
Abstract
According to classical sexual selection theory, complex multimodal courtship displays have evolved in males through female choice. While it is well-known that socially monogamous songbird males sing to attract females, we report here the first example of a multimodal dance display that is not a uniquely male trait in these birds. In the blue-capped cordon-bleu (Uraeginthus cyanocephalus), a socially monogamous songbird, both sexes perform courtship displays that are characterised by singing and simultaneous visual displays. By recording these displays with a high-speed video camera, we discovered that in addition to bobbing, their visual courtship display includes quite rapid step-dancing, which is assumed to produce vibrations and/or presumably non-vocal sounds. Dance performances did not differ between sexes but varied among individuals. Both male and female cordon-bleus intensified their dance performances when their mate was on the same perch. The multimodal (acoustic, visual, tactile) and multicomponent (vocal and non-vocal sounds) courtship display observed was a combination of several motor behaviours (singing, bobbing, stepping). The fact that both sexes of this socially monogamous songbird perform such a complex courtship display is a novel finding and suggests that the evolution of multimodal courtship display as an intersexual communication should be considered.
Similar content being viewed by others
Introduction
Elaborate courtship displays are assumed to have evolved under strong sexual selection pressure in males1,2. Males of polygynous species (e.g., spiders3, frogs4, fishes5 and birds6,7) use multimodal courtship displays to increase the efficacy of signalling8,9. Thus, by coordinating visual and acoustic displays without interference between display components, polygynous male birds can better convey sexual signals6,7,10.
Much research has focussed on male–female directed courtship displays performed by polygynous male birds, while the occurrence of both male–female and female–male directed courtship displays performed by socially monogamous birds has often been overlooked. Elaborate mutual dance displays between sexes are known to be performed by socially monogamous non-passerine birds, which are non-vocal learners, such as grebes11. In socially monogamous non-passerine birds, dance duets may serve similar functions as vocal duets do in songbirds12: contributing to pair formation, pair bonding, or mate guarding13. The evolution and mechanisms of dance duets have received far less attention than have vocal duets. Furthermore, why a few songbird species use both song and dance displays as intersexual communication is a puzzling question14.
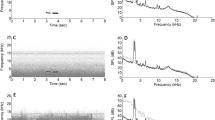
The blue-capped cordon-bleu (Uraeginthus cyanocephalus), a socially monogamous songbird, is one of the few species in which both sexes perform courtship displays. During these displays, they hold a piece of nesting material, then bob up and down and sing15 (Supplementary Video S1). Notably, the courtship bobbing produces rhythmical sounds (Supplementary Video S1, Supplementary Fig. S1). Here, we analysed the audio-visual displays of the blue-capped cordon-bleu in detail using high-speed video-camera recordings (Fig. 1a). As a result, we discovered that their visual courtship display includes quite rapid step-dancing during bobbing (see results). This specific “tap-dance” like behaviour has never been reported in songbirds and presumably produces non-vocal sounds and/or vibrations in addition to song.

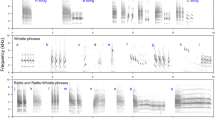
Courtship display in blue-capped cordon-bleus.
(a) When blue-capped cordon-bleus perform courtship display, (b) they simultaneous bob and step and (c) sing at certain times. Bobbing (shaded bars) with multiple steps (white bars) is repeated often and usually singing occurs several times. White and shaded arrows in (b) correspond with the colour of bars in (c). Note the number of steps in (c) involved in each bobbing action.
We predicted that such a physically demanding dance display plays an important role in intersexual communication as well as song in cordon-bleus. As a first step toward understanding the function of the stepping-dance display in both male and female blue-capped cordon-bleus, we investigated between- and within- individual variations in dance performance. We expected that male performances would be more exaggerated than those of females, as is observed in plumage15 and in song16,17. We also expected that to maximize signal efficacy cordon-bleus adjust their dance performances depending on their partner’s position and song6,8.
Results
(a) Courtship dance
High-speed video recordings revealed that in a single bobbing motion, birds hop with their heads pointed upwards and stamp their feet several times so rapidly that it is invisible to the naked eye (Supplementary Video S1, Fig. 1b). We confirmed that all cordon-bleus that performed courtship displays (7 males and 4 females) included rapid steps while bobbing (Fig. 1, Supplementary Video S1). The birds always performed the displays on the perch (Fig. 1a). On average, birds performed 3.17 steps (SEM, 0.03; range, 0–6) per bobbing action. One step took 6–12 frames of 300-frames/s video, thus equalling 20–40 ms, at an estimated rate of 25 Hz to 50 Hz. The bobbing tempo was 1.39 bobs/s (SEM, 0.02).
