
Abstract
Seasonal asymmetry in the interannual variations in the daytime and nighttime climate in the Northern Hemisphere (NH) is well documented, but its consequences for vegetation activity remain poorly understood. Here, we investigate the interannual responses of vegetation activity to variations of seasonal mean daytime and nighttime climate in NH (>30 °N) during the past decades using remote sensing retrievals, FLUXNET and tree ring data. Despite a generally significant and positive response of vegetation activity to seasonal mean maximum temperature ( ) in ~22–25% of the boreal (>50 °N) NH between spring and autumn, spring-summer progressive water limitations appear to decouple vegetation activity from the mean summer
) in ~22–25% of the boreal (>50 °N) NH between spring and autumn, spring-summer progressive water limitations appear to decouple vegetation activity from the mean summer  , particularly in climate zones with dry summers. Drought alleviation during autumn results in vegetation recovery from the marked warming-induced drought limitations observed in spring and summer across 24–26% of the temperate NH. Vegetation activity exhibits a pervasively negative correlation with the autumn mean minimum temperature, which is in contrast to the ambiguous patterns observed in spring and summer. Our findings provide new insights into how seasonal asymmetry in the interannual variations in the mean daytime and nighttime climate interacts with water limitations to produce spatiotemporally variable responses of vegetation growth.
, particularly in climate zones with dry summers. Drought alleviation during autumn results in vegetation recovery from the marked warming-induced drought limitations observed in spring and summer across 24–26% of the temperate NH. Vegetation activity exhibits a pervasively negative correlation with the autumn mean minimum temperature, which is in contrast to the ambiguous patterns observed in spring and summer. Our findings provide new insights into how seasonal asymmetry in the interannual variations in the mean daytime and nighttime climate interacts with water limitations to produce spatiotemporally variable responses of vegetation growth.
Similar content being viewed by others


Introduction
The vegetation activity in temperate and cold regions is strongly controlled by climate, which is causing rapid changes in the vegetation activity in the Northern Hemisphere (NH) as the Earth warms1,2,3,4. Satellite observations, Earth system models and atmospheric inversions suggest that growing-season warming has increased the photosynthetic activity of the NH terrestrial vegetation during past decades1,2,4 to a greater extent than other environmental processes (e.g., CO2 and nitrogen fertilization)5. However, the responses of vegetation to climate warming are not uniform in space or time; covariation between the growing season temperature and vegetation activity varies regionally within the NH and appears to be weakening in many regions6. Simultaneously, mean daytime and nighttime warming during the growing-season produce different responses in vegetation productivity in the NH3. As spatial-temporal patterns of daytime and nighttime warming interact with extant seasonal hydrology, energy and phenology cycles, stark differences in the vegetation responses are generated. However, such observed vegetation growth effects due to seasonal asymmetry in the interannual variations in the mean daytime and nighttime climate (VDNC)7,8,9 and the underlying mechanisms that cause these effects to vary spatially remain poorly understood. Therefore, the effects of seasonal asymmetry in interannual variations in the mean diurnal climate on vegetation activity in the NH need to be better understood, especially because this asymmetry is predicted to increase10.
Asymmetry in the seasonal VDNC11,12, together with other factors (e.g., land use changes13), could dramatically modify the seasonal thermo-hydrological patterns that govern land surface-climate feedbacks14,15. At middle and high latitudes in the NH, this asymmetry could hypothetically even modify the sign of biosphere-atmosphere feedbacks, which are known to be globally significant15,16,17. Therefore, in this study, we quantified the general patterns in the interannual responses of active vegetation growth (AVG, i.e., vegetation growth during the growing season) (for more detail, see Methods) at middle and high northern latitudes (>30°N) using 1) the satellite-derived Normalized Difference Vegetation Index (NDVI)18,19 and the Fraction of Absorbed Photosynthetically Active Radiation (FAPAR)20, 2) FLUXNET-based up-scaled gross primary productivity (GPP)21 and 3) tree growth increments to the interannual variability (IAV) of seasonal mean maximum temperature ( ), mean minimum temperature (
), mean minimum temperature ( ) and water availability index (WAI) during past decades (for more detail, see Methods).
) and water availability index (WAI) during past decades (for more detail, see Methods).
Results
Covariance between vegetation activity and the seasonal mean 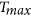
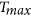
Partial correlation analyses reveal a clear spatial pattern in the interannual responses of the mean growing-season (April-October) NDVI ( ) to the IAV of the seasonal mean
) to the IAV of the seasonal mean  during 1982–2008 (Fig. 1).
during 1982–2008 (Fig. 1).  generally exhibits a positive correlation with
generally exhibits a positive correlation with  in spring (April-May) (
in spring (April-May) ( ), summer (June-August) (
), summer (June-August) ( ) and autumn (September-October)
) and autumn (September-October)  in most (~60–70%) of the boreal NH (Figs 1 and 2a, Supplementary Table S1, Figs S1–S3), with statistically significant (p < 0.10) partial correlation coefficients (R > 0.30) in ~22–25% of boreal NH despite the great spatial variations in the responses (Fig. 1, Supplementary Table S2 and Figs S1, S3). Such response patterns of the AVG to the IAV of the seasonal mean
in most (~60–70%) of the boreal NH (Figs 1 and 2a, Supplementary Table S1, Figs S1–S3), with statistically significant (p < 0.10) partial correlation coefficients (R > 0.30) in ~22–25% of boreal NH despite the great spatial variations in the responses (Fig. 1, Supplementary Table S2 and Figs S1, S3). Such response patterns of the AVG to the IAV of the seasonal mean  in the boreal NH are also supported by the analyses of mean growing-season FAPAR (
in the boreal NH are also supported by the analyses of mean growing-season FAPAR ( ) (Supplementary Figs S4a–c and S5a) and the results of independent ridge regression analyses (for more detail, see the supplementary text) of the relationships among
) (Supplementary Figs S4a–c and S5a) and the results of independent ridge regression analyses (for more detail, see the supplementary text) of the relationships among 
 and the seasonal mean
and the seasonal mean  during 1982–2008 (Supplementary Figs S6a–c and S7a–c). Coincidently, the ridge regression analyses reveal a generally positive interannual sensitivity of total growing-season GPP (
during 1982–2008 (Supplementary Figs S6a–c and S7a–c). Coincidently, the ridge regression analyses reveal a generally positive interannual sensitivity of total growing-season GPP ( ) to seasonal mean
) to seasonal mean  (
( , indicated by the coefficient of ridge regression) from spring to autumn in most (~51–89%) of the boreal NH (with statistical significance in ~20–62% of the boreal NH) despite large spatial variations (Fig. 3a, Supplementary Fig. S8a–c), with
, indicated by the coefficient of ridge regression) from spring to autumn in most (~51–89%) of the boreal NH (with statistical significance in ~20–62% of the boreal NH) despite large spatial variations (Fig. 3a, Supplementary Fig. S8a–c), with  ranging from 36.5 ± 62.4 to 51.3 ± 88.7 g C m−2 yr−1 °C−1 between seasons (Fig. 3a, Supplementary Fig. S8a–c).
