
Abstract
Acetoin is widely used in food and cosmetic industry as taste and fragrance enhancer. For acetoin production in this study, Saccharomyces cerevisiae JHY605 was used as a host strain, where the production of ethanol and glycerol was largely eliminated by deleting five alcohol dehydrogenase genes (ADH1, ADH2, ADH3, ADH4, and ADH5) and two glycerol 3-phosphate dehydrogenase genes (GPD1 and GPD2). To improve acetoin production, acetoin biosynthetic genes from Bacillus subtilis encoding α-acetolactate synthase (AlsS) and α-acetolactate decarboxylase (AlsD) were overexpressed, and BDH1 encoding butanediol dehydrogenase, which converts acetoin to 2,3-butanediol, was deleted. Furthermore, by NAD+ regeneration through overexpression of water-forming NADH oxidase (NoxE) from Lactococcus lactis, the cofactor imbalance generated during the acetoin production from glucose was successfully relieved. As a result, in fed-batch fermentation, the engineered strain JHY617-SDN produced 100.1 g/L acetoin with a yield of 0.44 g/g glucose.
Similar content being viewed by others
Introduction
Acetoin, also known as 3-hydroxy-2-butanone or acetylmethylcarbinol, is widely used in food industry as a flavor enhancer, giving a buttery taste1. It can also be used as a building block for various chemicals such as alkyl pyrazines, diacetyl, and acetylbutanediol2,3,4. Currently, most of commercial acetoin is produced by chemical synthesis, but the use of such non-natural acetoin is restricted in some applications, especially in food and cosmetic industry, because of safety concerns. Accordingly, many attempts have been reported to produce natural acetoin by biological process, including enzyme conversion and microbial fermentation1,5,6,7.
Many microorganisms, such as Bacillus subtilis, Bacillus amyloliquefaciens, Enterobacter cloacae, Serratia marcescens, and Paenibacillus polymyxa, can produce acetoin from pyruvate via α-acetolactate by two enzymatic steps catalyzed by α-acetolactate synthase and α-acetolactate decarboxylase1,8. Acetoin can be further converted to 2,3-butanediol by 2,3-butanediol dehydrogenase (also known as acetoin reductase) using NADH as a cofactor. Therefore, to accumulate acetoin, 2,3-butanediol production was inhibited in various bacteria mainly by adopting two strategies; disruption of 2,3-butanediol dehydrogenase and overexpression of NADH oxidase. Butanediol dehydrogenase-blocked B. subtilis (JNA-UD-6), isolated after mutagenesis using UV irradiation with diethyl sulfate, showed a 24.3% increase in acetoin production and a 39.8% decrease in 2,3-butanediol production compared with the parental strain in batch fermentation, and produced 53.9 g/L acetoin after 144 h fermentation in fed-batch fermentation9. On the other hand, NADH oxidase, which converts NADH to NAD+, was overexpressed to reduce NADH-dependent 2,3-butanediol production. In S. marcescens H32, introduction of NADH oxidase from Lactobacillus brevis decreased 2,3-butanediol titer by 48% and increased acetoin titer by 33%10. Both of these strategies have also been applied to B. subtilis and E. cloacae, resulting in 56.7 g/L and 55.2 g/L acetoin production, respectively11,12.
Saccharomyces cerevisiae, which is classified as generally recognized as safe (GRAS) microorganism, has been considered as a key cell factory platform for producing valuable chemicals because of its tolerance and robustness toward industrial conditions. Acetoin accumulation has been reported in commercial wine yeast engineered for glycerol overproduction13. The engineered strain overexpressing GPD1 and deleting ALD6 (BC GPD1 ald6) produced 26.9 g/L glycerol with 9.5 g/L acetoin from 200 g/L glucose. In addition, several efforts have been made to engineer S. cerevisiae to produce 2,3-butanediol, a neighboring metabolite of acetoin, leading to a significant improvement in both titer and yield of 2,3-butanediol production14,15,16. Previously, we monitored acetoin production levels to investigate aromatic amino acids-inducible promoter system17. Nevertheless, the study focused on acetoin production in S. cerevisiae has not yet been reported.
