
Abstract
Medaka is an ideal model for sex determination and sex reversal, such as XY phenotypically female patients in humans. Here, we assembled improved TALENs targeting the DMY gene and generated XYDMY− mutants to investigate gonadal dysgenesis in medaka. DMY-TALENs resulted in indel mutations at the targeted loci (46.8%). DMY-nanos3UTR-TALENs induced mutations were passed through the germline to F1 generation with efficiencies of up to 91.7%. XYDMY− mutants developed into females, laid eggs and stably passed the YDMY− chromosome to next generation. RNA-seq generated 157 million raw reads from WT male (WT_M_TE), WT female (WT_F_OV) and XYDMY− female medaka (TA_F_OV) gonad libraries. Differential expression analysis identified 144 up- and 293 down-regulated genes in TA_F_OV compared with WT_F_OV, 387 up- and 338 down-regulated genes in TA_F_OV compared with WT_M_TE. According to genes annotation and functional prediction, such as Wnt1 and PRCK, it revealed that incomplete ovarian function and reduced fertility of XYDMY− mutant is closely related to the wnt signaling pathway. Our results provided the transcriptional profiles of XYDMY− mutants, revealed the mechanism between sex reversal and DMY in medaka and suggested that XYDMY− medaka was a novel mutant that is useful for investigating gonadal dysgenesis in phenotypic female patients with the 46, XY karyotype.
Similar content being viewed by others

Introduction
Over two decades have passed since the male determining gene, Sry, was identified1. The SRY gene plays a pivotal role in sex determination: point mutations or deletions of the SRY gene are found in approximately 15% of XY females in humans2,3. XY female syndrome, phenotypic female patients with the XY karyotype, has been studied clinically, cytogenetically, hormonally, endoscopically and histologically4,5,6. However, the transcriptional and post-transcriptional regulation mechanisms of SRY-induced XY female syndrome remain largely unknown. A lack of animal models has meant that it is difficult to perform in-depth studies of the pathological changes and to determine the mechanisms underlying the XY female syndrome.
Sex-determining systems in fish are very diverse, which could be determined by heredity, environment, or both. Meanwhile, the pathway of sex determination can be manipulated by administering exogenous sex steroids7,8,9,10,11. In non-mammalian species, which also have a XX-XY sex-determination system, SRY is not present at all. Until DMY gene of medaka (Oryzias latipes) was identified in the teleost fish12,13, it is believed that the XX-XY sex-determination system was conserved in a wide range of animals, including C. elegans, Drosophila, fish and mammals. Although much is known about the master male sex-determining (SD) gene in medaka12,14,15,16, the precise mechanisms involved in primary sex determination and sex differentiation remain undefined. It is difficult to perform specific gene targeting in medaka, because of the lack of methodologies for homologous recombination and embryonic stem cell derivation, which has impeded its use in male heterogametic (XX-XY) sex determination system studies.
Modifications of genomes have laid the foundation of functional studies in modern biology and have led to significant discoveries17. Recently, Zinc finger nucleases (ZFNs)18,19,20, transcription activator-like effector nucleases (TALENs)21,22,23,24 and clusters of regularly interspaced short palindromic repeats (CRISPR)25,26,27 have been shown to edit genomic DNA in a variety of cell types and different model organisms at stable efficiency and specificity. In our previous studies, we modified the Golden Gate method to disrupt the gene and edit the genome in zebrafish28,29 and medaka30. Here, we directly produced TALEN-induced XYDMY− females in medaka using TALENs. Phenotype of XYDMY− mutant is very similar to human XY female syndrome4,5,6 and SRY KO mouse3,31, especially as several individuals in the population were fertile.
The specificity of targeted sites and the off-target phenomenon are core problems in gene knockout and gene editing research. To test for potential nonspecific mutations induced by TALENs, we also developed a simple and reliable off-target prediction program for confirming TALEN-induced mutation. The TALEN-induced XYDMY− medaka developed into females and laid eggs. Although the sex-determining function of the DMY protein is already recognized; however, as a transcription factor, how does it regulate the downstream factor(s) to control testis differentiation, development and germ cell maturation? To better explain DMY’s regulatory functions as a transcription factor, we performed RNA-seq, a recently developed approach to transcriptome profiling based on deep-sequencing32 and generated 157 million raw reads from WT male (WT_M_TE), WT female (WT_F_OV) and XYDMY− female medaka (TA_F_OV) gonad libraries. These transcriptomic data will contribute to unravel the relationship and mechanism between sex reversal and the DMY gene. Our results suggest that the medaka XYDMY− mutant is a novel mutant line that is useful for investigating XY to XX sex reversal and gonadal dysgenesis in phenotypically female patient with the 46, XY karyotype.
