences of climate, fire and atmospheric CO2, as suggested by modeling and modern ecosystem studies24,25. The invariant nature of the early portion of the δ13Cwax record, when compared with changes in δDwax, shows that these grasslands were insensitive to temporarily increased precipitation. The fire proxies also show that at this time, significant landscape-scale fires occurred at annual to interannual timescales. Our evidence for extensive grasslands across tropical Africa at this time with a high frequency of burning is supported by both vegetation synthesis and modeling studies26,27 and lower resolution fire reconstructions from marine sediment cores, which indicate that glacial conditions were accompanied by substantial export of fire-derived elemental carbon28. We hypothesize that these fires were sufficiently frequent and widespread to prevent woody plants from becoming established, as can occur in modern savannah ecosystems7,8. Prior to ~15,000 yr BP, this effect would have been exacerbated by low CO2 levels, which would have slowed the rate of woody plant germination, growth and recovery, making it difficult for C3 plants to recover from fire disturbance even as humidity increased4.
A critical role for fire and CO2 in controlling tropical vegetation stability
We propose that the high fire frequency early in the record is also the proximate cause of the abrupt and delayed expansion of C3 plants at 14,800 yr BP (Fig. 4). The rapid shift in vegetation occurred when precipitation was similar to earlier (>18,000 yr BP) and later (<3000 yr BP) periods when no systematic vegetation change happened, demonstrating that vegetation was not responding to changes in precipitation alone (Fig. 3). Instead, the abrupt vegetation change reflects a threshold response to the interactions between gradually declining fire frequency, increasing CO2 and increasing precipitation. The abruptness of this transition likely reflects a combination of the equilibration of C3 plant composition with available moisture and a feedback between woody plant expansion and a further reduction in the spread of subsequent grassland fires. Support for a threshold mechanism in controlling the vegetation response is also consistent with the ~300 year lag between the initial changes in δDwax and δ13Cwax, and the transition from a system where vegetation is unresponsive to changes in precipitation (>14,800 yr BP) to one where the dominant control on vegetation is precipitation (14,800–3,000 yr BP) (Fig. 3). Although other abrupt changes in vegetation occur in the record at ca. 11,500 yr BP and 5700 yr BP, these lack the characteristics indicative of a threshold vegetation response (Supplementary Information Fig. S4). For example, neither show clear evidence for a lag in the timing of the vegetation response to climate, nor is there evidence of changing sensitivity of the vegetation to environmental conditions. Both of these are evident in the shift at 14,800 yr BP.

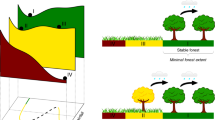
Left: cartoon of changes in the climate-ecosystem balance as a function of time. Right: schematic of changes in each parameter indicating the size and direction of change between timesteps (arrows) and the relationships between the parameters (red: negative feedback, blue: positive feedback, circle: hypothesized driver of the relationship, dashed: the relationship between these two parameters is weakened by other feedback relationships.
Following the initial expansion of C3 plants and for much of the remaining record, the frequency and intensity of fire remained low, CO2 was high and increasing, and vegetation responded on centennial to millennial timescales to changes in precipitation, with greater proportions of C3 plants accompanying wetter conditions (Fig. 3). However, this interval is also characterized by an overall trend towards greater C3 plant abundance and a decrease in the sensitivity of vegetation to changes in precipitation. We hypothesize that this reflects the influence of increasing CO2, which favors the growth of C3 plants relative to C4 grasses29. These effects would also have been enhanced by biogeophysical feedbacks; well-developed forests can act to suppress the growth and colonization of grasses by reducing ground light availability, maintaining higher sub-canopy humidity and keeping temperatures lower, all of which promote the growth of C3 plants and are capable of maintaining forest stability30, even as environmental conditions change.
