
Abstract
Small interfering RNA (siRNA) technology has vast potential for functional genomics and development of therapeutics. However, it faces many obstacles predominantly instability of siRNAs due to nuclease digestion and subsequently biologically short half-life. Chemical modifications in siRNAs provide means to overcome these shortcomings and improve their stability and potency. Despite enormous utility bioinformatics resource of these chemically modified siRNAs (cm-siRNAs) is lacking. Therefore, we have developed siRNAmod, a specialized databank for chemically modified siRNAs. Currently, our repository contains a total of 4894 chemically modified-siRNA sequences, comprising 128 unique chemical modifications on different positions with various permutations and combinations. It incorporates important information on siRNA sequence, chemical modification, their number and respective position, structure, simplified molecular input line entry system canonical (SMILES), efficacy of modified siRNA, target gene, cell line, experimental methods, reference etc. It is developed and hosted using Linux Apache MySQL PHP (LAMP) software bundle. Standard user-friendly browse, search facility and analysis tools are also integrated. It would assist in understanding the effect of chemical modifications and further development of stable and efficacious siRNAs for research as well as therapeutics. siRNAmod is freely available at: http://crdd.osdd.net/servers/sirnamod.
Similar content being viewed by others
Introduction
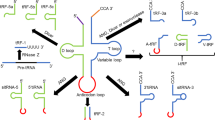
RNA interference (RNAi) phenomenon was described by A. Fire and coworkers in Caenorhabditis elegans1. They injected double stranded RNA that potentially and specifically interfere with the endogenous gene at mRNA level leading to its silencing1. RNA silencing pathway utilizes dicer to process long double stranded RNA (dsRNA) to 19–21 nucleotide small interfering RNAs (siRNAs) with 2-nucleotide unphosphorylated 3′ overhangs2. RNA induced silencing complex (RISC) incorporates siRNA antisense strand, resulting in cleavage of cognate mRNA target3 as shown in Fig. 1.

Cartoon representation of RNAi mechanism, siRNA molecular structure and its entry in RNAi pathway.
siRNAs are explored extensively in gene silencing experiments and therapeutics development4. However, their therapeutic usage is hampered due to many factors e.g. susceptibility to ribonucleases digestion, short biological half-life, lack of proper delivery vehicle, cellular uptake, immune-stimulatory effect, non-specific gene targeting and toxicity5,6. These limitations are due to the inherent physicochemical properties of siRNAs that includes short and stiff structure, high charge density (poly-anionic), hydrophilic and liability to nuclease cleavage5,6. To mitigate these restraints, chemical modifications in siRNAs have been substantially investigated7.
Synthetic oligonucleotides were used in the late 1970’s for gene inhibition known as antisense technology8. Modified nucleoside phosphoramidites are used in solid phase chemical synthesis of siRNA which allow site-specific incorporation of chemically modified nucleotide moiety at specific positions within the siRNA9. Chemical modifications on siRNA are categorized based on component of nucleotide modified as ribose, base or phosphate and change in properties like biological activity, thermodynamic stability and nuclease resistance7. Chemical modifications on sugar moiety e.g. locked nucleic acids (LNA), unlocked nucleic acids (UNA)10,11, 2′-deoxy, 2′-O-methyl12, 2′-fluoro, 2′-methoxyethyl, 2′-aminoethyl, were tested for RNAi activity13,14. In LNA, methylene bridge is created between 2′-oxygen and 4′-carbon to increase the RNAi activity favorably15,16 and also improves nuclease resistance17. UNA are open ring derivatives of ribose without C-2′ and C-3′ carbon-carbon bond but structurally mimic the RNA when incorporated into duplex18. Ribose 2′-OH and phosphorothioate modification increases stability of siRNA19,20. Besides the 2′-OH was not found to be essential for RNAi activity21. However, fluorine modification at 2′-OH on different nucleotide positions of siRNA maintains silencing potency as well as increases thermal stability22,23. Further, researchers carried out various chemical modifications in different combinations and tested them for RNAi activity13,14. Also 2′-O-aminoethyl addition on passenger strand (sense strand) at 3′-end enhanced RNAi activity of siRNA duplex due to asymmetry in thermodynamic stability24.