In our previous study, we developed S. cerevisiae strain for efficient production of 2,3-butanediol by introducing heterologous acetoin biosynthetic pathway from B. subtilis, overexpressing 2,3-butanediol dehydrogenase, and eliminating major byproduct pathways involved in ethanol and glycerol production14. Furthermore, the cofactor imbalance generated during 2,3-butanediol production in the engineered strain was restored by overexpressing water-forming NADH oxidase from Lactococcus lactis. In this study, for efficient production of acetoin, we additionally disrupted 2,3-butanediol dehydrogenase in adh1-5Δgpd1Δgpd2Δ strain and adopted above strategies comprised of introducing heterologous acetoin pathway and redox rebalancing.
Results and Discussion
Introduction of acetoin biosynthetic pathway in adh1-5Δgpd1Δgpd2Δ strain (JHY605)
To produce acetoin in S. cerevisiae, α-acetolactate synthase (alsS) and α-acetolactate decarboxylase (alsD) genes from B. subtilis were introduced into JHY605 strain (adh1-5Δgpd1Δgpd2Δ), which lacks five alcohol dehydrogenases (Adh1 to Adh5) and two glycerol 3-phosphate dehydrogenases (Gpd1 and Gpd2) (Fig. 1). The fermentation profiles of the control strain JHY605-C harboring empty vector p413GPD and strain JHY605-SD expressing alsS and alsD from p413-SD plasmid are shown in Fig. 2. JHY605-C produced only a trace amount of acetoin (0.1 g/L) and 1.4 g/L of 2,3-butanediol from 31.6 g/L of glucose after 96 h fermentation (Fig. 2a). Although five alcohol dehydrogenase genes were deleted, JHY605-C produced 3.8 g/L of ethanol as a major end product. This might be because pyruvate generated by glycolysis is mainly metabolized to ethanol production via pyruvate decarboxylases and remaining ADH isozymes including Sfa1, Adh6, and Adh7. In agreement with previous study, glycerol pathway was completely blocked in JHY605-C by the deletion of GPD1 and GPD214,18.

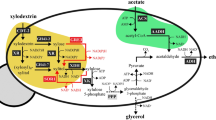
Two molecules of pyruvate are converted to one molecule of acetoin by α-acetolactate synthase (AlsS) and α-acetolactate decarboxylase (AlsD). Acetoin can be further reduced to 2,3-butanediol by 2,3-butanediol dehydrogenase (Bdh1). The water-forming NADH oxidase (NoxE) catalyzes the oxidation of NADH to NAD+ using oxygen as an electron acceptor. Dashed arrows indicate multiple enzymatic steps. The crosses represent blockage of the pathway by deleting the corresponding genes. DHAP, dihydroxyacetone phosphate; G3P, glycerol-3-phosphate; GAP, glyceraldehyde-3-phosphate.

Strain JHY605 (adh1-5Δgpd1Δgpd2Δ) harboring empty p413GPD plasmid (JHY605-C) (a) or p413-SD (JHY605-SD) (b) was grown in 10 mL SC-His media containing 50 g/L glucose in a 100 mL flask. Error bars indicate standard deviations of three independent experiments.
Whereas, strain JHY605 harboring p413-SD plasmid (JHY605-SD) showed an increase in glucose consumption rate and produced up to 5.9 g/L acetoin (Fig. 2b). Furthermore, 2,3-butanediol production level increased to 9.3 g/L, even though 2,3-butanediol dehydrogenase was not overexpressed, reflecting the endogenous 2,3-butanediol dehydrogenase activity in S. cerevisiae. By introducing this competing pathway, ethanol production yield was reduced to 0.02 g/g glucose in JHY605-SD compared with that in JHY605-C (0.12 g/g glucose).
In S. cerevisiae, NAD+ regeneration for glycolysis is mainly achieved by producing ethanol and glycerol (Fig. 1). Therefore, the growth defects of strain JHY605 might be due to the accumulation of NADH. In addition, the residual ADH activity might not be enough to prevent the accumulation of toxic acetaldehyde (Fig. 1). Introduction of acetoin biosynthetic pathway into JHY605 might relieve the growth defects by reducing acetaldehyde formation through efficient conversion of pyruvate to acetoin, and also by partly restoring NAD+ regeneration through 2,3-butanediol production (Fig. 1).
