
Abstract
In this study, we describe a web-based resource, developed for assisting the scientific community in designing an effective therapeutics against the Ebola virus. Firstly, we predicted and identified experimentally validated epitopes in each of the antigens/proteins of the five known ebolaviruses. Secondly, we generated all the possible overlapping 9mer peptides from the proteins of ebolaviruses. Thirdly, conserved peptides across all the five ebolaviruses (four human pathogenic species) with no identical sequence in the human proteome, based on 1000 Genomes project, were identified. Finally, we identified peptide or epitope-based vaccine candidates that could activate both the B- and T-cell arms of the immune system. In addition, we also identified efficacious siRNAs against the mRNA transcriptome (absent in human transcriptome) of all the five ebolaviruses. It was observed that three species can potentially be targeted by a single siRNA (19mer) and 75 siRNAs can potentially target at least two species. A web server, EbolaVCR, has been developed that incorporates all the above information and useful computational tools (http://crdd.osdd.net/oscadd/ebola/).
Similar content being viewed by others

Introduction
The Ebola virus (EBOV), came into the picture with the outbreak in 1976 in Sudan and subsequently in Zaire, where it was initially detected near the Ebola River (Zaire species) from which it received its current name1. It can infect both animals and humans. The medium of transmission of this virus is mainly body fluids such as milk, saliva, semen, urine, etc. The Ebola Virus Disease (EVD) is characterized by abrupt fever, stomach pain, headache, vomiting, and diarrhea2. Because the EVD has overlapping symptoms with other diseases such as malaria, flu and typhoid, it makes it all the more complicated to diagnose the infection from the symptoms3. The onset of the disease is confirmed using ELISA, PCR and virus isolation test4.
Once the disease has been diagnosed, the next challenge is the treatment and tackling symptoms by providing intravenous fluids for maintaining electrolytes, oxygen status and blood pressure5. Even so, there are limited options to treat this disease directly. Recently, ZMapp was invented against the Ebola virus (EBOV), which is a combination of 3 monoclonal antibodies6,7. Although it has not been tested in humans, successful animal studies have been carried out8. To the best of the authors’ knowledge, there is no known approved vaccine against this disease available at the moment9.
However, there are two promising vaccine candidates that are currently in phase-I of clinical trials10. The first candidate (ChAd3-ZEBOV) has been developed by GlaxoSmithKline (GSK) and the National Institute of Allergy and Infectious Diseases (NIAID)11. This vaccine comprises a glycoprotein gene from EBOV and Sudan Ebola Virus (SUDV) under the control of chimpanzee adenovirus 3. The second potential vaccine is rVSV-ZEBOV, developed by NewLink Genetics and Merck Vaccines USA in collaboration with the Public Health Agency of Canada that uses attenuated virus with glycoprotein of EBOV12. The rVSV-ZEBOV was found to be safe and effective in clinical trial results13.
In the past, RNAi has been proven to be a promising tool for the treatment of many viral diseases14 and has helped in circumventing viral infections15. A siRNA targeting the Zaire Ebola virus RNA polymerase L protein formulated in stable nucleic acid-lipid particles (SNALPs) showed complete protection in guinea pigs16. In a separate study, the authors have used a combination of siRNAs targeting L, VP24 and VP35 formulated in SNALPs and observed protection in two-third of the rhesus monkeys and in all macaques who were given seven post exposure treatments17. TKM-100802 is a set of small RNA molecules encapsulated in lipid nanoparticles that target the vital viral proteins (VP24, VP35, and L) and cleave viral RNA in the cells and prevent its multiplication16,17. A recent study shows that short interfering RNAs (siRNAs) packaged into lipid nanoparticles are able to protect all the rhesus monkeys under experimental design against Makona strain outbreak of Ebola virus18.
TKM-Ebola-Guinea drug trial is an outcome of the collaborative efforts of Tekmira (a Canadian company) and Oxford University; unfortunately, the clinical trial was halted recently as therapeutic benefits to the patients were not observed after reaching the predefined endpoint19.
Hence, there is a need to design computer-aided subunit vaccines and siRNA-based therapy against EVD. Fortunately, the genomes of the five ebolaviruses have been sequenced and annotated20,21. Thus, it is possible to develop computational approaches for designing potential therapy against the Ebola virus. In this study, we aim to create a web-based resource that provides information required for developing effective drugs or vaccines against this virus.