ranging from 36.5 ± 62.4 to 51.3 ± 88.7 g C m−2 yr−1 °C−1 between seasons (Fig. 3a, Supplementary Fig. S8a–c).

Spatial patterns of the interannual responses of the mean growing-season (April-October) NDVI and TRI to seasonal VDNC in the mid- and high-latitude NH.
Spearman partial correlation coefficients between the mean growing-season NDVI ( ) (during 1982–2008) and TRI (dots) (during 1950–2008, if available) as well as between the seasonal mean maximum temperature (
) (during 1982–2008) and TRI (dots) (during 1950–2008, if available) as well as between the seasonal mean maximum temperature ( ), mean minimum temperature (
), mean minimum temperature ( ) and water availability index (WAI) from spring to autumn are shown. R = ±0.39, R = ±0.30 and R = ±0.20 in the colorbar correspond to the 5%, 10% and 20% significance levels for the Spearman partial correlation between interannual variations of
) and water availability index (WAI) from spring to autumn are shown. R = ±0.39, R = ±0.30 and R = ±0.20 in the colorbar correspond to the 5%, 10% and 20% significance levels for the Spearman partial correlation between interannual variations of  and seasonal mean climate, respectively. This figure is created by MATLAB (R2012b).
and seasonal mean climate, respectively. This figure is created by MATLAB (R2012b).

PDFs of the partial correlation coefficients between the mean growing-season (April-October) NDVI and the seasonal VDNC.
PDFs of the partial correlation coefficients (PR) between the mean growing-season NDVI and spring (green lines), summer (blue lines) and autumn (brown lines) maximum temperature (a), minimum temperature (b) and water availability index (c) in temperate (<50 °N, dotted lines) and boreal (≥50 °N, solid lines) regions of the NH. The percentages of the pixels that exhibit significant (p < 0.10) positive (green bars) and negative (red bars) correlations between the mean growing-season NDVI and spring (SP), summer (SU) and autumn (AU) maximum temperature, minimum temperature and WAI in temperate (light color) and boreal (dark color) regions are specified in the insets. The Spearman correlation coefficients R ± 0.51 and ±0.30 in the x axis tick label correspond to the 1% and 10% significance levels of Student’s t-test, respectively.

Interannual sensitivity of the growing-season (April-October) GPP to the seasonal VDNC in different climate zones.
The sensitivity of the growing-season GPP to spring (light color), summer (medium color) and autumn (dark color) maximum temperature, minimum temperature and WAI in different climate zones are shown in panels a-c, respectively. The circles and short red lines in each box indicate the mean and median values, respectively, of the sensitivity of the growing-season GPP to the seasonal VDNC. The dashed lines and filled areas in each box represent the 5th to 95th and 25th to75th percentiles, respectively, of the sensitivity of the growing-season GPP to the seasonal VDNC. Seven major climate zones based on the Köppen–Geiger climate classification were considered in this study: RegAR (arid region, grouping of BWk, BWh, BSk, BSh), RegTH (temperate humid region, grouping of Cfa, Cfb and Cfc), RegTA (temperate dry region, grouping of Csa, Csb, Csc, Cwa, Cwb and Cwc), RegAH (cold humid region, grouping of Dfa, Dfb, Dfc and Dfd), RegAS (cold summer dry region, grouping of Dsa, Dsb, Dsc, and, Dsd), RegAW (cold winter dry region, grouping of Dwa, Dwb, Dwc, and, Dwd) and RegET (polar tundra, ET).
By contrast,  exhibits a significant (p < 0.10) negative correlation (R < −0.30) with the IAV of mean
exhibits a significant (p < 0.10) negative correlation (R < −0.30) with the IAV of mean  and
and  in ~13–16% of the temperate NH (<50 °N) during 1982–2008 (Fig. 1, Supplementary Table S1 and Fig. S1). The most prominent negative correlations are observed in the arid climate zones of the northern U.S.A., southern Eurasia and parts of the Mediterranean regions (Fig. 1, Supplementary Table S2 and Fig. S1), which are generally dominated by temperate grasslands, dry shrublands/forests or forest-grassland ecotones. Analyses of the relationships between the standard tree ring index (TRI) (during 1950–2008, if available),
in ~13–16% of the temperate NH (<50 °N) during 1982–2008 (Fig. 1, Supplementary Table S1 and Fig. S1). The most prominent negative correlations are observed in the arid climate zones of the northern U.S.A., southern Eurasia and parts of the Mediterranean regions (Fig. 1, Supplementary Table S2 and Fig. S1), which are generally dominated by temperate grasslands, dry shrublands/forests or forest-grassland ecotones. Analyses of the relationships between the standard tree ring index (TRI) (during 1950–2008, if available),  ,
,  and the seasonal mean
and the seasonal mean  confirm this pattern (Figs 1 and 3a, Supplementary Table S4, Figs S1, S4a–c, S5a, S7 and S8a–c). Partial correlation analyses reveal a generally negative correlation of TRI with mean
confirm this pattern (Figs 1 and 3a, Supplementary Table S4, Figs S1, S4a–c, S5a, S7 and S8a–c). Partial correlation analyses reveal a generally negative correlation of TRI with mean  and
and  in the temperate NH during 1950–2008 (if available) (with statistical significance for ~15% of samples) compared with a generally positive correlation in the boreal NH (with statistical significance for ~13% of samples) (Fig. 1). Similarly, ridge regression reveals that
in the temperate NH during 1950–2008 (if available) (with statistical significance for ~15% of samples) compared with a generally positive correlation in the boreal NH (with statistical significance for ~13% of samples) (Fig. 1). Similarly, ridge regression reveals that  in arid regions of the temperate NH (i.e., RegAR) exhibits a pervasively (~83–85% of RegAR) negative sensitivity to the IAV in
in arid regions of the temperate NH (i.e., RegAR) exhibits a pervasively (~83–85% of RegAR) negative sensitivity to the IAV in  and
and  during 1982–2008, with mean
during 1982–2008, with mean  values of –24.9 ± 29.7 and –24.33 ± 28.5 g C m−2 yr−1 °C−1, respectively (Fig. 3a, Supplementary Table S3).