Disruption of 2,3-butanediol dehydrogenase BDH1 to improve acetoin production
By introducing acetoin biosynthetic pathway into JHY605, pyruvate flux was successfully redirected toward acetoin pathway. However, acetoin was further converted to 2,3-butanediol, resulting in about 1.5-fold higher titer of 2,3-butanediol than that of acetoin. In S. cerevisiae, Bdh1 is a major enzyme catalyzing the reduction of acetoin to 2,3-butanediol (Fig. 1). Therefore, we further deleted BDH1 gene in JHY605, resulting in strain JHY617. When acetoin biosynthetic pathway was introduced into strain JHY617 (JHY617-SD), 2,3-butanediol production from acetoin was significantly reduced to 0.2 g/L, which then contributed to the increase in acetoin production accordingly. As a result, up to 15.4 g/L acetoin was produced after 72 h fermentation in SC-His medium containing 50 g/L glucose, with a yield of 0.30 g/g glucose (Fig. 3a). The trace amount of 2,3-butanediol production in JHY617-SD might be mediated by other minor putative enzymes such as D-arabinose dehydrogenase (Ara1) having 2,3-butanediol dehydrogenase activity19.

Strain JHY617 (adh1-5Δgpd1Δgpd2Δbdh1Δ) harboring p413-SD (JHY617-SD) (a) or p413-SDN (JHY617-SDN) (b) was grown in 10 mL SC-His containing 50 g/L glucose in a 100 mL flask. (c) NADH/NAD+ ratios in JHY617-SD and JHY617-SDN. Error bars indicate standard deviations of three independent experiments.
Recovering redox imbalance by expressing water-forming NADH oxidase noxE
Cofactor balance, especially NADH/NAD+ ratio plays an important role in a large number of biochemical reactions20,21. Thus, maintaining the cofactor balance is an essential requirement for sustaining cellular metabolism and cell growth22. In acetoin production pathway, NADH produced from glycolysis could not be converted to NAD+, leading to a redox cofactor imbalance. Furthermore, since NADH-dependent metabolic pathways, related to the production of ethanol, glycerol, and 2,3-butanediol, were disrupted in strain JHY617-SD, the redox imbalance might be more severe. Therefore, as an effort to resolve the redox imbalance in JHY617-SD, we introduced noxE from L. lactis, encoding water-forming NADH oxidase. To this end, FBA1 promoter controlled-noxE was inserted to the acetoin biosynthetic plasmid p413-SD, resulting in p413-SDN. Strain JHY617 harboring p413-SDN (JHY617-SDN) showed a significant improvement in glucose consumption rate, thereby taking less time (~48 h) to completely ferment 50 g/L glucose than it was taken for JHY617-SD (~72 h) (Fig. 3). Moreover, acetoin production was improved up to 20.1 g/L with a yield of 0.39 g/g glucose, reaching 80% of maximum theoretical yield. Accordingly, strain JHY617-SDN exhibited about two-fold increase in acetoin productivity (0.42 g/(L·h)) compared with JHY617-SD (0.21 g/(L·h)), suggesting that redox imbalance caused by acetoin production was successfully alleviated by expressing NADH oxidase (Table 1). To confirm the effect of noxE expression on redox state, we analyzed intracellular NADH/NAD+ ratios in JHY617-SD and JHY617-SDN. As expected, the NADH/NAD+ ratios in JHY617-SDN were lower than those in JHY617-SD throughout the growth phase, demonstrating the efficient conversion of NADH to NAD+ by NoxE (Fig. 3c).
Fed-batch fermentation for acetoin production
To evaluate the potential of JHY617-SDN as a host strain for acetoin production, fed-batch fermentation was performed with intermittent feeding of glucose and pH control. JHY617-SDN was grown in YPD medium containing 100 g/L glucose with initial OD600 of 9.5. In fed-batch fermentation, up to 100.1 g/L acetoin was produced with a yield of 0.44 g/g glucose after 55 h cultivation, reaching 90% of maximum theoretical yield (Fig. 4). Moreover, acetoin productivity was dramatically improved to 1.82 g/(L·h). Taken together, JHY617-SDN showed superior performance of acetoin production compared with the host strains reported in previous studies (Table 2). Notably, both acetoin titer and yield in this study are the highest among these studies. Although acetoin productivity reported in S. marcescens and E. cloacae were higher than that of our study10,12, these strains have potential pathogenicity23,24.