Base modifications have been used in various studies e.g. hypoxanthine, 2,4-di-fluorotoluene, dihydrouridine, 2′-thiouridine, pseudouridine12,25,26 etc. Modification in this component can contribute towards better understanding of off-target effects as well as gene silencing mechanism27. Besides, phosphate modifications were also employed in different studies to enhance the properties of siRNAs28,29. These modifications include phosphorothioate, boranophosphate, peptide nucleic acids (PNA)28,29 etc. For instance phosphorothioate enhances the stability, uptake and biodistribition of the siRNA30,31. Large-scale screens and high throughput platforms using different permutation and combinations of chemical modifications were explored to check their effect24,32. Native (un-modified) siRNA show immunological activation in vitro to various degrees, while certain chemical modifications (2′-O-methyl) of siRNA avoid immunological effects33. Chemical modifications may be useful to overcome the obstacle of delivery of siRNAs without compromising their gene silencing activity5.
As discussed, next generation siRNA technology will be defined by the chemistry of modifications on siRNAs to greater extent. But we currently lack the platform to explore the complete spectrum of chemical modifications synthesized and plotted with relation to siRNAs so far. To describe the rules governing the performance of these cm-siRNAs, we need to have birds-eye view on the entire set of chemical modifications. Furthermore, positions of modifications on sense and antisense strands, modified nucleotide component, chemical features like size, functional groups, etc. need to be investigated thoroughly. Other associated experimental details like target gene, transfection method, cell line or organism used are very important and needs to be considered before further investigation. Additionally, the data needs to be present in the format that can be easily retrieved and may be utilized for further computational analysis.
Many repositories for native (unmodified) siRNA are available in the literature like siRecords34, siRNAdb35, human specific siRNA database (HuSiDa)36, viral siRNAs (VIRsiRNAdb)37 and HIV specific siRNA (HIVsirDB)38. Also for naturally occurring modified RNA, there is a database namely RNAMDB39. The latter provides comprehensive listing of the post transcriptionally modified nucleotides occurring naturally as in messenger RNA (mRNA), ribosomal RNA (rRNA) and transfer RNA (tRNA). But no dedicated repository has been developed yet for cm-siRNA despite their usefulness. Therefore, we developed siRNAmod that will serve as the central repository of the experimental records of chemical modification of siRNAs.
Results
Database statistics
siRNAmod contains 4894 experimentally validated cm-siRNA entries. 128 unique chemical modifications incorporated on various components of nucleotides are archived. The overall statistics of the chemical modifications is depicted in Fig. 2. Chemical modifications distribution is illustrated in Fig. 2a. Majority of the reported modifications executed are 2′-O-methyl, locked nucleic acid, 2′-deoxy, 2′-fluoro and unlocked nucleic acid with 17%, 16%, 12%, 7% and 4% respectively. Both 2′-O-methyl and 2′-deoxy are natural RNA modifications. The percentage distribution of natural and non-natural chemical modifications of cm-siRNAs is shown in supplementary Fig. S1. The individual distribution of chemical modifications for antisense and sense siRNA strands is represented in Fig. 2c,d correspondingly. Main modifications are directed on ribose with 63% followed by base and phosphate modifications with 29% and 8% respectively (Fig. 2b). The chemical modifications of the base moiety include hypoxanthine, 2,4-difluorotoluene, boron cluster. While phosphate modifications contain phosphorothioate, boranophosphate, peptide nucleic acid, etc.

Chemical modification statistics pie charts (a) Overall siRNA modifications, (b) nucleotide component based (sugar, base phosphate), (c) Antisense strand modification distribution and (d) Sense strand modification distribution.
Complementarity based sense and antisense distribution of chemical modifications is shown in Fig. 3. Statistical trend of cumulative distribution and combinations of different chemical modifications is represented in supplementary Fig. S3 and S4. Different experimental methods like luciferase assay, RT-PCR, northern blotting etc. were used to validate cm-siRNAs. Most common method to evaluate the efficacy was luciferase assay. The cm-siRNA stability was checked using melting temperature experiments or thermodynamic stability. Transfection methods used on different cell lines mainly utilized lipid based reagents viz lipofectamine and Interfer (Polypus-Transfection). Percentage efficacy ranges from 0–100. However, negative efficacy values were also reported with respect to control taken in a particular experiment. Therefore, for the sake of simplicity, we have reported negative efficacies as zero.