Methods
Sequence of Genomes
We downloaded the protein, and mRNA sequences of five ebolaviruses: Bundibugyo ebolavirus (BDBV) (Accession Number FJ217161), Reston ebolavirus (RESTV) strain Pennsylvania (Accession number AF522874), Sudan ebolavirus (SUDV) strain Gulu (Accession Number AF086833), Tai Forest ebolavirus (TAFV) (Accession Number FJ217162), and Ebola virus - Mayinga, Zaire, 1976 (EBOV-May) (Accession Number AF086833) from the NCBI22,23. These genomes have sizes between 18–19 kb with 40–42% GC content. There are seven genes present in EBOV with two variants of glycoproteins (GP) gene in RESTV and SUDV, while three splice variants have been annotated in other species. Following are the splice variants of GP; i) GP is the spike protein, ii) sGP: (soluble GP) is the default 7A variant and iii) ssGP: (small soluble GP) is a -1A(6A) variant. The alignment file for each protein/mRNA across the different species was generated using ClustalW24.
Viral-specific conserved patterns
In order to investigate virus-specific conserved patterns, overlapping patterns of 9 amino acids (9mer) were generated, from proteins of above-mentioned five ebolaviruses. There were 43 proteins in total (including different splice variants of GP gene) from the five ebolaviruses. Further, all the possible 9mer peptides from each protein were predicted for their immunological potential. One of the challenges in vaccine development is to avoid self-tolerance as the body’s immune system rarely acts against self-antigens. Thus, all those 9mers that are available in the human body (or human genome) were removed. In order to achieve this goal, we removed all those viral patterns that are 100% identical to any pattern found in human thousand proteomes. Human thousand proteomes were created by using 1000 Genomes data and translating it to proteins sequences25,26. This led us to the 9mer peptide/pattern that is found in this virus but absent in human proteomes. In order to develop an effective vaccine against a wide range of species, it is important to identify conserved patterns across the different species of Ebola. Therefore, this study involves the identification of unique patterns of 9mer peptides along with the frequency of their occurrence across different ebolaviruses. Also, a high priority was given to a pattern, which was common in the most number of ebolaviruses, which led to patterns/peptides of size of 9 residues that were conserved among ebolaviruses.
Pipeline for epitope prediction
After the exploration of 9mer peptides which were found to be conserved in different ebolaviruses but absent in human proteomes, our next goal was to identify peptides that could activate the human immune system in order to generate memory cells against Ebola virus. In order to understand the immunomodulatory or stimulatory effect of these peptides, we computed different types of epitopes as described below:
B-cell epitope prediction
In this study, we have used the recently developed method LBtope27 for predicting linear B-cell epitopes among the aforementioned conserved patterns. LBtope is a highly accurate method trained on a large dataset that contains experimentally validated B-cell epitopes and non-epitopes. We applied the LBtope model with default threshold of 60% probability cut-off. Similarly, conformational B-cell epitopes have been predicted in the above conserved peptides using CBTOPE28. CBTOPE is a unique method that predicts conformational B-cell epitopes in an antigen from its amino acid sequence as opposed to previous methods, which required the structure of the antigen to predict the same. The CBTOPE results were obtained on the default threshold of −0.3 cut-off and implemented on our website.
MHC alleles binding peptides
The major histocompatibility complex (MHC) is a set of cell surface molecules that plays a vital role in the human immune system. The major role of the MHC is to bind to antigenic regions (or peptides) and present these peptides on the cell surface, where appropriate T-cells recognize these peptides. Thus, it is important to predict MHC binders, as these binders have the ability to activate the T-cells of the immune system. In this study, we have used ProPred1 with top 4% cutoff (default) for predicting MHC Class I binders29. ProPred1 is a matrix-based program, which predicts the peptide binding potential for 47 MHC Class I alleles. These binders may activate cytotoxic T-lymphocyte (CTL), so we may call these binders as potential CTL epitopes, which were predicted using CTLPred with default parameters. Similarly, to predict potential T-helper epitopes, we predicted MHC Class II binders in the above conserved peptides. The prediction of MHC Class II binders was performed using ProPred30 with top 3% threshold as suggested by the authors of ProPred. It is a quantitative/virtual matrix-based method that allows the users to predict promiscuous MHC Class II binders for 51 alleles.