Discussion
The Lake Bosumtwi record highlights the competing roles of climate, fire and CO2 on the long-term stability and evolution of grasslands and forests of tropical West Africa. In agreement with many studies of modern tropical grassland ecosystems, frequent fires played a crucial role in excluding woody plants during glacial to late glacial times, even as precipitation and CO2 increased4,5,6,9,10,31 (Fig. 4). However, this also resulted in a state of disequilibrium between climate and vegetation, leading to an abrupt vegetation response to changes in climate. In contrast, dramatically increased aridity and fire in the late Holocene did not cause a substantial increase in drought-tolerant C4 grasses, suggesting that tropical forests became more resilient to changes in climate and disturbance. We propose that this is a consequence of high CO2 and biogeophysical feedbacks, which resulted in conditions more favorable for C3 plant growth and recovery, stabilizing existing forest cover despite the increased aridity (Fig. 4). We hypothesize that, as with the late glacial grasslands, this has resulted in a dynamically unstable climate-vegetation state that is susceptible to rapid transitions between C3 and C4 plant-dominated landscapes32. Although elevated CO2 should continue to act as a stabilizing element, at higher CO2 the advantages for C3 plants decrease33, limiting its ability to stabilize these systems and increasing the potential for an abrupt collapse of tropical forest ecosystems in response to increased aridity. The sensitivity of these ecosystems to slight perturbations in climate today is evident in the Dahomey Gap, just to the east of Lake Bosumtwi, where precipitation is only 200–400 mm yr−1 lower than Bosumtwi but the natural landscape is dominated by grasslands with only isolated patches of forest34. Here, vegetation records indicate that small increases in rainfall resulted in wholesale return of forest cover briefly in the late Holocene35. In the future, much of West Africa could become more arid, provided that projected increases in precipitation are insufficient to overcome a net drying of soils driven by increased temperature36. Such gradual changes in hydroclimate and fire could potentially overwhelm the natural buffering capacity of high CO2 and allow current climate-vegetation stability to give way to rapid forest-savanna transformation in a region that is already a mosaic of these two land-cover types.
Methods
Study site
Past variations in the climate, vegetation and fire history of West Africa were reconstructed using the sediment geochemical record from Lake Bosumtwi, Ghana. Lake Bosumtwi is a small (~8 km diameter), internally draining lake occupying a meteorite impact crater in southern Ghana (Supplementary Fig. S1). The lake is deep (~75 m depth asl) and sheltering by the crater walls inhibit mixing, leading to stratification and permanently anoxic bottom waters. This limits bioturbation and allows for the preservation of mm-scale laminations that have been demonstrated to be annual18. The majority of the catchment is forested except for the flat lying terraces and drainages, which have been converted to agriculture37. The regional climate is controlled primarily by the West African monsoon, which brings moisture to southern Ghana in May-September when the Intertropical Convergence Zone (ITCZ) migrates to the north (Supplementary Fig. S1). In winter, as the ITCZ shifts southward, the climate of the region is dominated by dry northwesterly winds, which inhibit rainfall.
Age model development
A detailed description of the age model used in this study is published elsewhere38. It is based on a combination of radiocarbon dating, Bayesian age depth modeling and varve counting. The chronology consists of 107 radiocarbon dates on bulk organic matter and macrofossils covering the upper 21.4 meters of sediment. Dates from different cores were correlated using marker laminae matches and Bayesian age depth modeling was performed using the R software package BACON39 and using the IntCal09 radiocarbon calibration curve40. Over the deglacial section of the core, we revised the age model by wiggle matching the uncalibrated radiocarbon ages to the IntCal09 calibration curve using the varves as a constraint on the time elapsed between radiocarbon age. An optimal fit was determined using Χ2 minimization38.