values of –24.9 ± 29.7 and –24.33 ± 28.5 g C m−2 yr−1 °C−1, respectively (Fig. 3a, Supplementary Table S3).
Both partial correlation and ridge regression analyses of the relationships among  ,
,  ,
,  , TRI and the seasonal mean
, TRI and the seasonal mean  consistently reveal a divergent response of the AVG to the IAV in the seasonal mean
consistently reveal a divergent response of the AVG to the IAV in the seasonal mean  for portions of the middle and high latitudes of the NH in two aspects. First, the AVG in parts of the boreal NH, especially regions with dry summer (i.e., RegAS) and warm temperate regions (e.g., the western and southeastern U.S.A.), responds differently to mean
for portions of the middle and high latitudes of the NH in two aspects. First, the AVG in parts of the boreal NH, especially regions with dry summer (i.e., RegAS) and warm temperate regions (e.g., the western and southeastern U.S.A.), responds differently to mean  and
and  (Fig. 1, Supplementary Table S2, Table S3, Figs S1, S3, S4a,b and S6-S7a,b): The pervasive (~31%) significant positive correlation (R > 0.30) between
(Fig. 1, Supplementary Table S2, Table S3, Figs S1, S3, S4a,b and S6-S7a,b): The pervasive (~31%) significant positive correlation (R > 0.30) between  and
and  in RegAS tends to weaken (~9.1%) in summer (Fig. 1, Supplementary Table S2, Figs S1 and S3), whereas the
in RegAS tends to weaken (~9.1%) in summer (Fig. 1, Supplementary Table S2, Figs S1 and S3), whereas the  maintains a significant positive correlation (R > 0.30) with the WAI (for ~15–17% of the samples) in both spring and summer (Fig. 1, Supplementary Table S2, Figs S1 and S2c). Eddy covariance observations in these regions support this notion of seasonal water limitation, indicating that
maintains a significant positive correlation (R > 0.30) with the WAI (for ~15–17% of the samples) in both spring and summer (Fig. 1, Supplementary Table S2, Figs S1 and S2c). Eddy covariance observations in these regions support this notion of seasonal water limitation, indicating that  responds positively to WAI in spring and summer during 1982–2008 (with spring and summer
responds positively to WAI in spring and summer during 1982–2008 (with spring and summer  values of 1.2 ± 1.7 and 0.9 ± 1.8 g C m−2 yr−1 mm−1, respectively, in RegAS) (Fig. 3c, Supplementary Table S3, Figs S8g,h,S9g,h and S10). In addition to the regions with dry summers in the boreal NH,
values of 1.2 ± 1.7 and 0.9 ± 1.8 g C m−2 yr−1 mm−1, respectively, in RegAS) (Fig. 3c, Supplementary Table S3, Figs S8g,h,S9g,h and S10). In addition to the regions with dry summers in the boreal NH,  in certain parts of the temperate NH also exhibits divergent interannual sensitivity to
in certain parts of the temperate NH also exhibits divergent interannual sensitivity to  and
and  (Fig. 3a, S9a,b and S10a), with
(Fig. 3a, S9a,b and S10a), with  values of 8.9 ± 22.1 in spring and −20.9 ± 43.6 g C m−2 yr−1 °C−1 in summer in RegTH (Fig. 3a, Supplementary Fig. S8a,b). At the same time, the TRI values in the temperate NH exhibit pervasive positive correlations with
values of 8.9 ± 22.1 in spring and −20.9 ± 43.6 g C m−2 yr−1 °C−1 in summer in RegTH (Fig. 3a, Supplementary Fig. S8a,b). At the same time, the TRI values in the temperate NH exhibit pervasive positive correlations with  and
and  (with statistical significance at p < 0.05 level in ~14% and ~40% of samples, respectively) during 1950–2008 (Figs 1 and S1). Second, the interannual responses of the AVG to
(with statistical significance at p < 0.05 level in ~14% and ~40% of samples, respectively) during 1950–2008 (Figs 1 and S1). Second, the interannual responses of the AVG to  diverge from the responses to
diverge from the responses to  and
and  in the majority of the temperate NH, especially in regions with drought limitations (e.g., RegAR and RegTA) (Supplementary Table S2). The
in the majority of the temperate NH, especially in regions with drought limitations (e.g., RegAR and RegTA) (Supplementary Table S2). The  exhibits a significant positive correlation (R > 0.30) with mean
exhibits a significant positive correlation (R > 0.30) with mean  in ~15–16% of the temperate NH but exhibits a significant positive correlation with
in ~15–16% of the temperate NH but exhibits a significant positive correlation with  and
and  in only ~<8% of that area (Figs 1 and 2a, Table S2, Fig. S1 and S2a).
in only ~<8% of that area (Figs 1 and 2a, Table S2, Fig. S1 and S2a).