T-cell response prediction
In addition to the prediction of MHC binders which are potential T-cell epitopes, we also predicted CTL epitopes using a direct method, CTLPred31. CTLPred is a direct method that predicts T-cell epitopes (CTL) from a primary sequence of the antigen instead of using the intermediate step where MHC Class I binders are predicted. CTLPred has a number of modules (e.g., ANN, SVM) for predicting CTL epitopes; in this study we have used SVM-based module with default parameters. In the past, numerous methods have been developed for predicting MHC Class II binders but none of them has been able to predict the type of interleukins released by these binders. In this study, we have used a method, IFNepitope with a default threshold of 0.0 to predict the antigenic region or MHC binders that can activate Th1 cells (T-helper cell type I)32 responsible for releasing interferon-gamma (IFN-γ). In brief, we predicted IFN-γ inducing peptides in Ebola antigens using IFNepitope. Similarly, we also predicted antigenic regions that can activate Th2 cells responsible for releasing of cytokine, interleukin-4 (IL4). In this study, we used IL4pred, at a default threshold of 0.2, for predicting the IL4-inducing antigenic regions or peptides in Ebola antigens33.
RNA therapeutics
In order to design siRNA-based therapeutics against the Ebola virus, we generated 19mer oligonucleotide from the corresponding mRNA sequences, as this length is sufficient to silence the cognate mRNA target sequence. In literature, 19mer is considered the core length of siRNA duplex (apart from 2 nucleotide overhang) that is used for RNAi-based therapy to silence a target transcript. Most of the prediction methods are developed based on 19mer length of siRNA sequence34,35,36.
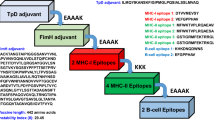
First we generated 19mer overlapping oligonucleotides in each gene of the above-mentioned five ebolaviruses. Secondly, we removed all the oligonucleotides present in human genes sequenced so far, using human reference mRNA. Thirdly, we identified oligonucleotides conserved in different species of the virus. Finally, we got 19mer oligonucleotides conserved in the viral species but absent in the human genome. In order to identify which oligonucleotide could serve as an efficient siRNA against the Ebola virus, we predicted the efficacy of each oligonucleotide (siRNA) using DesiRm software with default parameters34. The final set of 19mers was further subgrouped gene-wise and in each subgroup, we separated the highly potent siRNAs (having efficacy greater than 0.80 value). The overall architecture of the EbolaVCR developed in this study has been shown in Fig. 1.

Figures inside this figure are created with the help of ScienceSlides software (http://www.visiscience.com/).
Tool section
Phylogenetic trees
MEGA software version 5.037 has been used for creating the phylogenetic tree from protein sequences of Ebola virus. In order to generate multiple sequence alignment of proteins we used the software ClustalW24. These alignments were used to construct the tree using Maximum-Likelihood algorithm where branch lengths were visualized from the given options.
Protein structure of Ebola antigens
In order to understand the function of Ebola proteins, we have predicted the tertiary structure of Ebola proteins. To assist the scientific community, we predicted tertiary structures of viral proteins using I-TASSER suite38. The templates for structure prediction were derived using Hhbuilt suite with default parameters. All the predicted protein structures for 38 proteins (except for L protein) are available from our web resource. Further, visualization tools have been integrated to visualize these structures. In addition to predicted structures, we have also provided the structure of Ebola virus proteins present in protein data bank (PDB). We believe these structures will be helpful in computer-aided drug design.
Results
Subunit vaccine against Ebola virus
9mer peptides
In our study, we have generated all the possible overlapping 9mer peptides for each of the ebolavirus. We have removed the redundant as well as peptides found in human reference proteome (Table 1). Further, the peptides present in 1000 Genomes-based human proteome were also filtered to avoid any cross presentation with any individual. We have observed 2 peptides from RESTV and single peptide from SUDV, which were shared with human reference proteome. In addition to that, we have also observed a peptide from EBOV that was not present in the human reference proteome but present among the peptides of 1000 Genomes-based human proteome. The protein-wise distribution of peptides from different ebolaviruses is shown in supplementary files (Figure S1).