Reverse complement based chemical modification of siRNA position wise on sense and antisense siRNA sequences.
For experimentation, both in vivo and in vitro systems were reported in the literature. Commonly used cell lines were HeLa, HEK 293, Hepa 1–6, Vero, SH-SY5Y etc. Further, in vivo system includes Balb C mice, C57BL/6 mice etc. Pie chart depicting their distribution is presented in supplementary Fig. S5. 128 unique chemical modification structures used to modify the siRNA nucleotides are organized in the form of list with hyperlinks to information of individual modification. Here, we have combined the similar types of modifications into single categories as synonyms to represent the same or similar modifications reported in different studies.
Database browsing
siRNAmod browsing option helps user to surf via these five categories: i) modification in sense strand, ii) position of modification in sense strand, iii) modification in antisense strand, iv) position of modification in antisense strand and v) reference. Modifications that include more than one chemical moiety on same strand are separated by the asterisk (*) symbol. “0” modification in the fields of sense or antisense strand indicates that the modification is present on the other strand.
Simultaneously, facility to navigate 128 chemical modifications is also provided. This offers user an overview of chemical moieties used for modification to modulate siRNA properties. Clicking on the individual modification, structure and related chemical information about the chemical moiety is displayed via jmol40.
Database search
There are two search options provided (i) basic search (ii) structure based search. For basic search, user can enter keyword in query-box and find relevant information. User can choose exact or containing operators. As indicated from the name, “exact” option is the strict search mode, which finds the exact term that user provides while “containing” is lenient search option and provides wider results. The output is displayed in Fig. 4a. On clicking individual Id, user can look for its detail. Further, structure of the chemical modification can be viewed by clicking on the SMILES link as provided (Fig. 4b,c). The result output can be filtered and sorted by clicking or writing on the column headers.

Display of search output with cm-siRNA details (a) Result output of search or browse (b) Details page of individual entry (c) Structure and other chemical information of modification.
For structure drawing and search, MarvinSketch tool is provided. Chemical structures can be hand drawn and used to search for their presence or absence in the database. Step by step instructions for searching database and its output are displayed using screen shots in “How to use” menu on the web server.
Tools
siRNAmod-BLAST provides similarity search option to look for siRNA sequences against the database. This will help the user to know whether similar siRNA sequences are already reported in database or not. siRNAmod-Map enables the user to map cm-siRNAs reported in the database against a particular nucleotide sequence. In addition, we have also provided submit option for new chemical entries by various researchers. Further help page and other related links are provided.
Implementation
Database front end is developed using Perl, PHP, JavaScript HTML and CSS. It is maintained on MySQL server using LAMP software solution. The resource is hosted on IBM SAS ×3800 machine. siRNAmod server and its integrated tools are compatible with all browsers like Google Chrome, Mozilla Firefox and Safari. However, structure based search tool (MarvinSketch) needs java to be enabled on the browser.
Discussion
siRNAs are being actively tested as new potential therapeutics against various disorders as well as pathogens37,41,42. More than 20 siRNA based therapeutics are currently at different phases of clinical trials43. Examples of such cm-siRNA drugs are SPC2996, EZN3042 and SPC3649 for leukemia, solid tumors and Hepatitis C virus (HCV) infection respectively43. Chemical modifications remain the critical aspect for development of siRNA therapeutics to reduce their inherent limitations6,44. For example, 2′-fluoro, 2′-O-methyl, 2′-deoxy, unlocked nucleic acid, 2′-hydroxy and phosphorothioate modification increase siRNA serum stability19,20. Likewise, certain nucleotide motifs in siRNA sequence as UA and CA upon modification with 2′-O-methyl or 2′-fluoro enhances the nuclease resistance45. Locked nucleic acid, 2′-O-methyl, 2′-fluoro moieties thermodynamically stabilize siRNA ends while unlocked nucleic acid, phosphorothioate, ethylamino, dihydrouracil modifications destabilize them46. Stabilizing the 5′-end and destabilizing the 3′-end of passenger strand enhances the potency of siRNA and vice versa46. siRNAs of length 19 to 21 base pairs are less immunogenic than longer ones47. Modifications like 2′-thiouridine, 4′-thiouridine, 2′-deoxyuridine reduce Protein kinase R (PKR) activation48 while 2′-fluoro, 2′-deoxy and 2′-O-methyl modifications abolish Toll like receptor (TLR) interaction preserving silencing activity even upon heavy modification46.