Covariance between vegetation activity and the seasonal mean 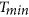
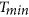
The interannual responses of the AVG in the middle and high latitudes of the NH to the mean spring and summer minimum temperatures ( and
and  , respectively) are relatively ambiguous (Fig. 1, Supplementary Fig. S1, S2b, S4d,e, S5b and S6-S9d,e), yet the trends in
, respectively) are relatively ambiguous (Fig. 1, Supplementary Fig. S1, S2b, S4d,e, S5b and S6-S9d,e), yet the trends in  ,
,  and TRI indicate that the AVG responds positively to
and TRI indicate that the AVG responds positively to  in certain parts of the temperate NH, particularly in arid regions (primarily in the central U.S.A. and southern Eurasia). The correlation with
in certain parts of the temperate NH, particularly in arid regions (primarily in the central U.S.A. and southern Eurasia). The correlation with  is statistically significant (R > 0.30, p < 0.10) in ~17.5% of these regions (Fig. 1, Supplementary Table S2, Fig. S1). Interestingly, such positive responses of the AVG to mean
is statistically significant (R > 0.30, p < 0.10) in ~17.5% of these regions (Fig. 1, Supplementary Table S2, Fig. S1). Interestingly, such positive responses of the AVG to mean  tend to weaken from spring to summer in arid and temperate arid regions (Figs 1 and 3d,e, Supplementary Fig. S1) and instead become predominant in the high-latitude NH. Significant positive responses of
tend to weaken from spring to summer in arid and temperate arid regions (Figs 1 and 3d,e, Supplementary Fig. S1) and instead become predominant in the high-latitude NH. Significant positive responses of  and
and  to
to  are observed in ~12–20% and ~23–64%, respectively, of cold regions and tundra (RegAH, RegAS and RegET) (Figs 1 and 3b, Supplementary Fig. S1, S3 and Table S2). Coincidently, the TRI exhibits a more pervasive positive correlation with
are observed in ~12–20% and ~23–64%, respectively, of cold regions and tundra (RegAH, RegAS and RegET) (Figs 1 and 3b, Supplementary Fig. S1, S3 and Table S2). Coincidently, the TRI exhibits a more pervasive positive correlation with  (~15.8%) than with
(~15.8%) than with  (~7.9%) in the temperate NH (Fig. 1).
(~7.9%) in the temperate NH (Fig. 1).
Contrary to the ambiguous relationships between AVG and  and
and  , both
, both  and
and  exhibit pervasively (60–63%) negative responses to
exhibit pervasively (60–63%) negative responses to  in most of the temperate and cold NH except in certain mountainous regions (e.g., the Rocky Mountains in western North America). Statistical significance is observed for
in most of the temperate and cold NH except in certain mountainous regions (e.g., the Rocky Mountains in western North America). Statistical significance is observed for  in ~16% and ~18% of the temperate and boreal NH, respectively (Figs 1,2b and 3b Supplementary Table S1, Table S3, Fig. S1 and Fig. S3). However, TRI data indicate a persistently positive rather than negative correlation with
in ~16% and ~18% of the temperate and boreal NH, respectively (Figs 1,2b and 3b Supplementary Table S1, Table S3, Fig. S1 and Fig. S3). However, TRI data indicate a persistently positive rather than negative correlation with  during 1950–2008 (Fig. 1, Supplementary Fig. S1). Additionally, the observed divergent response patterns between AVG and seasonal VDNC are not susceptible to the arbitrary definitions of active growing seasons (Supplementary Fig. S11-S12) and selection of analysis periods (see Methods for details) (Supplementary Fig. S13).
during 1950–2008 (Fig. 1, Supplementary Fig. S1). Additionally, the observed divergent response patterns between AVG and seasonal VDNC are not susceptible to the arbitrary definitions of active growing seasons (Supplementary Fig. S11-S12) and selection of analysis periods (see Methods for details) (Supplementary Fig. S13).
Discussion
The results of both the partial correlation and ridge regression analyses demonstrate that the AVG in the boreal NH is generally temperature-limited: a warmer mean daytime temperature (i.e.,  ) during the active growing season can intuitively enhance the vegetation activity1,2,4,22,23. However, we have identified an obvious seasonally divergent response of the AVG to the IAV in seasonal mean
) during the active growing season can intuitively enhance the vegetation activity1,2,4,22,23. However, we have identified an obvious seasonally divergent response of the AVG to the IAV in seasonal mean  in parts of the high-latitude NH, especially in regions with dry summers. The AVG responds differently to
in parts of the high-latitude NH, especially in regions with dry summers. The AVG responds differently to  and
and  (Fig. 1, Supplementary Table S2, Figs S1, S3, S4a,b and S5-S7a,b) and a pervasive (~31%) significant positive correlation is observed between
(Fig. 1, Supplementary Table S2, Figs S1, S3, S4a,b and S5-S7a,b) and a pervasive (~31%) significant positive correlation is observed between  and
and  shading in summer (Fig. 1, Supplementary Table S2 and Fig. S3). In fact, the AVG in cold regions with dry summers is vulnerable to water stress under daytime warming, thereby resulting in seasonal drought16,24,25, as confirmed by the close (~65.8–66%) positive coupling between the IAV in
shading in summer (Fig. 1, Supplementary Table S2 and Fig. S3). In fact, the AVG in cold regions with dry summers is vulnerable to water stress under daytime warming, thereby resulting in seasonal drought16,24,25, as confirmed by the close (~65.8–66%) positive coupling between the IAV in  and the WAI during spring and summer. In addition to regions with dry summers in the boreal NH, the AVG in certain parts of the temperate NH also responds differently to mean
and the WAI during spring and summer. In addition to regions with dry summers in the boreal NH, the AVG in certain parts of the temperate NH also responds differently to mean  and
and  (Fig. 1 Supplementary Figs S3, S6-S9a,b and S10a). The observed divergent responses of the AVG to
(Fig. 1 Supplementary Figs S3, S6-S9a,b and S10a). The observed divergent responses of the AVG to  and
and  in these regions may be partially attributed to the daytime warming-induced progressive drought from spring to summer, as revealed by a water balance equation and the relationships between the seasonal soil moisture and the VDNC during 1982–2008 (Fig. 4, Supplementary Figs S14-S16). TRI data reveal that tree growth in the temperate NH experiences continuous water limitations during spring and summer, as revealed by pervasive positive correlations of the TRI with
in these regions may be partially attributed to the daytime warming-induced progressive drought from spring to summer, as revealed by a water balance equation and the relationships between the seasonal soil moisture and the VDNC during 1982–2008 (Fig. 4, Supplementary Figs S14-S16). TRI data reveal that tree growth in the temperate NH experiences continuous water limitations during spring and summer, as revealed by pervasive positive correlations of the TRI with  and
and  . In fact, warmer daytime temperatures in spring can lead to an earlier spring phenology and greater photosynthesis26,27, thereby enhancing vegetation growth as long as the soil water and atmospheric vapor pressure are sufficient. Conversely, if the increased warming rates during the early growing season are unmatched by water resources, the warmer temperatures can trigger a prolonged warming-induced drought, especially in regions that suffer seasonal drought limitations (e.g., RegAS and most regions in the temperate NH) (Fig. 4). Such cases of excessive soil water stress or, more commonly, vapor pressure deficits (Supplementary Figs S14-S16, ref. Liu et al., 201328) can dramatically reduce vegetation growth and even negate springtime ecosystem carbon gains16,25,29.