Lipid and compound-specific stable isotope analysis of n-alkanes
Sediment samples (1–5 g) were freeze dried, homogenized and solvent-extracted by either accelerated solvent extraction (Dionex ASE 200) or a microwave extraction (MARS) using dichloromethane:methanol (9:1; v/v). n-alkanes and levoglucosan were isolated from the total lipid extract by silica gel column chromatography. n-alkanes were additionally isolated by silver nitrate silica gel chromatography and molecular sieve or urea adduction. Stable carbon and hydrogen isotope analysis of the long chain n-alkanes was performed by gas chromatography isotope ratio mass spectrometry (GC-IR-MS). Description of the methods for δDwax analysis and corrections for changes in vegetation and ice volume are described in the online Supplementary Materials. δ13Cwax values were measured against internal calibrated propane reference gas standards and are reported in % VPDB (Vienna Pee Dee Belemnite). The precision of the analysis, based on repeated analysis of the standard mix was better than 0.3%. Each sample was measured in triplicate and the mean precision for the long chain n-alkanes (C27, C29, C31, C33) based on this replicate analysis was better than 0.4%. Levoglucosan analysis was performed by gas chromatography mass spectrometry. Identification of the trimethylsilyl ether derivative of levoglucosan was identified in the mass scan by its characteristic fragments (m/z 363, 333, 217, 204) and its retention time determined with an authentic standard. Quantification was performed using an internal standard (androstanol). Levoglucosan concentrations (ppm) were normalized to time (ppm/yr) using varve counts over the laminated intervals of the core and using the Bacon age-depth model over the unlaminated sections.
Reconstruction of fire frequency from sedimentary charcoal
Paleofire was reconstructed from a combination of microscopic particulate charcoal counts and frequency analysis of charcoal layers visually identified in sediment thin sections under transmitted light (Supplementary Fig. S2). Particulate charcoal fragments above 10 μm were counted on slides prepared for palynological analysis. A minimum of 2000 charcoal particles were counted per sample and abundance relative to an exotic marker, Lycopodium, was calculated. Charcoal counts were normalized to time (particles/cm2/yr) using the same approach as for levoglucosan. Fire frequency was estimated using visual identification of annual charcoal layers in sediment thin sections under transmitted light. Previous work on the laminations from Lake Bosumtwi has demonstrated that they are annual in nature and visible charcoal occurs as a distinct layer within many of the annual laminations, suggesting that the charcoal is deposited after seasonal fire events, rather than being remobilized material from fires occurring in previous years. Charcoal layer frequency analysis was performed on a single continuous set of thin sections spanning the record. Reproducibility was assessed via replication over a several short, overlapping intervals.
Additional Information
How to cite this article: Shanahan, T. M. et al. CO2 and fire influence tropical ecosystem stability in response to climate change. Sci. Rep. 6, 29587; doi: 10.1038/srep29587 (2016).
References
Scholes, R. J. & Archer, S. R. Tree-grass interactions in savannas. Annual Review of Ecological Systems 28, 517–544 (1997).
Grace, J., Jose, J. S., Meir, P., Miranda, H. S. & Montes, R. A. Productivity and carbon fluxes of tropical savannas. Journal of Biogeography 33, 387–400 (2006).
Settele, J. et al. In Climate Change. Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds C. B. Field et al.) 271–359 (Cambridge University Press, 2014).
Bond, W. J. & Midgley, G. F. Carbon dioxide and the uneasy interactions of trees and savannah grasses. Philosphical Transactions of the Royal Society B 367, 601–612 (2012).
Higgins, S. I. & Scheiter, S. Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nature 488, 209–213 (2012).
Lehmann, C. E. R. et al. Savanna Vegetation-Fire-Climate Relationships Differ Among Continents. Science 343, 548–552 (2014).
Bond, W. J. Large parts of the world are brown or black: a different view on the “Green World Hypothesis”. Journal of Vegetation Science 16, 261–266 (2005).
Sankaran, M. et al. Determinants of woody cover in African savannas. Nature 438, 846–849 (2005).
Bond, W. J. What Limits Trees in C4 Grasslands and Savannas? Annual Review of Ecology, Evolution, and Systematics 39, 641–659 (2008).