Vaccine candidates
The conservation in a peptide among the different ebolaviruses is helpful for its applicability in a broad spectrum. Therefore, we have extracted conserved peptides (9mer), and found that 629 peptides were shared among all the five ebolaviruses in our study (Figure S2) and 812 peptides were conserved among four human pathogenic ebolaviruses (excluding RESTV) (Fig. 2, Table 2). These common peptides, shared among different ebolaviruses, have the potential to design vaccine for all types of ebolaviruses. We computed protein-wise distribution of conserved peptides among different species of Ebola virus (Figure S3, Table S1). In order to identify conserved regions, we computed multiple sequence alignment of each protein of virus using ClustalW (Figures S4 and S5). We also predicted immunogenicity of these peptides, particularly of conserved peptides, using our immuno-pipeline. In our analysis, we have not found any of the conserved peptides with B-cell epitopes and promiscuous MHC binders. There were 21 peptides that were common among all the four human pathogenic ebolaviruses (excluding RESTV, not pathogenic to human). These peptides are highly promiscuous vaccine candidates (Table 2). We further analyzed 14 peptides that can activate most of the arms of the immune system (Table 3). Out of these 14 peptides, 2 were conserved in three species and three peptides were conserved in at least two ebolaviruses.

The Venn Diagram for five sets of peptides from four human pathogenic ebolaviruses.
siRNA-based therapy
Alignment of mRNA sequences
We performed the multiple sequence alignment (MSA) for genes across all the five ebolaviruses in order to identify the conserved regions in different genes. We were unable to locate a single 19mer oligonucleotide, common in all the five ebolaviruses. We got a stretch of maximum length of nine nucleotides across five ebolaviruses. In this study we used ClustalW software24 for MSA and Jalview39 visualization of alignment (Figure S6).
Potential siRNA suppressors
The specific 19mer oligonucleotides were generated in all the five ebolaviruses. Figure 3 shows the architecture of the Ebola RNAi pipeline, which presents the strategy adopted to identify potential therapeutic siRNAs. First, we computed the efficacy of each 19mer oligonucleotide using desiRm software at default threshold value of 0. Next, we identified 19mers, which were present in human genome in order to eliminate off-target interaction. Finally, we selected all those siRNAs, which may silence targets gene with efficacy more than a value of 0.80. We wanted to identify promiscuous siRNA that could target all five ebolaviruses. Unfortunately, we did not find a single common 19mer oligonucleotide in any of the five ebolaviruses (Figure S7). The distribution of siRNAs selected against Ebola across the five ebolaviruses at gene level has been shown in Fig. 4. The distribution of potential siRNAs against each target gene in ebolaviruses is shown in Table 4 and the number of siRNAs targeting common ebolaviruses is enlisted in Supplementary Table S2. Our server shows potential siRNAs against target genes in the form of tables with the Circos plots40 for visualization. Users can get detailed information like strain ID, gene, siRNA sequence, cognate mRNA sequence, starting position and efficacy from our website. Users can sort the table according to a position or efficacy to get the most efficacious siRNA along with other details. The detailed gene-wise distribution of siRNA from different ebolaviruses is shown in supplementary files (Figure S8).

RNAi pipeline of EbolaVCR and number of potential therapeutic siRNA predicted for each ebolavirus.

(A) Gene-wise distribution of siRNA across five ebolaviruses. (B) Shared siRNA across four human pathogenic ebolaviruses.
Integration of Computational Tools
In this web-based platform, we have integrated a number of computational tools to provide a comprehensive service to the scientific community working on Ebola virus. These tools will be very useful in designing therapeutics against new ebolavirus or drug resistant ebolavirus. Following is the brief description of modules integrated in EbolaVCR.
Mapping of experimentally validated epitopes
The Immune Epitope Database (IEDB) is the largest database of experimentally characterized antigenic regions or epitopes that may modulate immune system. In summary, IEDB maintains experimentally validated epitopes. In order to identify antigenic regions that have already been characterised in the past, we mapped experimentally proven IEDB epitopes on Ebola antigens. Here, we have extracted the epitopes using the filter of source organism as ebolavirus or name of any ebolavirus. We have linked these epitopes and their source organism(s) to the original source page of IEDB. The source molecule of these epitopes is linked to NCBI for detailed description. We have also provided the facility to map the peptides from the user sequence to IEDB, MHCBN and BCIPEP databases41,42,43.