. In fact, warmer daytime temperatures in spring can lead to an earlier spring phenology and greater photosynthesis26,27, thereby enhancing vegetation growth as long as the soil water and atmospheric vapor pressure are sufficient. Conversely, if the increased warming rates during the early growing season are unmatched by water resources, the warmer temperatures can trigger a prolonged warming-induced drought, especially in regions that suffer seasonal drought limitations (e.g., RegAS and most regions in the temperate NH) (Fig. 4). Such cases of excessive soil water stress or, more commonly, vapor pressure deficits (Supplementary Figs S14-S16, ref. Liu et al., 201328) can dramatically reduce vegetation growth and even negate springtime ecosystem carbon gains16,25,29.

Conceptual diagram for water-mediated interannual responses of vegetation activity to the seasonal VDNC in the mid- and high-latitudinal NH.
The green and red lines indicate the effects of seasonal variations in diurnal warming on plant photosynthesis/GPP and autotrophic respiration ( ), respectively. The relative importance of the impacts and the relative warming rates between seasons are indicated by the line width (broader lines indicate greater importance). The blue arrow indicates the general soil water content following the transitions of seasons. The lighter blue color indicates more intense drought limitations for vegetation growth. The gray arrow indicates an unclear process. See also Reichstein et al. (2013) for the effects of extreme events on terrestrial ecosystems.
), respectively. The relative importance of the impacts and the relative warming rates between seasons are indicated by the line width (broader lines indicate greater importance). The blue arrow indicates the general soil water content following the transitions of seasons. The lighter blue color indicates more intense drought limitations for vegetation growth. The gray arrow indicates an unclear process. See also Reichstein et al. (2013) for the effects of extreme events on terrestrial ecosystems.
In addition to the divergent responses of the AVG to mean  and
and  in parts of the mid- and high-latitude NH, the AVG responds differently to
in parts of the mid- and high-latitude NH, the AVG responds differently to  than to
than to  and
and  across most of the temperate NH. Significant positive correlations between AVG and
across most of the temperate NH. Significant positive correlations between AVG and  are observed in nearly twice as many areas as those of
are observed in nearly twice as many areas as those of  and
and  . Early and mid-growing season (e.g., spring and early summer) warming tends to trigger regional droughts (Supplementary Fig. S14-S16), which can exert strong control on vegetation growth and even cause tree mortality in the temperate NH and some parts of the cold NH28,30,31,32. Nevertheless, the emergence of a positive response of
. Early and mid-growing season (e.g., spring and early summer) warming tends to trigger regional droughts (Supplementary Fig. S14-S16), which can exert strong control on vegetation growth and even cause tree mortality in the temperate NH and some parts of the cold NH28,30,31,32. Nevertheless, the emergence of a positive response of  to
to  in autumn may reflect alleviation of water stress on photosynthetic activity in that period (Figs 1 and 4, Supplementary Figs S1, S14)33, which is confirmed by a generally negative correlation between
in autumn may reflect alleviation of water stress on photosynthetic activity in that period (Figs 1 and 4, Supplementary Figs S1, S14)33, which is confirmed by a generally negative correlation between  and the autumn WAI (Fig. 1). Consequently, alleviation of water limitations in autumn (Fig. 4, Supplementary Fig. S14) results in a generally positive correlation between the TRI and
and the autumn WAI (Fig. 1). Consequently, alleviation of water limitations in autumn (Fig. 4, Supplementary Fig. S14) results in a generally positive correlation between the TRI and  in ~49% of the temperate NH except in portions of arid regions (e.g., the central U.S.A.). By contrast, a predominantly (~60–70%) negative correlation exists between the TRI and
in ~49% of the temperate NH except in portions of arid regions (e.g., the central U.S.A.). By contrast, a predominantly (~60–70%) negative correlation exists between the TRI and  and
and  (Fig. 1, Supplementary Figs S1, S13). These factors also lead to a conversion from a generally (~55–81%) negative correlation between
(Fig. 1, Supplementary Figs S1, S13). These factors also lead to a conversion from a generally (~55–81%) negative correlation between  and
and  and
and  to a positive correlation (in ~57–67% of the temperate NH) between
to a positive correlation (in ~57–67% of the temperate NH) between  and
and  in the temperate NH (Supplementary Fig. S8a–c).
in the temperate NH (Supplementary Fig. S8a–c).