Various comprehensive studies in this field have been conducted which employed new strategies and methods. Bramsen and coworkers24 utilize 21 different chemical modifications on various positions of sense and antisense strands. Both strands were then combined to generate permutations and combinations of cm-siRNAs. Butora et al. employed walkthrough method13,32, which replaces single or multiple nucleotides in siRNA with different modifications sequentially from one end to another and checking their biological impact. Further effect on enhanced efficacy due to chemical modifications was tested using different siRNAs concentrations49. Further, compatibility of RNAi protein machinery with cm-siRNA format has been investigated50,51. Peel et al. examined target degradation efficacy of small molecules conjugated to siRNAs52. Similarly, other groups also studied terminal modifications like 5′-phosphate53 or serinol modification54.
We plotted overall database chemical modifications statistics. Complementary positions from 1 to 19 of cm-siRNAs show comparatively similar frequency distribution of cumulative modifications as depicted in Fig. 3. Overall statistical trend of chemical modifications depicts that terminal positions 1, 2, 3 and 20, 21 are highly altered comparative to central portion as shown in supplementary Fig. S2. This may be probably due to ease of end modifications and their role in nuclease resistance55. Further, antisense seems a bit less modified than sense strand, may be due to their differential roles in RISC-machinery56. Hence, modifying each strand distinctly may prove a handy tool to improve their cm-siRNA functioning.
Pattern of cumulative occurrence of nucleotides modified on sense and antisense strand of cm-siRNAs is depicted in supplementary Fig. S3. It reveals that majorly one, two or three nucleotides have been engineered for chemical modifications on either siRNA strand. However only in limited studies nucleotides on all twenty-one positions in a siRNA have been modified. Reason for this could be the toxicity associated with increased modifications57. Combinations of different unique modifications i.e. how many exclusively different chemical moieties are integrated in cm-siRNA is displayed in supplementary Fig. S4. It elucidates that maximum 5 different combinations of modifications are seen so far in cm-siRNAs. These conglomerates need to be optimized for construction of biologically significant siRNAs24. So, there is further scope to explore these combinations.
For data retrieval and usage we have designed user-friendly tools and web pages. Users can select the unique chemical modification list on web server and check their patterns i.e. on what positions the modifications are reported along with their biological effects. We have provided the browse option with respect to modifications and positions on either sense or antisense strand of siRNA. User can also analyze cm-siRNA sequences based on the positions of modifications. Furthermore, structure search option is also available to draw the structure and search against the database. The database would be updated half yearly/annually or on availability of the enough data in the literature.
Methods
Data acquisition
Literature was exhaustively searched for experimentally validated chemically modified-siRNAs (cm-siRNAs) for extraction of relevant information using keywords related to (i) RNAi (ii) modifications (iii) stability, utilizing advanced search option in PubMed. The search resulted in about 900 articles as on June 2015, which we examined for relevant experimental data on cm-siRNAs. Studies related to chemical modification for their synthesis, lacking complete biological information and reviews were excluded. About 500 potential articles were scrutinized to mine the required fields. Finally from 96 articles, comprehensive information of 4894 cm-siRNAs was extracted.
We have also integrated chemical information about modifications introduced in the siRNAs. This information is derived form public chemical databases namely PubChem58, ChemSpider (http://www.chemspider.com), ChEMBL (https://www.ebi.ac.uk/chembl). Structures of some chemical modifications were hand drawn using MarvinSketch package (http://www.chemaxon.com), as they were not available in above chemical repositories.
Database organization
siRNAmod provides comprehensive information on experimentally validated cm-siRNA sequences and their chemical modifications. Database is structured to provide information in more than 20 fields broadly divided into; (A) Chemical modification (B) siRNA sequence (C) siRNA activity (D) Experimental details. The database architecture is shown in Fig. 5.