(b) Sex and individual differences
Dance probability, bobbing tempo and the number of steps during bobs did not differ significantly between males and females (Table 1a, Supplementary Video S1), although males danced more often (males: 24.8%, females: 10.6%; see Supplementary Table S1a for detailed information) and tended to bob more quickly and take more steps (Fig. 2a,b; Table 1b,c). Variations in dance probability and the number of steps differed significantly between individuals (p < 0.05, Table 1a,c), but bobbing tempo did not (p = 0.425, Table 1b). Although each subject chose to dance to one or two particular individuals, the birds exhibiting greater motor performance did not tend to receive more dance displays (Supplementary Table S1b).

Changes of dance performance within and among individuals.
Individual differences were calculated in (a) bobbing tempo and (b) the number of steps per bob (shaded bars: males, white bars: females). Bobbing tempo depended on (c) whether the partner was on the same perch and (d) whether birds were singing. Similarly, the number of steps depended on (e) whether the partner was on the same perch and (f) whether birds were singing. (a–f) All box plots show median, quartiles and minimum and maximum values. (c–f) Mean individual values (closed circles: males, open circles: females) and their within-individual changes are indicated.
(c) Within-individual changes
Both males and females bobbed more quickly and took more steps when their partners were present on the same perch (p < 0.001; Table 1b,c; Fig. 2c,e), which was likely caused by the presence of their nearby partners (see Supplementary Video S2). Individuals also adjusted the number of steps and the bobbing tempo when singing, bobbing faster (p < 0.001, Table 1b, Fig. 2d) and taking fewer steps (p < 0.001, Table 1c, Fig. 2f).
Discussion
Both male and female blue-capped cordon-bleus exhibited quite rapid stepping behaviour during courtship displays, which varied among individuals but not across sexes. The evolution of this complex courtship display in both sexes is surprising because male ornaments tend to become exaggerated even in monogamous species because females are generally the choosier sex1. According to our pairing experiment, both male and female cordon-bleus chose to court with particular individuals, but high-motor performance individuals were not necessarily popular among the opposite sex (Supplementary Table S1b). Assortative mating did not tend to occur with respect to dance performances. The only sex difference in temporal pattern of the blue-capped cordon-bleus courtship display is the slightly shorter song produced by females17. Even if the small sample size prevented us from finding sex-related differences in dance performance, such sex differences would likely be small because the distribution of dance performances overlapped between males and females (Fig. 2a,b). Although other estrildid finches are known to show both dance and song courtship display14, the cordon-bleus displays (blue-capped and red-cheeked cordon-bleus; Supplementary Video S1, S3) are exceptional because as far as we know, theirs are the only dances that produce acoustic and vibratory signals and that are performed by both sexes.
Performing rapid stepping behaviour seemed to enable male and female cordon-bleus to communicate via multiple modalities. Our results suggest that both sexes produce multimodal (acoustic, visual and tactile) signals for intersexual communication that involves the coordination of several motor systems that control singing, bobbing, stepping and beak movements. Attention should be paid to where the birds dance in the wild because cordon-bleus reportedly dance also on the ground15. However, all dance displays were performed on perches in this captive study. Intensified stepping performance when partners were on the same perch likely enables performers to send vibration signals context-dependently. Considering that the limit of flicker fusion frequency is usually over 100 Hz in many birds18, stepping movements between 25 and 50 Hz could be visible and function as a visual signal for cordon-bleus. The bobbing movements seem to exaggerate the sexually dimorphic plumage of their heads and nesting materials that they hold. Intensified bobbing performance when partners stayed on the same perch could make them more visually appealing to highly motivated partners. Fewer steps and more frequent bobbing during singing are likely adaptations that help avoid interference between signals (i.e., stepping sound, song and body movements). Such coordination could be caused either by physical constraints associated with body movements (e.g., brown-headed cowbirds10), or by deliberately adjusting two independent signals. The amazing courtship display of cordon-bleus had features that are analogous to courtship displays in manakins, particularly with regard to multimodality and acrobatic movements19,20,21, but these species differ in vocal learning ability, behavioural mutuality and mating systems.