The positive responses of the AVG to variations in the mean nighttime temperatures  and
and  in parts of NH are most likely attributable to the reported compensation effects of nocturnal warming34. However, the shifting responses of the AVG to
in parts of NH are most likely attributable to the reported compensation effects of nocturnal warming34. However, the shifting responses of the AVG to  and
and  in arid and temperate arid regions (Fig. 1, Supplementary Fig. S1, S3 and Table S2) imply that the compensation effects of nocturnal warming on vegetation growth might be seasonally and spatially dependent and are likely modified by other bio-physical factors, such as water conditions (Fig. 4) and plant functional types35. In contrast to the ambiguous relationships between the AVG and
in arid and temperate arid regions (Fig. 1, Supplementary Fig. S1, S3 and Table S2) imply that the compensation effects of nocturnal warming on vegetation growth might be seasonally and spatially dependent and are likely modified by other bio-physical factors, such as water conditions (Fig. 4) and plant functional types35. In contrast to the ambiguous relationships between the AVG and  and
and  , there is a pervasive negative response of the AVG, from the perspective of
, there is a pervasive negative response of the AVG, from the perspective of  and
and  , to
, to  across most of the temperate and cold NH except in certain mountainous regions (e.g., the Rocky Mountains in western North America). The dominant effect of autumn nocturnal warming on NH vegetation growth may be enhanced plant respiration, which consequently leads to autumn carbon loss36. However, the TRI data indicate a persistent positive rather than negative correlation with
across most of the temperate and cold NH except in certain mountainous regions (e.g., the Rocky Mountains in western North America). The dominant effect of autumn nocturnal warming on NH vegetation growth may be enhanced plant respiration, which consequently leads to autumn carbon loss36. However, the TRI data indicate a persistent positive rather than negative correlation with  (Fig. 1). In addition to enhanced whole-plant respiration, warmer autumn (both day and night) temperatures may reactivate cambial activity37. Combined with alleviation of water limitations (Fig. 4, Supplementary Figs S14-S17), these process may partially explain the observed widespread positive correlation between the TRI and
(Fig. 1). In addition to enhanced whole-plant respiration, warmer autumn (both day and night) temperatures may reactivate cambial activity37. Combined with alleviation of water limitations (Fig. 4, Supplementary Figs S14-S17), these process may partially explain the observed widespread positive correlation between the TRI and  in the temperate NH. Overall, the observed divergent responses of AVG to seasonal
in the temperate NH. Overall, the observed divergent responses of AVG to seasonal  in portions of the NH are to some degree attributable to the water-mediated counterbalancing effects of
in portions of the NH are to some degree attributable to the water-mediated counterbalancing effects of  on vegetation growth and respiration34,38,39.
on vegetation growth and respiration34,38,39.
In summary, remote sensing retrievals, empirical up-scaling measurements from FLUXNET and tree growth data consistently indicate a possible water-mediated divergent response of vegetation activity to seasonal VDNC, especially in temperate regions and boreal regions with dry summers in the NH. Our research provides new insights into the seasonally divergent responses of the AVG to the seasonal asymmetry in the VDNC and reinforces the idea that the projected intensification in the asymmetry in the seasonal VDNC under continued global warming and increasing climate extremes40 may have strong, spatially variable effects on vegetation growth and carbon dynamics in the NH, with associated changes in regional ecosystem services, the terrestrial carbon cycle and global climate feedbacks.
Methods
Remote sensing data and up-scaled GPP
We used three types of data to investigate how AVG (i.e., vegetation growth during the growing season) responded to the mean seasonal VDNC: the satellite-derived AVHRR/GIMMS Normalized Difference Vegetation Index (NDVI, specifically version NDVI3g)18,19, the satellite-derived Fraction of Absorbed Photosynthetically Active Radiation (FAPAR)20 and FLUXNET-based up-scaled gross primary production (GPP) data21. The data featured a spatial resolution of 0.5° × 0.5° and a monthly temporal resolution for 1982–2008. Pixels with a multi-year mean annual NDVI value <0.10 were masked in our study.
Tree ring data
In total, 149 standard TRI chronologies from the mid- and high-latitude NH were used to investigate the interannual responses of tree growth to the seasonal VDNC. Standard TRI chronologies were built following the standard procedures of dendrochronology41, including fitting a curve to the raw ring-width series and then dividing each ring-width value by the corresponding curve value to generate a series of growth indices. Of these chronologies, 136 were obtained from the International Tree-Ring Data Bank (ITRDB, http://www.ncdc.noaa.gov/data-access/paleoclimatology-data/datasets/tree-ring) based on four criteria: 1) intact records of latitude, longitude, elevation, species and sample depth; 2) the chronologies cover at least 1950–1998 or extend to more recent times (i.e., early 21st century); 3) the sample depth for each site-year is greater than 10; and 4) the altitude of the sample site is less than the 90th percentile of the elevation distribution of the surrounding 0.5° × 0.5° region based on the GTOPO30 dataset (with ~1 km resolution, available from U.S. Geological Survey, https://lta.cr.usgs.gov/GTOPO30) to match the regional vegetation growth patterns and avoid site-specific representations. Our research group provided another 13 standard chronologies from northern China. Our chronologies satisfy the four criteria listed above (see Table S4 for details).
Climate data
Monthly mean maximum ( , averaging daily maximum temperature) and mean minimum temperature (
, averaging daily maximum temperature) and mean minimum temperature ( , averaging daily minimum temperature) at a spatial resolution of 0.5° were obtained from the newly updated CRU TS 3.22 dataset (http://www.cru.uea.ac.uk/). The seasonal (in detail see section of sensitivity test) mean
, averaging daily minimum temperature) at a spatial resolution of 0.5° were obtained from the newly updated CRU TS 3.22 dataset (http://www.cru.uea.ac.uk/). The seasonal (in detail see section of sensitivity test) mean  and
and  were then calculated by averaging the monthly mean climate data. The seasonal mean
were then calculated by averaging the monthly mean climate data. The seasonal mean  and
and  are used in this study to represent the general climate conditions of seasonal mean daytime and nighttime temperature3, respectively. Remotely sensed soil moisture data with a daily resolution for the period 1978–2013 were obtained from the Essential Climate Variable Soil Moisture dataset (ECV SM 02.0) (http://www.esa-soilmoisture-cci.org/). These data were produced following the method described by Liu et al. (2011) and Liu et al. (2012)42,43, representing surface soil moisture with a global coverage and spatial resolution of 0.25°. We then resampled this surface soil moisture data to a spatial resolution of 0.5° × 0.5°. Unfortunately, the soil moisture data were not continuous during 1982–2008 and only pixels with at least 19 years of data between 1982 and 2008 were retained.
are used in this study to represent the general climate conditions of seasonal mean daytime and nighttime temperature3, respectively. Remotely sensed soil moisture data with a daily resolution for the period 1978–2013 were obtained from the Essential Climate Variable Soil Moisture dataset (ECV SM 02.0) (http://www.esa-soilmoisture-cci.org/). These data were produced following the method described by Liu et al. (2011) and Liu et al. (2012)42,43, representing surface soil moisture with a global coverage and spatial resolution of 0.25°. We then resampled this surface soil moisture data to a spatial resolution of 0.5° × 0.5°. Unfortunately, the soil moisture data were not continuous during 1982–2008 and only pixels with at least 19 years of data between 1982 and 2008 were retained.