Diagrammatic representation of siRNAmod architecture.
(A)Chemical modification
It includes the information about the chemical moiety and all its related details for both sense and antisense strand distinctly.
(i) Name of chemical modifications for example “2-O-methyl”. If multiple modifications are present then they are separated by asterisk (*) mark e.g. “ 2,4-Bridged nucleic acid* Deoxythymidine”; (ii) their unique and cumulative numerical magnitude (iii) positions of chemical modifications i.e. positions on the siRNA sequence where the modifications are incorporated. When one modification is present at more than one position, it is represented by the position separated by comma e.g. “1,2,3,4,5” and two modifications at respective positions are separated by asterisks as “1,2,3,4,5 * 23,24”; (iv) modification component of nucleotide presents the information about the modified substructure of the nucleotide i.e. whether sugar, base or phosphate part is modified; (v) structures of modification; (vi) SMILES (simplified molecular input line entry system canonical), InCheI (International Chemical Identifier); (vii) systematic name etc.; (viii) natural or non-natural presence of chemical modification.
(B) siRNA sequences
It encompasses information relevant to cm-siRNA sequences and includes (ix) sequence of unmodified siRNA i.e. the basic sequence on which the modification is performed both sense and antisense; (x) length of the siRNA; (xi) Id or name of siRNA used in the respective article. Besides, 435 unmodified siRNAs that were used as controls in the respective studies were also extracted and provided separately in the database.
(C) siRNA activity
It comprises of (xii) biological activity or efficacy of the cm-siRNA as percentage of target mRNA degraded. Besides, inhibition concentration-50 (IC50) values are sometimes provided in the studies instead of percentage inhibition. Qualitative representation of activity is also used to represent the activity of modified siRNA where exact quantitative values were not given, using the terms like high or low activity comparatively.
(D) Experimental details
It covers rest of the relevant information as (xiii) target gene; (xiv) melting temperature of modified siRNA sequence; (xv) siRNA concentration; (xvi) experiment used to check activity; (xvii) cell line or organism; (xviii) transfection method; (xix) post transfection duration; (xx) reference.
Conclusion and Future Developments
Chemical modifications in siRNAs are one of the inevitable steps to ameliorate their therapeutic potential. siRNAmod is the first comprehensive platform of 4894 cm-siRNAs with 128 unique chemical modifications. It provides manually curated information about chemical modification, sequences, siRNA activity and experimental details. Chemical information includes structure, SMILES, InCheI to make the database chemically aware. User-friendly web interface is provided to retrieve the desired curated information with ease. Exploration of chemical modifications and analysis in siRNAs and has been accelerated in last decade and data is continuously growing. In future, we would update the database and include cm-siRNA based design tools and algorithm. We hope that siRNAmod would be helpful for further analysis, interpretation and to accelerate the development of siRNA-based therapeutics.
Additional Information
How to cite this article: Dar, S. A. et al. siRNAmod: A database of experimentally validated chemically modified siRNAs. Sci. Rep. 6, 20031; doi: 10.1038/srep20031 (2016).
References
Fire, A. et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391, 806–811, 10.1038/35888 (1998).
Woessmann, W., Damm-Welk, C., Fuchs, U. & Borkhardt, A. RNA interference: new mechanisms for targeted treatment? Reviews in clinical and experimental hematology 7, 270–291 (2003).
Filipowicz, W. RNAi: the nuts and bolts of the RISC machine. Cell 122, 17–20, 10.1016/j.cell.2005.06.023 (2005).
Davis, M. E. et al. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 464, 1067–1070, 10.1038/nature08956 (2010).
Joo, M. K., Yhee, J. Y., Kim, S. H. & Kim, K. The potential and advances in RNAi therapy: Chemical and structural modifications of siRNA molecules and use of biocompatible nanocarriers. Journal of controlled release : official journal of the Controlled Release Society, 10.1016/j.jconrel.2014.05.030 (2014).
Gallas, A., Alexander, C., Davies, M. C., Puri, S. & Allen, S. Chemistry and formulations for siRNA therapeutics. Chemical Society reviews 42, 7983–7997, 10.1039/c3cs35520a (2013).