The most striking aspect of our findings is that they appear to produce two acoustic signals (non-vocal sounds and songs) simultaneously (Supplementary Video S1, S3, Supplementary Figure S1). Past research showed that some species spontaneously synchronize their movements with musical rhythms to which they are exposed22,23,24,25. However, little is known about how animals can temporally coordinate naturally produced multicomponent acoustic signals (i.e., vocal and non-vocal sounds26). Fewer steps during singing can be explained from the perspective of motor constraints because dance displays and singing are both physically demanding27. In contrast, increased bobbing tempo cannot be explained by trade-offs between singing and dancing signals. Sound pressure and the timing of steps seemed to be coordinated with the songs. A next step in the study of blue-capped cordon-bleus courtship display should be to investigate how singing, bobbing and stepping behaviours are coordinated within individuals and between partners. This would yield insights into how multimodal and multicomponent courtship display evolved.
Material & Methods
Procedure
We used 16 blue-capped cordon-bleus (8 females and 8 males). A male and a female were randomly paired, housed together in a cage (120 × 48 × 48 cm) and their behaviour recorded in a 2-hour recording session (2.02 ± 0.40 hours) with normal (Q3HD ZOOM, 30 frames/s) and high-speed (GC-PX1 Victor, 300 frames/s) video cameras. If either member of a pair failed to dance, we repeated the recording session using the same pair, or switched their partners. The observation time for each individual was arbitrary (see Supplementary Table S1) because we wanted to maximize the sample size by allowing birds to participate as often as they would. We recorded 102 sessions (206.5 hours of footage).
We analysed the video using Adobe Premiere Pro software (Adobe Systems Inc.) and marked three behavioural measures: (i) the occurrence of dance displays for each individual in each session; (ii) the bobbing tempo (number of bobs/s); and (iii) the number of steps in one bobbing action (Fig. 1c). We examined how the sex of the performers, their partners’ positions (same perch or not) and the act of singing while bobbing affected performance on these three metrics.
All individuals were kept on a 14:10 h light:dark schedule (lights on 07:00–21:00) at about 22 °C and 50% humidity. Birds were provided with finch seed mixture, cuttlebone, water and cucumber ad libitum. Nests and nesting materials were always available in the cages. Procedures were in accordance with national laws and approved by the Government of Upper Bavaria.
Statistical analyses
To examine the effect of sex on the probability that an individual danced during sessions, we used a generalized linear mixed-effect model (GLMM) with a binomial distribution. To examine the effects of sex, partners’ position and singing behaviour on dance performance, we used a GLMM with a Poisson distribution to analyse the number of steps and a linear mixed-effect (LME) model to analyse the bobbing tempo. In these analyses, we considered session number nested within bird ID as random effects to control for non-independence of the data. To investigate individual differences in dance display, we used a likelihood ratio test to compare this model with a model in which bird ID was removed. All statistical analyses were performed using R 3.1.2 (R development Core Team 2014).
Additional Information
How to cite this article: Ota, N. et al. Tap dancing birds: the multimodal mutual courtship display of males and females in a socially monogamous songbird. Sci. Rep. 5, 16614; doi: 10.1038/srep16614 (2015).
References
Andersson, M. Sexual selection. (Princeton University Press, Princeton, 1994).
Candolin, U. The use of multiple cues in mate choice. Biol. Rev. Camb. Philos. Soc. 78, 575–595 (2003).
Girard, M. B., Kasumovic, M. M. & Elias, D. O. Multi-modal courtship in the peacock spider, Maratus volans (O.P.-Cambridge, 1874). PLoS One 6, e25390 (2011).
De Luna, A. G., Hödl, W. & Amézquita, A. Colour, size and movement as visual subcomponents in multimodal communication by the frog Allobates femoralis. Anim. Behav. 79, 739–745 (2010).
Amorim, M. C. P., Simões, J. M., Fonseca, P. J. & Turner, G. F. Species differences in courtship acoustic signals among five Lake Malawi cichlid species (Pseudotropheus spp.). J. Fish Biol. 72, 1355–1368 (2008).
Cooper, B. G. & Goller, F. Multimodal signals: enhancement and constraint of song motor patterns by visual display. Science 303, 544–546 (2004).
Dalziell, A. H. et al. Dance choreography is coordinated with song repertoire in a complex avian display. Curr. Biol. 23, 1132–1135 (2013).
O’Loghlen, A. L. & Rothstein, S. I. It’s not just the song: male visual displays enhance female sexual responses to song in brown-headed cowbirds. Condor 112, 615–621 (2010).
Uetz, G. W., Roberts, J. A. & Taylor, P. W. Multimodal communication and mate choice in wolf spiders: female response to multimodal versus unimodal signals. Anim. Behav. 78, 299–305 (2009).
Hoepfner, A. R. & Goller, F. Atypical song reveals spontaneously developing coordination between multi-modal signals in brown-headed cowbirds (Molothrus ater). PLoS One 8, e65525 (2013).