Phylogenetic trees
It is important to understand the similarity among ebolaviruses or evolution of a species. In order to understand evolution in ebolavirus, we generated phylogenetic trees, based on similarity in their protein sequences. In this study, MEGA software has been used for generating phylogenetic trees using default parameters37. The trees are available at our web resource. In our analysis of phylogenetic trees for each gene, we have observed the evolution of ebolaviruses. We found that RESTV and SUDV have been coevolved from the same phylogenetic node, and BDBV and TAFV are closely related, while EBOV evolved from a distant node of ancestry.
Discussion
Promising vaccine epitopes
The vaccine against the Ebola virus is the need of the hour6,9,10. In this study, we made an attempt to analyze the conserved epitopes so that the resulting vaccine covers a broad range of species, sequenced till date. Besides conservation, immune response is the next challenge to be addressed by the vaccine candidates. We have predicted diverse kinds of immune responses for better efficacy of the candidates. In the past, computational studies have been carried out for designing vaccines against viruses44,45. But these studies were limited in terms of the number of proteins covered and the types of immune responses considered46,47,48. We have observed the peptides (9mers) that are capable of stimulating different kinds of immunity. In the current study, we have uncovered 123 conserved peptides among all the human pathogenic ebolavirus that can activate cytotoxic T lymphocytes (CTL). In addition to CTL activation, we have also identified 42 peptides that can activate the T helper response after binding with MHC class II. We have also eliminated the peptides that could share sequence identity with the human proteome to avoid any cross-reaction of vaccine candidates with human proteins. We are hopeful that these vaccine candidates will provide better immunity and lead to a successful vaccine in the future. We have also discovered 45 conserved peptides that were able to trigger B-cell response to generate antibody among all the human pathogenic subtypes of the Ebola virus. These peptides could serve in the diagnosis of the Ebola virus.
We have also examined experimentally validated epitopes available in the Immune Epitope Database (IEDB). It was observed that 1138 epitopes, covered in our study, out of 1513 peptides were available in IEDB. This demonstrates the validity of our predictions. In addition, we suggested 16 epitopes in our study, one of which has been validated experimentally to bind with 2 MHC alleles (HLA*24-02, HLA-B*07:02). The peptide (IPVYQVNNL) was identified and submitted to IEDB (http://www.iedb.org/epId/91517)49 in 2009 by Kent J. Weinhold, Georgia D. Tomaras, Yongting Cai, Kelly Plonk, Scott Pruitt and Smita K. Nair.
siRNA therapy
The most potential siRNAs against every gene are reported in this study. We have predicted the efficacy of exclusive siRNAs against all the genes of known ebolaviruses and found that none of the siRNAs is present against all the five ebolaviruses. There are 75 siRNAs, which are shared by any two species, and 1 siRNA is shared by three species (BDBV, EBOV and RESTV). In this study, we used a published siRNA efficacy predictor named ‘desiRm’, developed by our group. One may question the use of desiRm software when a large number of existing software packages for predicting the efficacy of siRNA are available. We used it because its standalone version allowed us to analyze a large number of 19mer oligonucleotides. Further, this algorithm has been shown to have comparable performance with the other methods34. Thus, selection is purely based on availability and not on any other criteria, like performance.
In this study, almost all the software used for predicting potential candidates for vaccine or siRNA therapeutics, were developed by our group. Some of these are extensively used software (e.g., ProPred, ProPred1 and CTLPred) developed in our laboratory. Some of the software packages are unique and not available from any other group, like IL4pred and IFNepitope. In conclusion, we have tried our best to use the most appropriate software packages to generate the above resources.
Additional Information
How to cite this article: Dhanda, S. K. et al. A web-based resource for designing therapeutics against Ebola Virus. Sci. Rep. 6, 24782; doi: 10.1038/srep24782 (2016).
References
Feldmann, H. & Geisbert, T. W. Ebola haemorrhagic fever. Lancet 377, 849–62 (2011).