Water deficit and WAI
The monthly water deficit was estimated using a simple water balance equation:
 where
where  ,
,  and
and  are the water deficit (positive values indicate water surpluses), precipitation and potential evapotranspiration in month i. The potential evapotranspiration is calculated using Thornthwaite’s equation44. Then, the total seasonal water deficit is calculated by summing the monthly WD values. Negative values express the degree of dryness; the more negative the value, the more intense the water limitation. In our study, the monthly WAI was calculated based on the water balance equation proposed by Kleidon and Heimann (1998)45 and was used to represent the soil moisture conditions.
are the water deficit (positive values indicate water surpluses), precipitation and potential evapotranspiration in month i. The potential evapotranspiration is calculated using Thornthwaite’s equation44. Then, the total seasonal water deficit is calculated by summing the monthly WD values. Negative values express the degree of dryness; the more negative the value, the more intense the water limitation. In our study, the monthly WAI was calculated based on the water balance equation proposed by Kleidon and Heimann (1998)45 and was used to represent the soil moisture conditions.
Statistical analysis
Spearman partial correlation and ridge regression were performed to investigate the responses of vegetation activity in the mid- and high-latitude NH to the mean seasonal VDNC. The interannual sensitivity of vegetation growth to IAV in the seasonal VDNC was evaluated using the ridge regression coefficients. Ridge regression has been demonstrated to be an effective method for addressing collinearity in multivariate regression46. The probability density function (PDF) of the Spearman partial correlation coefficients between the seasonal VDNC and the IAV in the AVG was estimated for seven major climate zones (as defined by the Köppen–Geiger climate classification47), including arid (RegAR, grouping of BWk, BWh, BSk and BSh), warm temperate humid (RegTH, grouping of Cfa, Cfb and Cfc), warm temperate arid (RegTA, grouping of Csa, Csb, Csc, Cwa, Cwb and Cwc), cold humid (RegAH, grouping of Dfa, Dfb, Dfc and Dfd), cold summer dry (RegAS, grouping of Dsa, Dsb, Dsc and Dsd), cold winter dry (RegAW, grouping of Dwa, Dwb, Dwc, Dwd) and polar tundra (RegET). Prior to performing the Spearman partial correlation and ridge regression analyses, all variables (i.e., NDVI, FAPAR, GPP, TRI and the climate variables) were detrended using a linear function. The statistical analyses and figure creation were performed using the MATLAB software package (R2012b).
Sensitivity test
In this study, we attempted to define the seasons (spring, summer and autumn) in a manner that is consistent across the mid- and high-latitude NH. Notably, three different definitions of seasons were used in our study to verify the robustness of our conclusions relative to the arbitrary definitions of vegetation growth seasons in the mid- and high-latitude NH. The three definitions are as follows:
Definition 1: spring: April-May; summer: June-August; autumn: September-October
Definition 2: spring: April-May; summer: June-August; autumn: September
Definition 3: spring: May; summer: June-August; autumn: September-October
We then performed the same analyses for the three different season definitions. The results of Definition 1 are presented in the main text and the results of the other two definitions are presented in the supplementary materials.
Additional Information
How to cite this article: Wu, X. et al. Seasonal divergence in the interannual responses of Northern Hemisphere vegetation activity to variations in diurnal climate. Sci. Rep. 6, 19000; doi: 10.1038/srep19000 (2016).
References
Myneni, R., Keeling, C., Tucker, C., Asrar, G. & Nemani, R. Increased plant-growth in the northern high-latitudes from 1981 to 1991. Nature 386, 698–702 (1997).
Nemani, R. R. et al. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300, 1560–1563 (2003).
Peng, S. et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 501, 88–92 (2013).
Keeling, C., Chin, J. & Whorf, T. Increased activity of northern vegetation inferred from atmospheric CO2 measurements. Nature 382, 146–149 (1996).
Reay, D. S., Dentener, F., Smith, P., Grace, J. & Feely, R. A. Global nitrogen deposition and carbon sinks. Nature Geoscience 1, 430–437 (2008).
Piao, S. et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nature Communications 5, doi: 10.1038/ncomms6018 (2014).
Braganza, K., Karoly, D. J. & Arblaster, J. Diurnal temperature range as an index of global climate change during the twentieth century. Geophysical Research Letters 31, doi: 10.1029/2004GL019998, L13217 (2004).
Easterling, D. R. et al. Maximum and minimum temperature trends for the globe. Science 277, 364–367 (1997).
Karl, T. et al. A new perspective on recent global warming: Asymmetric trends of daily maximum and minimum temperature. Bulletin of the American Meteorological Society 74, 1007–1023 (1993).
IPCC. Climate change 2013: The physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. 1535 pp (Cambridge, United Kingdom and Now York, NY, USA, 2013).
Xu, L. et al. Temperature and vegetation seasonality diminishment over northern lands. Nature Climate Change 3, 581–586 (2013).
Sun, D., Kafatos, M., Pinker, R. T. & Easterling, D. R. Seasonal variations in diurnal temperature range from satellites and surface observations. IEEE Transactions on Geoscience and Remote Sensing 44, 2779–2785 (2006).
Betts, A. K., Desjardins, R., Worth, D. & Cerkowniak, D. Impact of land use change on the diurnal cycle climate of the Canadian Prairies. Journal of Geophysical Research: Atmospheres 118, doi: 10.1002/2013jd020717 (2013).
Chou, C. et al. Increase in the range between wet and dry season precipitation. Nature Geoscience 6, 263–267 (2013).
Bonan, G. B. Forests and Climate Change: Forcings, Feedbacks and the Climate Benefits of Forests. Science 320, 1444–1449 (2008).
Angert, A. et al. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proceedings of the National Academy of Sciences of the United States of America 102, 10823–10827 (2005).