Nuechterlein, G. L. & Storer, R. W. The pair-formation displays of the western grebe. Condor 84, 350–369 (1982).
Hall, M. L. & Magrath, R. D. Temporal coordination signals coalition quality. Curr. Biol. 17, R406–R407 (2007).
Malacarne, G., Cucco, M. & Camanni, S. Coordinated visual displays and vocal duetting in different ecological situations among Western Palearctic non-passerine birds. Ethol. Ecol. Evol. 3, 207–219 (1991).
Soma, M. & Garamszegi, L. Z. Evolution of courtship display in estrildid finches: dance in relation to female song and plumage ornamentation. Front. Ecol. Evol. 3, 4 (2015).
Goodwin, D. Estrildid finches of the world. (Cornell University Press, Ithaca, 1982).
Geberzahn, N. & Gahr, M. Song learning in male and female (Uraeginthus cyanocephalus), a tropical songbird species. J. Comp. Psychol. 127, 352–364 (2013).
Geberzahn, N. & Gahr, M. Undirected (solitary) birdsong in female and male blue-capped cordon-bleus (Uraeginthus cyanocephalus) and its endocrine correlates. PLoS One 6, e26485 (2011).
Jones, M. P., Pierce, K. E. & Ward, D. Avian vision: a review of form and function with special consideration to birds of prey. J. Exot. Pet Med. 16, 69–87 (2007).
Bostwick, K. S. & Prum, R. O. Courting bird sings with stridulating wing feathers. Science. 309, 736 (2005).
DuVal, E. H. Cooperative display and lekking behavior of the lance-tailed M manakin (Chiroxiphia Lanceolata). Auk 124, 1168–1185 (2007).
Fusani, L. & Schlinger, B. A. Proximate and ultimate causes of male courtship behavior in golden-collared manakins. J. Ornithol. 153, S119–S124 (2012).
Hasegawa, A., Okanoya, K., Hasegawa, T. & Seki, Y. Rhythmic synchronization tapping to an audio-visual metronome in budgerigars. Sci. Rep. 1, 120 (2011).
Patel, A. D., Iversen, J. R., Bregman, M. R. & Schulz, I. Experimental evidence for synchronization to a musical beat in a nonhuman animal. Curr. Biol. 19, 827–830 (2009).
Hattori, Y., Tomonaga, M. & Matsuzawa, T. Spontaneous synchronized tapping to an auditory rhythm in a chimpanzee. Sci. Rep. 3, 1566 (2013).
Cook, P., Rouse, A., Wilson, M. & Reichmuth, C. A California sea lion (Zalophus californianus) can keep the beat: motor entrainment to rhythmic auditory stimuli in a non vocal mimic. J. Comp. Psychol. 127, 412–427 (2013).
Soma, M. & Mori, C. The songbird as a percussionist: syntactic rules for non-vocal sound and song production in Java sparrows. PLoS One 10, e0124876 (2015).
Fusani, L., Barske, J., Day, L. D., Fuxjager, M. J. & Schlinger, B. A. Physiological control of elaborate male courtship: female choice for neuromuscular systems. Neurosci. Biobehav. Rev. 46, 534–546 (2014).
Acknowledgements
We thank the staff of the Max Planck Institute for Ornithology for providing the experimental equipment and for maintaining the birds. This study was supported by JSPS Grants-in-Aid for Young Scientists (22800002, 23680027) to MS, Grant-in-Aid for JSPS Fellows (15J02516) to NO.
Author information
Authors and Affiliations
Contributions
N.O. and M.S. conducted the experiments and analysed the data. All authors designed the experiments and wrote the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Ota, N., Gahr, M. & Soma, M. Tap dancing birds: the multimodal mutual courtship display of males and females in a socially monogamous songbird. Sci Rep 5, 16614 (2015). https://doi.org/10.1038/srep16614
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep16614
This article is cited by
-
Simulating individual movement in fish
Scientific Reports (2023)
-
Song system neuroanatomy, and immediate early gene expression in a finch species with extensive male and female song
Journal of Comparative Physiology A (2023)
-
Homosexual Courtship in Young Wild Capuchin Monkeys: A Case Study
Archives of Sexual Behavior (2023)
-
Star finches Neochmia ruficauda have a visual preference for white dot patterns: a possible case of trypophilia
Animal Cognition (2022)
-
Parameterizing animal sounds and motion with animal-attached tags to study acoustic communication
Behavioral Ecology and Sociobiology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