Strain JHY617-SDN was cultivated in YPD medium containing 100 g/L glucose with initial OD600 of 9.5. Glucose was intermittently added into culture medium using the feeding solution (800 g/L glucose) before glucose was completely consumed.
In this study, we developed S. cerevisiae strain for efficient production of acetoin by introducing heterologous acetoin pathway from B. subtilis and eliminating 2,3-butanediol dehydrogenase using JHY605 as a host strain, where the production of ethanol and glycerol was largely eliminated. In addition, cofactor imbalance generated during acetoin production was successfully alleviated by expressing NADH oxidase from L. lactis, leading to significantly enhanced acetoin production. As a result, to the best of our knowledge, the highest titer and yield in microbial production of acetoin were achieved. These results suggest that S. cerevisiae might be a promising host for the production of acetoin.
Methods
Strains and media
All strains used in this study are described in Table 3. JHY617 strain, a BDH1 deletion mutant derived from JHY60514, was generated by PCR-mediated homologous recombination. The bdh1Δ::KanMX6 cassette flanked by 300 bp upstream and 282 bp downstream of the BDH1 open reading frame was obtained by PCR amplification from genomic DNA of bdh1Δ strain (BY4741 bdh1Δ::KanMX6, EUROSCARF) as a template, using the primer pair of d_BDH1 F (5′-GATTTGCTCACGCTACTTTG-3′) and d_BDH1 R (5′-GCCATGCTTTGTTTTAGACG-3′). The resulting PCR product was transformed into JHY605 strain and transformants were selected on YPD plate (10 g/L yeast extract, 20 g/L bacto-peptone, and 20 g/L glucose) supplemented with 200 μg/mL G418 sulfate (AG Scientific, Inc.)
Yeast cells were cultured in YPD medium or in synthetic complete medium lacking histidine (SC-His) (20 or 50 g/L glucose, 6.7 g/L yeast nitrogen base without amino acids, and 1.92 g/L amino acids mixture lacking histidine).
Plasmid construction
Plasmids used in this study are described in Table 4. The recombinant plasmids for acetoin pathway were constructed by using the multiple cloning system as previously described with minor modifications14. The alsS-expression cassette (PTDH3-alsS-TCYC1) flanked by MauBI and and NotI sites was obtained by PCR from p413_PTDH3-alsS-TCYC1 using the primers, Univ F2 (5′-GACTCGCGCGCGGGAACAAAAGCTGGAGCTC-3′) and Univ R (5′-GACTACGCGTGCGGCCGCTAATGGCGCGCCATAGGGCGAATTGGGTACC-3′), and cloned into AscI and NotI sites of p413-D plasmid14, resulting in p413-SD. The noxE-expression cassette (PFBA1-noxE-TFBA1) was additionally cloned into p413-SD as previously described14, resulting in p413-SDN.
Fermentation conditions
For flask fermentation, yeast cells harboring appropriate plasmids were pre-cultured in 5 mL of SC-His medium containing 20 g/L glucose in a 50 mL flask, inoculated to OD600 of 0.3 in 10 mL of SC-His medium containing 50 g/L glucose in a 100 mL flask, and then cultivated at 30 °C with shaking at 170 rpm.
Fed-batch fermentation was performed in 500 mL YPD medium containing 100 g/L glucose using a 1 L bench-top fermenter FMT-DS (Fermentec, Korea) at 30 °C with agitation speed of 500 rpm and air flow rate of 1.0 vvm. The pH of the culture medium was maintained at 5.5 by using 4 N NaOH. Strain JHY617-SDN was pre-cultured in SC-His medium containing 20 g/L glucose and inoculated into the fermenter with initial OD600 of 9.5. The feeding solution (800 g/L glucose) was intermittently added to the culture medium when the glucose concentration was lower than 20 g/L.