Higgins, S. I., Bond, W. J. & Trollope, W. S. W. Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna. Journal of Ecology 88, 213–229 (2000).
Kgope, B. S., Bond, W. J. & Midgley, G. F. Growth responses of African savanna trees implicate atmospheric CO2 as a driver of past and current changes in savanna tree cover. Austral Ecology 35, 451–463 (2010).
Idso, S. B. Shrubland expansion in the American southwest. Climatic Change 22, 85–86 (1992).
Farquhar, G. On the Nature of Carbon Isotope Discrimination in C4 Species. Functional Plant Biology 10, 205–226 (1983).
Chikaraishi, Y., Naraoka, H. & Poulson, S. R. Hydrogen and carbon isotopic fractionations of lipid biosynthesis among terrestrial (C3, C4 and CAM) and aquatic plants. Phytochemistry 65, 1369–1381 (2004).
Shanahan, T. M. et al. The time-transgressive termination of the African Humid Period. Nature Geoscience 8, 140–144 (2015).
Clark, R. L. Point count estimation of charcoal in pollen preparations and thin sections of sediments. Pollen et spores 25, 523–535 (1982).
Elias, V., Simoneit, B., Cordeiro, R. & Turcq, B. Evaluating levoglucosan as an indicator of biomass burning in Carajas, Amazonia: A comparison to the charcoal record. Geochimica et Cosmochimica Acta 65, 267–272 (2001).
Shanahan, T. M. et al. The formation of biogeochemical laminations in Lake Bosumtwi, Ghana, and their usefulness as indicators of past environmental changes. Journal of Paleolimnology 40, 339–355 (2008).
Clark, J. S. Fire and climate change during the last 750 yr in northwestern Minnesota. Ecological Monographs 60, 135–159 (1990).
Kuo, L.-J., Herbert, B. E. & Louchouarn, P. Can levoglucosan be used to characterize and quantify char/charcoal black carbon in environmental media? Organic Geochemistry 39, 1466–1478 (2008).
Leys, B., Brewer, S. C., McConaghy, S., Mueller, J. & McLauchlan, K. K. Fire history reconstruction in grassland ecosystems: amount of charcoal reflects local area burned. Environmental Research Letters 10, 114009 (2015).
Lynch, J. A., Clark, J. S. & Stocks, B. J. Charcoal production, dispersal, and deposition from the Fort Providence experimental fire: interpreting fire regimes from charcoal records in boreal forests. Canadian Journal of Forest Research 34, 1642–1656 (2004).
Whitlock, C. & Millspaugh, S. H. Testing the assumptions of fire-history studies: an examination of modern charcoal accumulation in Yellowstone National Park, USA. The Holocene 6, 7–15 (1996).
Bragg, F. J. et al. Stable isotope and modelling evidence for CO2 as a driver of glacial–interglacial vegetation shifts in southern Africa. Biogeosciences 10, 2001–2010 (2013).
Martin Calvo, M., Prentice, I. C. & Harrison, S. P. Climate vs. carbon dioxide controls on biomass burning: a model analysis of the glacial-interglacial contrast. Biogeosciences Discussions 11, 2569–2593 (2014).
Harrison, S. P. & Prentice, C. I. Climate and CO2 controls on global vegetation at the last glacial maximum: analysis based on paleovegetation data, biome modeling and paleoclimate simulations. Global Change Biology 9, 983–1004 (2003).
Prentice, I. C. & Jolly, D. Mid-Holocene and glacial-maximum vegetation geography of the northern continents and Africa. Journal of Biogeography 27, 507–519 (2000).
Verardo, D. J. & Ruddiman, W. F. Late Pleistocene charcoal in tropical Atlantic deep-sea sediments: climatic and geochemical significance. Geology 24, 855–857 (1996).
Ehleringer, J. R., Cerling, T. E. & Helliker, B. R. C4 photosynthesis, atmospheric CO2, and climate. Oecologia 112, 285–299 (1997).
Cochrane, M. A. Fire science for rainforests. Nature 421, 913–919 (2003).