Gaglione, M. & Messere, A. Recent progress in chemically modified siRNAs. Mini reviews in medicinal chemistry 10, 578–595 (2010).
Stephenson, M. L. & Zamecnik, P. C. Inhibition of Rous sarcoma viral RNA translation by a specific oligodeoxyribonucleotide. Proceedings of the National Academy of Sciences of the United States of America 75, 285–288 (1978).
Wilson, C. & Keefe, A. D. Building oligonucleotide therapeutics using non-natural chemistries. Current opinion in chemical biology 10, 607–614, 10.1016/j.cbpa.2006.10.001 (2006).
Elmen, J. et al. Locked nucleic acid (LNA) mediated improvements in siRNA stability and functionality. Nucleic acids research 33, 439–447, 10.1093/nar/gki193 (2005).
Kenski, D. M. et al. Analysis of acyclic nucleoside modifications in siRNAs finds sensitivity at position 1 that is restored by 5′-terminal phosphorylation both in vitro and in vivo. Nucleic acids research 38, 660–671, 10.1093/nar/gkp913 (2010).
Takahashi, M. et al. Intracellular stability of 2′-OMe-4′-thioribonucleoside modified siRNA leads to long-term RNAi effect. Nucleic acids research 40, 5787–5793, 10.1093/nar/gks204 (2012).
Prakash, T. P. et al. Positional effect of chemical modifications on short interference RNA activity in mammalian cells. Journal of medicinal chemistry 48, 4247–4253, 10.1021/jm050044o (2005).
Soutschek, J. et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature 432, 173–178, 10.1038/nature03121 (2004).
Petersen, M. & Wengel, J. LNA: a versatile tool for therapeutics and genomics. Trends in biotechnology 21, 74–81, 10.1016/S0167-7799(02)00038-0 (2003).
Grunweller, A. et al. Comparison of different antisense strategies in mammalian cells using locked nucleic acids, 2′-O-methyl RNA, phosphorothioates and small interfering RNA. Nucleic acids research 31, 3185–3193 (2003).
Glud, S. Z. et al. Naked siLNA-mediated gene silencing of lung bronchoepithelium EGFP expression after intravenous administration. Oligonucleotides 19, 163–168, 10.1089/oli.2008.0175 (2009).
Langkjaer, N., Pasternak, A. & Wengel, J. UNA (unlocked nucleic acid): a flexible RNA mimic that allows engineering of nucleic acid duplex stability. Bioorganic & medicinal chemistry 17, 5420–5425, 10.1016/j.bmc.2009.06.045 (2009).
Aboul-Fadl, T. Antisense oligonucleotides: the state of the art. Current medicinal chemistry 12, 2193–2214 (2005).
Detzer, A. & Sczakiel, G. Phosphorothioate-stimulated uptake of siRNA by mammalian cells: a novel route for delivery. Current topics in medicinal chemistry 9, 1109–1116 (2009).
Chiu, Y. L. & Rana, T. M. siRNA function in RNAi: a chemical modification analysis. Rna 9, 1034–1048 (2003).
Layzer, J. M. et al. In vivo activity of nuclease-resistant siRNAs. Rna 10, 766–771 (2004).
Odadzic, D. et al. Synthesis of 2′-O-modified adenosine building blocks and application for RNA interference. Bioorganic & medicinal chemistry 16, 518–529, 10.1016/j.bmc.2007.09.019 (2008).
Bramsen, J. B. et al. A large-scale chemical modification screen identifies design rules to generate siRNAs with high activity, high stability and low toxicity. Nucleic acids research 37, 2867–2881, 10.1093/nar/gkp106 (2009).
Somoza, A., Silverman, A. P., Miller, R. M., Chelliserrykattil, J. & Kool, E. T. Steric effects in RNA interference: probing the influence of nucleobase size and shape. Chemistry 14, 7978–7987, 10.1002/chem.200800837 (2008).
Sipa, K. et al. Effect of base modifications on structure, thermodynamic stability and gene silencing activity of short interfering RNA. Rna 13, 1301–1316, 10.1261/rna.538907 (2007).
Peacock, H., Kannan, A., Beal, P. A. & Burrows, C. J. Chemical modification of siRNA bases to probe and enhance RNA interference. The Journal of organic chemistry 76, 7295–7300, 10.1021/jo2012225 (2011).