Analytical methods
Cell growth was determined by measuring the optical density at 600 nm (OD600). To analyze profile of metabolites, 1 mL of culture supernatants were collected and filtered through a 0.22 μm syringe filter. The concentrations of glucose, glycerol, acetoin, 2,3-butanediol, and ethanol were determined by high performance liquid chromatography (HPLC) using UltiMate 3000 HPLC system (Thermo fishers scientific) equipped with a BioRad Aminex HPX-87H column (300 mm×7.8 mm, 5 μm, Bio-rad) at 60 °C with 5 mM H2SO4 at a flow rate of 0.6 mL/min and refractive index (RI) detector at 35 °C. The intracellular concentrations of NADH and NAD+ were measured using EnzyChrom™ NAD/NADH Assay Kit (E2ND-100, BioAssay Systems). Strains JHY617-SD and JHY617-SDN were harvested at different time points of fermentation and washed with cold phosphate-buffered saline (PBS; 8 g/L NaCl, 0.2 g/L KCl, 1.42 g/L Na2HPO4, 0.24 g/L KH2PO4 [pH 7.4]) solution. Cells of OD600 of 1.0 were pelleted and analyzed according to the manufacturer’s instructions.
Additional Information
How to cite this article: Bae, S.-J. et al. Efficient production of acetoin in Saccharomyces cerevisiae by disruption of 2,3-butanediol dehydrogenase and expression of NADH oxidase. Sci. Rep. 6, 27667; doi: 10.1038/srep27667 (2016).
References
Xiao, Z. & Lu, J. R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 32, 492–503 (2014).
Xiao, Z., Ma, C., Xu, P. & Lu, J. R. Acetoin catabolism and acetylbutanediol formation by Bacillus pumilus in a chemically defined medium. PLoS One 4, e5627 (2009).
Chen, H. et al. Shanghai Apple Flavor & Fragrance Co. Ltd. A preparation method of butadione by gas-phase oxidating 3-hydroxy-butanone. European patent EP1826194 A1. 2007 Aug 29.
Xiao, Z., Hou, X., Lyu, X., Xi, L. & Zhao, J. Accelerated green process of tetramethylpyrazine production from glucose and diammonium phosphate. Biotechnol. Biofuels 7, 106 (2014).
Wang, X. et al. Efficient bioconversion of 2,3-butanediol into acetoin using Gluconobacter oxydans DSM 2003. Biotechnol. Biofuels 6, 155 (2013).
Li, S., Xu, N., Liu, L. & Chen, J. Engineering of carboligase activity reaction in Candida glabrata for acetoin production. Metab. Eng. 22, 32–39 (2014).
Bai, F. et al. Engineered Serratia marcescens for efficient (3R)-acetoin and (2R, 3R)-2,3-butanediol production. J. Ind. Microbiol. Biotechnol. 42, 779–786 (2015).
Xiao, Z. & Xu, P. Acetoin metabolism in bacteria. Crit. Rev. Microbiol. 33, 127–140 (2007).
Zhang, X. et al. Mutation breeding of acetoin high producing Bacillus subtilis blocked in 2,3-butanediol dehydrogenase. World J. Microbiol. Biotechnol. 29, 1783–1789 (2013).
Sun, J. A., Zhang, L. Y., Rao, B., Shen, Y. L. & Wei, D. Z. Enhanced acetoin production by Serratia marcescens H32 with expression of a water-forming NADH oxidase. Bioresour. Technol . 119, 94–98 (2012).
Zhang, X. et al. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis . Metab. Eng. 23, 34–41 (2014).
Zhang, L. et al. Biotechnological production of acetoin, a bio-based platform chemical, from a lignocellulosic resource by metabolically engineered Enterobacter cloacae . Green Chem. 18, 1560–1570 (2016).
Cambon, B., Monteil, V., Remize, F., Camarasa, C. & Dequin, S. Effects of GPD1 overexpression in Saccharomyces cerevisiae commercial wine yeast strains lacking ALD6 genes. Appl. Environ. Microbiol. 72, 4688–4694 (2006).
Kim, S. & Hahn, J. S. Efficient production of 2,3-butanediol in Saccharomyces cerevisiae by eliminating ethanol and glycerol production and redox rebalancing. Metab. Eng. 31, 94–101 (2015).
Kim, S. J., Seo, S. O., Jin, Y. S. & Seo, J. H. Production of 2,3-butanediol by engineered Saccharomyces cerevisiae . Bioresour. Technol . 146, 274–281 (2013).