Results
DMY- and DMY-nanos3UTR- TALENs effectively induced DMY gene disruption in Medaka
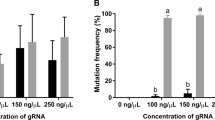
To improve the germline integration efficiency, we incorporated the zebrafish nanos-3′UTR into the TALEN construct (Fig. 1A). Potential TALEN target sites were scanned and designed in the exon of the DMY/DMRT1bY gene (ENSORLT000000025382 and ENSORLT000000025383) (Fig. 1B). We generated TALEN constructs using our previously published method29. The mixture containing a pair of TALEN mRNAs was microinjected into one-cell stage embryos of medaka (Fig. 2A and File S1). 72 hours after microinjection, ten injected embryos were randomly pooled for extracting genomic DNA. As illustrated in Fig. 2B, primers DMY F and DMY R bridge both the effector binding element (EBE) regions, while primers DMY F1 and DMY R link the spacer region and the downstream EBE. If primer DMY F and DMY R generated a 396 bp fragment, while primer DMY F1 and DMY R failed to generate the 167 bp fragment, the result suggested that the targeted gene was disrupted by the TALENs (Fig. 2C). Sequenced PCR positive clones had mutated sequences in the spacer (Fig. 2D). Both DMY-TALENs and DMY-Nanos-3UTR TALENs were effective at disrupting the targeted genes in medaka embryos. Various concentrations (200, 400, 600 and 800 pg) of the TALENs mRNA were microinjected (Fig. 2E,F, File S1 and S2). The targeting efficiency of the DMY-TALENs was good, with a higher TALEN-induced mutation ratio of 42.07% and lower levels of dead and deformed embryos when 600 pg mRNA was microinjected (Fig. 2G,I). Similarly, the targeting efficiency of DMY-nanos-3UTR TALENs was about 41.96% (Fig. 2H,I). Thus, 600 pg was determined as the appropriate concentration in medaka. These results indicated that the TALEN activity was dose-dependent and high-dose microinjection of TALEN mRNA might cause nonspecific and toxic defects in medaka embryos (Fig. 2E–I, File S1 and S2).

The design and assembly of DMY-TALEN sites.
(A) Reconstructions of pCS2- DMY-TALENs -ELD/KKR-nanos UTR plasmids. (B) The location and sequence information of DMY-TALENs. Red uppercase letters indicate the DNA sequence of DMY-TALEN-L sites. Green uppercase letters indicate the complementary paired DNA sequence of DMY-TALEN-R sites. Lowercase letters indicate the spacer region of TALEN sites.

The dosage effects and efficiency evaluation of DMY-TALENs.
(A) Microinjection of TALENs mRNA into medaka. (B) Detection of mutations in TALEN targeted medaka embryos. Primers DMY F, DMY F1 and DMY R were used to amply the DMY gene. Primers DMY F and DMY R bridge both EBE regions, while primers DMY F1 and DMY R link the spacer region and the downstream EBE. DMY F and DMY R generated a 396 bp PCR fragment. DMY F1 and DMY R generated a 167 bp PCR fragment. If primer DMY F and DMY R generated a PCR fragment, while primer DMY F1 and DMY R failed to do so, this suggested that the targeted gene is disrupted by the DMY-TALEN. (C) Electrophoretic detection of mutations in TALEN-injected medaka embryos. Line 1 to 16, TALENs injected embryos. 1,3,6,11,14 show mutated embryos. (D) Sequencing detection of mutations in TALEN-induced medaka embryos. -, deleted nucleotide; lowercase letter, added nucleotide. +, insertions; Δ, deletions. (E) Evaluation of embryonic toxicity of DMY-TALENs. (F) Evaluation of embryonic toxicity of DMY-nanos3UTR-TALENs. (G) The targeting efficiency statistics of DMY- TALENs. (H) The targeting efficiency statistics of DMY-nanos-3UTR TALENs. (I) Comparative analysis of mutation rate and germline transmission rate between DMY-TALENs and DMY-nanos3UTR-TALENs.
Successful germline transmission is essential to establish knockout lines. To evaluate the germline transmission efficiency of the TALEN-mediated gene disruption, ten embryos for DMY and DMY-nanos3UTR from each independent cross were individually collected at 3 days post fertilization (dpf) and genomic DNA was extracted from each cross to assess mutagenesis at the TALEN-targeted site (Fig. 2C,D). 9.02% of DMY-TALEN-induced F1 embryos carried mutations; and 37.56% of DMY-nanos3UTR-TALEN-induced F1 embryos carried mutations in the DMY gene (Fig. 2I and File S2). The higher proportion induced by DMY-nanos3UTR-TALEN indicated that a majority of gametes in the F0 medaka were mutant. These results indicated that there was no significant difference in the targeting efficiency of F0 somatic mutations at the targeted loci (Fig. 2H,I); however, DMY-nanos3UTR-TALEN induced a higher portion of mutations in the germline than DMY-TALENs did.