Hoffmann, W. A. et al. Ecological thresholds at the savanna-forest boundary: how plant traits, resources and fire govern the distribution of tropical biomes. Ecology Letters 15, 759–768 (2012).
Renssen, H., Brovkin, V., Fichefet, T. & Goosse, H. Simulation of the Holocene climate evolution in Northern Africa: The termination of the African Humid Period. Quaternary International 150, 95–102 (2006).
Cowling, S. A. & Sage, R. F. Interactive effects of low atmospheric CO2 and elevated temperature on growth, photosynthesis and respiration in Phaseolus vulgaris. Plant, Cell & Environment 21, 427–435 (1998).
Jenik, J. The Dahomey-Gap: an important issue in African phytogeography. Mémoire de la Société Biogéographique IV, 125–133 (1994).
Salzmann, U. & Hoelzmann, P. The Dahomey Gap: an abrupt climatically induced rain forest fragmentation in West Africa during the late Holocene. Holocene 15, 190–199 (2005).
Collins, M. et al. In Climate Change: The Physical Science Basis . Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds T. F. Stocker et al.) (Cambridge University Press, 2013).
Shanahan, T. M. et al. Spatial and temporal variability in sedimentological and geochemical properties of sediments from an anoxic crater lake in West Africa: Implications for paleoenvironmental reconstructions. Paleogeogr. Paleoclimatol. Paleoecol. 374, 96–109 (2013).
Shanahan, T. M. et al. Late Quaternary sedimentological and climate changes at Lake Bosumtwi Ghana: New constraints from laminae analysis and radiocarbon age modeling. Paleogeogr. Paleoclimatol. Paleoecol. 361, 49–60 (2012).
Blaauw, M. & Christen, J. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Analysis 6, 457–474 (2011).
Reimer, P. J. et al. INTCAL09 and MARINE09 radiocabon age calibration curves, 0–50,000 Years cal BP. Radiocarbon 51, 1111–1150 (2009).
Still, C. J. & Powell, R. L. Isoscapes: Understanding Movement, Pattern, and Process on Earth Through Isotope Mapping. (Springer, 2010).
Monnin, E. et al. Atmospheric CO2 Concentrations over the Last Glacial Termination. Science 291, 112–114 (2001).
Indermühle, A., Monnin, E., Stauffer, B., Stocker, T. F. & Wahlen, M. Atmospheric CO2 concentration from 60 to 20 kyr BP from the Taylor Dome ice core, Antarctica. Geophysical Research Letters 27, 735–738 (2000).
Laskar, J. et al. A long term numerical solution for the insolation quantities of the Earth. Astronomy and Astrophysics 428, 261–285 (2004).
Acknowledgements
Additional information and data are available in the online Supplementary materials. This work was supported by NSF grants EAR0601998, EAR0602355, AGS0402010, ATM0401908, ATM0214525, ATM0096232 and AGS1243125 and a Chevron Centennial Fellowship at the University of Texas at Austin awarded to T.M.S.
Author information
Authors and Affiliations
Contributions
T.M.S. and K.A.H. generated the data and prepared Figures 2 and 3. N.M. prepared Figure 1. W.D.G. and C.S.M. generated the particulate charcoal data. T.M.S., K.A.H., N.M. and J.T.O. prepared the written manuscript. J.A.P., C.A.S., J.W.K. and C.W.H. assisted with experimental design, fieldwork and sampling. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Shanahan, T., Hughen, K., McKay, N. et al. CO2 and fire influence tropical ecosystem stability in response to climate change. Sci Rep 6, 29587 (2016). https://doi.org/10.1038/srep29587
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep29587
This article is cited by
-
Spatial distribution of Vachellia karroo in Zimbabwean savannas (southern Africa) under a changing climate
Ecological Research (2018)
-
Local and regional wildfire activity in central Maine (USA) during the past 900 years
Journal of Paleolimnology (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