Lian, J., Chao, R. & Zhao, H. Metabolic engineering of a Saccharomyces cerevisiae strain capable of simultaneously utilizing glucose and galactose to produce enantiopure (2R, 3R)-butanediol. Metab. Eng. 23, 92–99 (2014).
Kim, S., Lee, K., Bae, S. J. & Hahn, J. S. Promoters inducible by aromatic amino acids and g-aminobutyrate (GABA) for metabolic engineering applications in Saccharomyces cerevisiae . Appl. Microbiol. Biotechnol. 99, 2705–2714 (2015).
Ansell, R., Granath, K., Hohmann, S., Thevelein, J. M. & Adler, L. The two isoenzymes for yeast NAD+-dependent glycerol 3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation. EMBO J. 16, 2179–2187 (1997).
Gonzalez, E. et al. Role of Saccharomyces cerevisiae oxidoreductases Bdh1p and Ara1p in the metabolism of acetoin and 2,3-butanediol. Appl. Environ. Microbiol. 76, 670–679 (2010).
Chen, X. L., Li, S. B. & Liu, L. M. Engineering redox balance through cofactor systems. Trends Biotechnol. 32, 337–343 (2014).
Forster, J., Famili, I., Fu, P., Palsson, B. O. & Nielsen, J. Genome-scale reconstruction of the Saccharomyces cerevisiae metabolic network. Genome Res. 13, 244–253 (2003).
Heux, S., Cachon, R. & Dequin, S. Cofactor engineering in Saccharomyces cerevisiae: Expression of a H2O-forming NADH oxidase and impact on redox metabolism. Metab. Eng. 8, 303–314 (2006).
Hejazi, A. & Falkiner, F. R. Serratia marcescens . J. Med. Microbiol. 46, 903–912 (1997).
Gaston, M. A. Enterobacter: an emerging nosocomial pathogen. J. Hosp. Infect. 11, 197–208 (1988).
Dai, J. Y., Cheng, L., He, Q. F. & Xiu, Z. L. High acetoin production by a newly isolated marine Bacillus subtilis strain with low requirement of oxygen supply. Process Biochem. 50, 1730–1734 (2015).
Zhang, X. et al. Two-stage pH control strategy based on the pH preference of acetoin reductase regulates acetoin and 2,3-butanediol distribution in Bacillus subtilis . PLoS One 9, e91187 (2014).
Luo, Q., Wu, J. & Wu, M. Enhanced acetoin production by Bacillus amyloliquefaciens through improved acetoin tolerance. Process Biochem. 49, 1223–1230 (2014).
Zhang, L., Chen, S., Xie, H., Tian, Y. & Hu, K. Efficient acetoin production by optimization of medium components and oxygen supply control using a newly isolated Paenibacillus polymyxa CS107. J. Chem. Technol. Biotechnol. 87, 1551–1557 (2012).
Mumberg, D., Muller, R. & Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 156, 119–122 (1995).
Acknowledgements
This work was supported by the National Research Foundation of Korea (NRF) Grant funded by the Korean Government (NRF-2015R1A2A2A01005429).
Author information
Authors and Affiliations
Contributions
S.-J.B. and S.K. performed the experiments and analyzed the data. S.-J.B., S.K. and J.-S.H. designed the experiments and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Bae, SJ., Kim, S. & Hahn, JS. Efficient production of acetoin in Saccharomyces cerevisiae by disruption of 2,3-butanediol dehydrogenase and expression of NADH oxidase. Sci Rep 6, 27667 (2016). https://doi.org/10.1038/srep27667
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep27667
This article is cited by
-
Sugarcane bagasse: an important lignocellulosic substrate for production of enzymes and biofuels
Biomass Conversion and Biorefinery (2024)
-
Systems metabolic engineering of microorganisms for food and cosmetics production
Nature Reviews Bioengineering (2023)
-
Efficient acetoin production from pyruvate by engineered Halomonas bluephagenesis whole-cell biocatalysis
Frontiers of Chemical Science and Engineering (2023)
-
Efficient production of acetoin from lactate by engineered Escherichia coli whole-cell biocatalyst
Applied Microbiology and Biotechnology (2023)
-
Acetoin production from lignocellulosic biomass hydrolysates with a modular metabolic engineering system in Bacillus subtilis
Biotechnology for Biofuels and Bioproducts (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