Bellan, S. E., Pulliam, J. R. C., Dushoff, J. & Meyers, L. A. Ebola control: effect of asymptomatic infection and acquired immunity. Lancet 384, 1499–500 (2014).
Mishra, B. The threat of Ebola: An update. Indian J. Med. Microbiol. 32, 364–70 (2014).
Brizendine, K. D. Ebola virus: Questions, answers, and more questions. Cleve. Clin. J. Med. 81, 729–35 (2014).
Wong, G., Qiu, X., Olinger, G. G. & Kobinger, G. P. Post-exposure therapy of filovirus infections. Trends Microbiol. 22, 456–63 (2014).
Strauss, S. Biotech drugs too little, too late for Ebola outbreak. Nat. Biotechnol. 32, 849–50 (2014).
Qiu, X. et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 514, 47–53 (2014).
Enserink, M. Infectious diseases. Debate erupts on ‘repurposed’ drugs for Ebola. Science 345, 718–9 (2014).
Cohen, J. Infectious disease. Ebola vaccine: little and late. Science 345, 1441–2 (2014).
Sullivan, N. J., Sanchez, A., Rollin, P. E., Yang, Z. Y. & Nabel, G. J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 408, 605–9 (2000).
Martin, J. E. et al. A DNA vaccine for Ebola virus is safe and immunogenic in a phase I clinical trial. Clin. Vaccine Immunol. 13, 1267–77 (2006).
Marzi, A. et al. Vesicular stomatitis virus-based Ebola vaccines with improved cross-protective efficacy. J. Infect. Dis. 204 Suppl, S1066–74 (2011).
Henao-Restrepo, A. M. et al. Efficacy and effectiveness of an rVSV-vectored vaccine expressing Ebola surface glycoprotein: interim results from the Guinea ring vaccination cluster-randomised trial. Lancet 386, 857–866 (2015).
Zhang, Y. et al. Silencing SARS-CoV Spike protein expression in cultured cells by RNA interference. FEBS Lett. 560, 141–6 (2004).
Dykxhoorn, D. M. & Lieberman, J. Silencing viral infection. PLos Med. 3, e242 (2006).
Geisbert, T. W. et al. Postexposure protection of guinea pigs against a lethal ebola virus challenge is conferred by RNA interference. J. Infect. Dis. 193, 1650–7 (2006).
Geisbert, T. W. et al. Postexposure protection of non-human primates against a lethal Ebola virus challenge with RNA interference: a proof-of-concept study. Lancet 375, 1896–905 (2010).
Thi, E. P. et al. Lipid nanoparticle siRNA treatment of Ebola-virus-Makona-infected nonhuman primates. Nature 521, 362–5 (2015).
Update on Ebola treatment trial in Sierra Leone. (19th June 2015) Available at: http://www.wellcome.ac.uk/News/2015/WTP059401.htm. (Accessed: 18th December 2015).
Kuhn, J. H. et al. Evaluation of perceived threat differences posed by filovirus variants. Biosecur. Bioterror. 9, 361–71 (2011).
Gire, S. K. et al. Genomic surveillance elucidates Ebola virus origin and transmission during the 2014 outbreak. Science. 345, 1369–1372 (2014).
Kuhn, J. H. et al. Virus nomenclature below the species level: a standardized nomenclature for natural variants of viruses assigned to the family Filoviridae. Arch. Virol. 158, 301–11 (2013).
Kuhn, J. H. & Jahrling, P. B. Clarification and guidance on the proper usage of virus and virus species names. Arch. Virol. 155, 445–53 (2010).
Larkin, M. A. et al. Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–8 (2007).
Abecasis, G. R. et al. A map of human genome variation from population-scale sequencing. Nature 467, 1061–73 (2010).
Abecasis, G. R. et al. An integrated map of genetic variation from 1,092 human genomes. Nature 491, 56–65 (2012).
Singh, H., Ansari, H. R. & Raghava, G. P. S. Improved method for linear B-cell epitope prediction using antigen’s primary sequence. PLos One 8, e62216 (2013).
Ansari, H. R. & Raghava, G. P. Identification of conformational B-cell Epitopes in an antigen from its primary sequence. Immunome Res. 6, 6 (2010).