Wang, X. et al. Spring temperature change and its implication in the change of vegetation growth in North America from 1982 to 2006. Proceedings of the National Academy of Sciences of the United States of America 108, 1240–1245 (2011).
Tucker, C. J. et al. An extended AVHRR 8‐km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. International Journal of Remote Sensing 26, 4485–4498 (2005).
Pinzon, J. E. & Tucker, C. J. A non-stationary 1981–2012 AVHRR NDVI3g time series. Remote Sensing 6, 6929–6960 (2014).
Zhu, Z. et al. Global data sets of vegetation leaf area index (LAI) 3 g and Fraction of Photosynthetically Active Radiation (FPAR) 3 g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sensing 5, 927–948 (2013).
Jung, M. et al. Global patterns of land‐atmosphere fluxes of carbon dioxide, latent heat and sensible heat derived from eddy covariance, satellite and meteorological observations. Journal of Geophysical Research: Biogeosciences 116, G00J07, doi: 10.1029/2010JG001566 (2011).
Beer, C. et al. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science 329, 834–838 (2010).
Lucht, W. et al. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science 296, 1687–1689 (2002).
Zhao, M. & Running, S. W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 329, 940–943 (2010).
Barber, V. A., Juday, G. P. & Finney, B. P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 405, 668–673 (2000).
Zhang, X., Tarpley, D. & Sullivan, J. T. Diverse responses of vegetation phenology to a warming climate. Geophysical Research Letters 34, L19405, doi: 10.1029/2007GL031447 (2007).
Turnbull, M., Murthy, R. & Griffin, K. The relative impacts of daytime and night‐time warming on photosynthetic capacity in Populus deltoides. Plant, Cell & Environment 25, 1729–1737 (2002).
Liu, H. et al. Rapid warming accelerates tree growth decline in semi‐arid forests of Inner Asia. Global Change Biology 19, 2500–2510 (2013).
Breshears, D. D. et al. Regional vegetation die-off in response to global-change-type drought. Proceedings of the National Academy of Sciences of the United States of America 102, 15144–15148 (2005).
Allen, C. D. et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259, 660–684 (2010).
Williams, A. P. et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nature Climate Change 3, 292–297 (2013).
Peng, C. et al. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nature Climate Change 1, 467–471 (2011).
Vaz, M. et al. Drought-induced photosynthetic inhibition and autumn recovery in two Mediterranean oak species (Quercus ilex and Quercus suber). Tree Physiology 30, 946–956 (2010).
Wan, S., Xia, J., Liu, W. & Niu, S. Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration. Ecology 90, 2700–2710 (2009).
Alward, R. D., Detling, J. K. & Milchunas, D. G. Grassland vegetation changes and nocturnal global warming. Science 283, 229–231 (1999).
Piao, S. et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 451, 49–52 (2008).
De Luis, M., Gricar, J., Cufar, K. & Raventos, J. Seasonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. Iawa Journal 28, 389–404 (2007).
Xia, J., Han, Y., Zhang, Z. & Wan, S. Effects of diurnal warming on soil respiration are not equal to the summed effects of day and night warming in a temperate steppe. Biogeosciences 6, 1361–1370 (2009).
Still, C. Biogeochemistry: As different as night and day. Nature 501, 39–40 (2013).
Reichstein, M. et al. Climate extremes and the carbon cycle. Nature 500, 287–295 (2013).
Cook, E. R. & Kairiukstis, L. A. Methods of dendrochronology: applications in the environmental sciences. (Springer Science & Business Media, 1990).
Liu, Y. et al. Trend-preserving blending of passive and active microwave soil moisture retrievals. Remote Sensing of Environment 123, 280–297 (2012).
Liu, Y. et al. Developing an improved soil moisture dataset by blending passive and active microwave satellite-based retrievals. Hydrology and Earth System Sciences 15, 425–436 (2011).
Thornthwaite, C. W. An approach toward a rational classification of climate. Geographical Review 38, 55–94 (1948).
Kleidon, A. & Heimann, M. A method of determining rooting depth from a terrestrial biosphere model and its impacts on the global water and carbon cycle. Global Change Biology 4, 275–286 (1998).
Hoerl, A. E. & Kennard, R. W. Ridge regression: Biased estimation for nonorthogonal problems. Technometrics 12, 55–67 (1970).
Kottek, M., Grieser, J., Beck, C., Rudolf, B. & Rubel, F. World map of the Koppen-Geiger climate classification updated. Meteorologische Zeitschrift 15, 259–264 (2006).
Acknowledgements
The digital elevation model data, GTOPO30, was obtained from the U.S. Geological Survey. X.L. is supported by the National Natural Science Foundation of China (NSFC No. 41390462 and 41130640). X.W. is supported by the National Natural Science Foundation of China (NSFC No. 41571038) and the Fundamental Research Funds for the Central Universities (Grant No. 2014NT40). H.L. is financially supported by the National Natural Science Foundation of China (NSFC No. 41530747).
Author information
Authors and Affiliations
Contributions
X.W. conceived the idea, designed the paper, performed the numerical analysis and led the interpretation of the results. H.L. and E.L. contributed parts of the tree growth increment data. X.L., P.B. and H.L. assisted with selection of the main results for presentation. X.W., H.L., X.L., E.L., P.B. and Y.H. contributed to interpreting the results and writing the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wu, X., Liu, H., Li, X. et al. Seasonal divergence in the interannual responses of Northern Hemisphere vegetation activity to variations in diurnal climate. Sci Rep 6, 19000 (2016). https://doi.org/10.1038/srep19000
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep19000
This article is cited by
-
Seasonal compensation implied no weakening of the land carbon sink in the Northern Hemisphere under the 2015/2016 El Niño
Science China Earth Sciences (2024)
-
Enhanced growth after extreme wetness compensates for post-drought carbon loss in dry forests
Nature Communications (2019)
-
Long-term spatiotemporal variations in satellite-based soil moisture and vegetation indices over Iran
Environmental Earth Sciences (2019)
-
Recent asymmetric warming trends of daytime versus nighttime and their linkages with vegetation greenness in temperate China
Environmental Science and Pollution Research (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