Hall, A. H., Wan, J., Shaughnessy, E. E., Ramsay Shaw, B. & Alexander, K. A. RNA interference using boranophosphate siRNAs: structure-activity relationships. Nucleic acids research 32, 5991–6000, 10.1093/nar/gkh936 (2004).
Potenza, N. et al. RNA interference in mammalia cells by RNA-3′-PNA chimeras. International journal of molecular sciences 9, 299–315 (2008).
Detzer, A., Overhoff, M., Mescalchin, A., Rompf, M. & Sczakiel, G. Phosphorothioate-stimulated cellular uptake of siRNA: a cell culture model for mechanistic studies. Current pharmaceutical design 14, 3666–3673 (2008).
Braasch, D. A. et al. Biodistribution of phosphodiester and phosphorothioate siRNA. Bioorganic & medicinal chemistry letters 14, 1139–1143, 10.1016/j.bmcl.2003.12.074 (2004).
Butora, G. et al. Nucleoside optimization for RNAi: a high-throughput platform. Journal of the American Chemical Society 133, 16766–16769, 10.1021/ja2068774 (2011).
Broering, R. et al. Chemical modifications on siRNAs avoid Toll-like-receptor-mediated activation of the hepatic immune system in vivo and in vitro. International immunology 26, 35–46, 10.1093/intimm/dxt040 (2014).
Ren, Y. et al. siRecords: a database of mammalian RNAi experiments and efficacies. Nucleic acids research 37, D146–149, 10.1093/nar/gkn817 (2009).
Chalk, A. M., Warfinge, R. E., Georgii-Hemming, P. & Sonnhammer, E. L. siRNAdb: a database of siRNA sequences. Nucleic acids research 33, D131–134, 10.1093/nar/gki136 (2005).
Truss, M. et al. HuSiDa–the human siRNA database: an open-access database for published functional siRNA sequences and technical details of efficient transfer into recipient cells. Nucleic acids research 33, D108–111, 10.1093/nar/gki131 (2005).
Thakur, N., Qureshi, A. & Kumar, M. VIRsiRNAdb: a curated database of experimentally validated viral siRNA/shRNA. Nucleic acids research 40, D230–236, 10.1093/nar/gkr1147 (2012).
Tyagi, A. et al. HIVsirDB: a database of HIV inhibiting siRNAs. PloS one 6, e25917, 10.1371/journal.pone.0025917 (2011).
Cantara, W. A. et al. The RNA Modification Database, RNAMDB: 2011 update. Nucleic acids research 39, D195-201, 10.1093/nar/gkq1028 (2011).
Herraez, A. Biomolecules in the computer: Jmol to the rescue. Biochemistry and molecular biology education : a bimonthly publication of the International Union of Biochemistry and Molecular Biology 34, 255–261, 10.1002/bmb.2006.494034042644 (2006).
Ozcan, G., Ozpolat, B., Coleman, R. L., Sood, A. K. & Lopez-Berestein, G. Preclinical and clinical development of siRNA-based therapeutics. Advanced drug delivery reviews, 10.1016/j.addr.2015.01.007 (2015).
Burnett, J. C., Rossi, J. J. & Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnology journal 6, 1130–1146, 10.1002/biot.201100054 (2011).
Lares, M. R., Rossi, J. J. & Ouellet, D. L. RNAi and small interfering RNAs in human disease therapeutic applications. Trends in biotechnology 28, 570–579, 10.1016/j.tibtech.2010.07.009 (2010).
Engels, J. W. Gene silencing by chemically modified siRNAs. New biotechnology 30, 302–307, 10.1016/j.nbt.2012.07.002 (2013).
Turner, J. J., Jones, S. W., Moschos, S. A., Lindsay, M. A. & Gait, M. J. MALDI-TOF mass spectral analysis of siRNA degradation in serum confirms an RNAse A-like activity. Molecular bioSystems 3, 43–50, 10.1039/b611612d (2007).
Bramsen, J. B. & Kjems, J. Development of Therapeutic-Grade Small Interfering RNAs by Chemical Engineering. Frontiers in genetics 3, 154, 10.3389/fgene.2012.00154 (2012).