A novel program to identify potential off-target sites of TALENs
To test for potential nonspecific mutations induced by TALENs, we designed a program to scan the medaka genomic sequence (http://www.ensembl.org/Oryzias_latipes) to identify potential off-target sites potentially targeted by DMY TALENs. Potential off-target sites of DMY-TALENs were searched using the program and 55 candidate sites were identified (File S3). When the spacers were less than 10 bp or more than 24 bp long, the scaffold of TALENs had lower disrupting activity33,34. Five of the 55 candidates had spacers less than 10 bp and 43 of 55 candidates had spacers of more than 24 bp, indicating that it is unlikely that the TALENs could induce mutations at these sites. We analyzed one candidate site (Chr.3: 36,197,357–36,197,403) that had 7-bp mismatches in the recognition sequences and a 12-bp spacer (File S4). PCR amplified the identified potential off-target regions using genomic DNA from TALEN-injected embryos as template; no mutations were found at these sites by DNA sequencing. This result suggested that the novel program could predict the potential off-target sites of TALENs; and that TALENs have high specificity for their target sequences.
Mating scheme of TALENs-induced DMY- mutants and mutant phenotypes
The mating scheme for the TALENs-induced DMY-mutant lines is shown in Fig. 3A. The F0 generation was produced by microinjecting 600 pg DMY-nanos3UTR-TALENs into one-cell stage embryos of medaka; the mutation rate was 46.8% (15/32) (Fig. 3B). The DMY gene knockout medaka could develop into females. 11-bp deletions (named DMYΔ11) and 16-bp insertions (named DMY+16) were identified and chosen to establish mutant lines (Fig. 3C,D). Notably, during the establishment of the mutant lines, two genotyping alleles of DMY gene fragments were identified in individual TALEN-induced F1 mutations using the Li-con 4300 system (Fig. 4A). This result indicated that YDMY−Y male mutants were present in the testcross F1 generation. The genetic males (XY) of the DMY gene mutants, DMYΔ11 and DMY+16 mutant types, developed into females in the F2 generation, which was identified using genomic PCR of the DMY gene (Fig. 4B). To confirm un-expression and dysfunction of DMY gene in the XYDMY− F2 generation, using RT-PCR, expression of DMY gene was not identified in the XYDMY− female medaka (both DMYΔ11 and DMY+16) (Fig. 4C). The first morphological sex difference manifested in the gonads is reflected in the number of germ cells14,35. The number of germ cells in several DMY mutants identified from wild populations resembled that of the female13,16,36. To elucidate sex reversal during the development of XYDMY− mutants, we evaluated the effect of DMY on germ cell number at 5 days after hatching (DAH) in the XYDMY− F3 generation. The XYDMY− mutant fry had more germ cells than that of the WT XY male at 5 DAH (Fig. 4D). This implied sex reversal of XYDMY− mutants took place in early developmental stages and the increased number of germ cells in the XYDMY− mutants may be due to the disruption of DMY gene expression.

Flowchart and establishment of DMY-TALENs-induced mutant lines.
(A) Flowchart of DMY-TALENs-induced mutant lines. (B) The genotypes of TALENs-induced mutations in the F0 generation. (C) The genotypes of TALENs induced F1 founders. (D) The genotypes of TALENs induced F2 mutant lines. Red lowercase letters indicate an additional nucleotide; “-” indicates a deleted nucleotide. +, insertions; Δ, deletions. F, female; M, male.

DMY genotyping, expression and CDS frameshift mutations of DMY mutants.
(A) Genotyping of DMY gene fragments using Li-con 4300 system. 1 to 16, randomly selected individuals in F1 generation. F, female; M, male. (B) Genomic PCR confirmed the DMY gene. M: marker DL2000; 1: WT Male; 2: WT Female; 3: Founder 2; 4: the female mutant from the F2 generation of Founder 2; 5: Founder 3; 6: the female mutant from the F2 generation of Founder 3; Ctrl: no template control. (C) Reverse transcription- PCR with DMY-specific primers. OLA Actin expression was determined for calibration. M: 250 bp Marker; 1. XYDMY− mutant (DMYΔ11); 2. XYDMY− mutant (DMY+16); 3. WT XY male; 4. WT XX female. (D) Numbers of germ cells in mutants and WT medaka fry at 5DAH. XX F: WT XX females; XYDMY− F: XYDMY− matants; XY M: WT XY males. Open circles represent the number of germ cells in individuals (N = 9). **P < 0.001. (E) The CDS frameshift mutations of TALEN-induced DMY gene. WT: CDS sequence of DMY.