Singh, H. & Raghava, G. P. S. ProPred1: prediction of promiscuous MHC Class-I binding sites. Bioinformatics 19, 1009–14 (2003).
Singh, H. & Raghava, G. P. ProPred: prediction of HLA-DR binding sites. Bioinformatics 17, 1236–7 (2001).
Bhasin, M. & Raghava, G. P. S. Prediction of CTL epitopes using QM, SVM and ANN techniques. Vaccine 22, 3195–204 (2004).
Dhanda, S. K., Vir, P. & Raghava, G. P. S. Designing of interferon-gamma inducing MHC class-II binders. Biol. Direct 8, 30 (2013).
Dhanda, S. K., Gupta, S., Vir, P. & Raghava, G. P. S. Prediction of IL4 inducing peptides. Clin. Dev. Immunol. 2013, 263952 (2013).
Ahmed, F. & Raghava, G. P. S. Designing of highly effective complementary and mismatch siRNAs for silencing a gene. PLos One 6, e23443 (2011).
Katoh, T. & Suzuki, T. Specific residues at every third position of siRNA shape its efficient RNAi activity. Nucleic Acids Res. 35, e27 (2007).
Shabalina, S. A., Spiridonov, A. N. & Ogurtsov, A. Y. Computational models with thermodynamic and composition features improve siRNA design. BMC Bioinformatics 7, 65 (2006).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–9 (2011).
Roy, A., Kucukural, A. & Zhang, Y. I-TASSER: a unified platform for automated protein structure and function prediction. Nat. Protoc. 5, 725–38 (2010).
Waterhouse, A. M., Procter, J. B., Martin, D. M. A., Clamp, M. & Barton, G. J. Jalview Version 2–a multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189–91 (2009).
Krzywinski, M. et al. Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–45 (2009).
Bhasin, M., Singh, H. & Raghava, G. P. S. MHCBN: a comprehensive database of MHC binding and non-binding peptides. Bioinformatics 19, 665–6 (2003).
Saha, S., Bhasin, M. & Raghava, G. P. S. Bcipep: a database of B-cell epitopes. BMC Genomics 6, 79 (2005).
Vita, R. et al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 43, D405–D412 (2014).
Becker, Y. Computer simulations of proteolysis of Marburg and Ebola-Zaire filovirus coded proteins to generate nonapeptides with motifs of known HLA class I haplotypes and detection of antigenic domains in the viral glycoproteins. Virus Genes 13, 189–201 (1996).
He, L. et al. Integrated assessment of predicted MHC binding and cross-conservation with self reveals patterns of viral camouflage. BMC Bioinformatics 15 Suppl 4, S1 (2014).
Simmons, G. et al. Identification of murine T-cell epitopes in Ebola virus nucleoprotein. Virology 318, 224–30 (2004).
Wu, S. et al. Prediction and identification of mouse cytotoxic T lymphocyte epitopes in Ebola virus glycoproteins. Virol. J. 9, 111 (2012).
Sundar, K., Boesen, A. & Coico, R. Computational prediction and identification of HLA-A2.1-specific Ebola virus CTL epitopes. Virology 360, 257–63 (2007).
IPVYQVNNL. (2014) Available at: http://www.iedb.org/epId/91517. (Accessed: 18th December 2015).
Acknowledgements
The authors would like to acknowledge funding from the Council of Scientific and Industrial Research through the OSDD consortium and GENESIS projects. We are also thankful to the Department of Biotechnology, Government of India for funding the project through BTISNET. SKD also acknowledges the Indian Council of Medical Research for fellowship. SKB and GPSR acknowledge the J. C. Bose National Fellowship conferred by the Department of Science and Technology.
Author information
Authors and Affiliations
Contributions
S.K.D. carried out the vaccine analysis part of the project, while the siRNA therapeutics analysis was done by K.C. and S.G. created the 9mer peptides from human 1000 Genomes and helped S.K.D. in creating immunoinformatics pipeline. S.K.B. and G.P.S.R. conceived and supervised the project.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Dhanda, S., Chaudhary, K., Gupta, S. et al. A web-based resource for designing therapeutics against Ebola Virus. Sci Rep 6, 24782 (2016). https://doi.org/10.1038/srep24782
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep24782
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