Foster, D. J. et al. Comprehensive evaluation of canonical versus Dicer-substrate siRNA in vitro and in vivo. Rna 18, 557–568, 10.1261/rna.031120.111 (2012).
Nallagatla, S. R. & Bevilacqua, P. C. Nucleoside modifications modulate activation of the protein kinase PKR in an RNA structure-specific manner. Rna 14, 1201–1213, 10.1261/rna.1007408 (2008).
Haringsma, H. J. et al. mRNA knockdown by single strand RNA is improved by chemical modifications. Nucleic acids research 40, 4125–4136, 10.1093/nar/gkr1301 (2012).
Collingwood, M. A. et al. Chemical modification patterns compatible with high potency dicer-substrate small interfering RNAs. Oligonucleotides 18, 187–200, 10.1089/oli.2008.0123 (2008).
Somoza, A., Terrazas, M. & Eritja, R. Modified siRNAs for the study of the PAZ domain. Chemical communications 46, 4270–4272, 10.1039/c003221b (2010).
Peel, B. J., Hagen, G., Krishnamurthy, K. & Desaulniers, J. P. Conjugation and Evaluation of Small Hydrophobic Molecules to Triazole-Linked siRNAs. ACS medicinal chemistry letters 6, 117–122, 10.1021/ml500260j (2015).
Kenski, D. M., Willingham, A. T., Haringsma, H. J., Li, J. J. & Flanagan, W. M. In vivo activity and duration of short interfering RNAs containing a synthetic 5′-phosphate. Nucleic acid therapeutics 22, 90–95, 10.1089/nat.2011.0333 (2012).
Kamiya, Y. et al. Enhancement of stability and activity of siRNA by terminal substitution with serinol nucleic acid (SNA). Chembiochem : a European journal of chemical biology 15, 2549–2555, 10.1002/cbic.201402369 (2014).
Huang, Y. et al. Exploring directional invasion of serum nuclease into siRNA duplexes by asymmetrical terminal modifications. ChemMedChem 9, 2111–2119, 10.1002/cmdc.201402115 (2014).
Hamada, M. et al. Effects on RNA interference in gene expression (RNAi) in cultured mammalian cells of mismatches and the introduction of chemical modifications at the 3′-ends of siRNAs. Antisense & nucleic acid drug development 12, 301–309, 10.1089/108729002761381285 (2002).
Amarzguioui, M., Holen, T., Babaie, E. & Prydz, H. Tolerance for mutations and chemical modifications in a siRNA. Nucleic acids research 31, 589–595 (2003).
Wang, Y. et al. PubChem BioAssay: 2014 update. Nucleic acids research 42, D1075–1082, 10.1093/nar/gkt978 (2014).
Acknowledgements
Financial support for this work is provided by CSIR-Institute of Microbial Technology (OLP 0083), Council of Scientific and Industrial Research (BSC0121) and Department of Biotechnology, Government of India (GAP0001). Funding for open access charge: CSIR-Institute of Microbial Technology, Sector 39-A, Chandigarh, India.
Author information
Authors and Affiliations
Contributions
M.K. conceived the idea, planned the design, visualization and tools pipeline and coordinated the entire project. S.D. and A.T. manually collected and curated all the data. S.D. and A.Q. designed the web server part of the database. S.D. and M.K. performed data analysis, interpreted results and drafted the manuscript. All authors read and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Dar, S., Thakur, A., Qureshi, A. et al. siRNAmod: A database of experimentally validated chemically modified siRNAs. Sci Rep 6, 20031 (2016). https://doi.org/10.1038/srep20031
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep20031
This article is cited by
-
Small interfering RNA (siRNA)-based therapeutic applications against viruses: principles, potential, and challenges
Journal of Biomedical Science (2023)
-
Innate immune regulations and various siRNA modalities
Drug Delivery and Translational Research (2023)
-
Investigating the pathogenic SNPs in BLM helicase and their biological consequences by computational approach
Scientific Reports (2020)
-
Delivery of RNAi-Based Therapeutics for Bone Regeneration
Current Osteoporosis Reports (2020)
-
Plant small RNAs: the essential epigenetic regulators of gene expression for salt-stress responses and tolerance
Plant Cell Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