According to the amino acid sequence of DMY, DMYΔ11 and DMY+16 are frame-shifted mutant alleles that would produce truncated DMY protein caused by a region of altered translation (Fig. 4E). Thus, an error of the coding sequence has occurred in the mutated DMY gene that resulted in the loss function of the DMY gene. The XYDMY− female or YDMY−Y male (N = 10) from F2 generation that had lost the DMY gene were crossed with the WT to obtain the testcross F3 generation. XX female, XY male, XYDMY− mutant female and YDMY−Y mutant male were identified in the XY female testcross F3 generation. XY males and XYDMY− mutant females were identified in the YDMY−Y male testcross F3 generation. This showed that the DMY gene in the Y chromosome of WT medaka rescued the female phenotype of DMY gene disruption in mutated XY female medaka. Unfortunately, an YDMY−YDMY− mutant female with a genomic homozygous DMY gene mutation on the Y chromosome was never found.
Mature XYDMY− mutants in the F3 generation were obtained for phenotype identification, histological analyses and fluorescence in situ hybridization (FISH) (Fig. 5). There are significant differences between females and males in the size and shape of dorsal and anal fins13,14, which are main part of the secondary morphological sexual characteristics in medaka. The shape of both dorsal and anal fins of XYDMY− sex reversal female were similar to that of the WT XX female, rather than to that of the WT XY male. The size of both dorsal and anal fins of XYDMY− female was significant smaller than those of the WT XY males (Fig. 5A). In addition, ovarian tissue was identified in XYDMY− mutants (Fig. 5B). To confirm the Y chromosome in the TALENs-induced mutants, we analyzed the karyotypes of metaphase cells from the WT male, WT female and XYDMY− female using FISH showing the male specific hybridization signal12. Compared with the two spots in females, three hybridization spots for the specific probe were identified in males (Fig. 5C). The additional FISH signal in males is on the Y chromosome. XYDMY− mutants did not express DMY gene, which were different from WT XY individuals (Fig. 4C). The gonadosomatic index (GSI) and the maturation stages of oocytes are commonly used to evaluate the gonad and gonad development37. At 90 days after hatching, the GSI of XYDMY− mutated female was 10.9 (N = 10); that of the WT female and WT male were 19.1 and 0.7, respectively (Fig. 5D). During the generation of offspring from the XYDMY− mutated female testcross, we found that it was difficult to obtain a sufficient number of offspring. Therefore, we performed the comparative analysis of the number of matured oocytes between the WT female and XYDMY− mutants (N = 10). During 10 days’ embryos collecting, the XYDMY− mutated female produced six eggs per day on average; whereas, the WT XX female produced 20 eggs per day (Fig. 5E). The results from F2 phenotypes revealed that the genetic males (XY) of TALENs-induced DMY gene disruption mutants all developed into females and laid eggs (DMYΔ11 and DMY+16 mutants). Furthermore, histological analyses demonstrated that all XY mutants developed into females; however, there were significant differences in germ cells and gonad (Fig. 5F). The ovary of XYDMY− mutants (12/15) appeared to have fewer oocytes than that of the WT XX female (Fig. 5F). According to the developmental stage of oocytes in medaka, there were equilibrium distributions in each stage of oocytes in the WT XX female ovary, containing cortical alveolar oocytes (CA), vitellogenic oocytes (VO) and mature oocytes (MO). Among 15 XYDMY− mutants (15 mutants were identified in the same genotype, fed in the same tank and sampled at the same time), three did not form mature oocytes, with majority of chromatin nucleolar oocytes (CN), several perinucleolar oocytes (PO) and few CA oocytes in the ovary and no further developmental stage oocytes (Fig. 5F). Twelve of them successfully formed MOs, however, the size of ovary and total number of each developmental stage oocytes were significant smaller and fewer than that of WT XX female (Fig. 5F). This can also explain it is difficult to collect embryos in XYDMY− mutants, even there is sometimes no embryos could be collection (Fig. 5E). The analysis of gonadal histology, GSI and the statistics of mature eggs demonstrated significant differences in the gonadal development and maturity between WT females and XYDMY− mutant females.

Phenotypic identification and analysis of XYDMY− mutants.
(A) Phenotypic diagnostics of the secondary sexual characters in XYDMY− mutants. White dashed area shows the anal fin of medaka. *shows the dosal fin of medaka. (B) The gonad of WT and TA mutants. The red dashed area shows the gonad tissue of medaka. (C) Fluorescence in situ hybridization of the karyotypes of metaphases cell from WT male, WT female and XY mutant female. The pink signal is the male specific hybridization signal. (D) The comparative analyses of gonadosomatic index (GSI) among WT-F, WT-M and XY mutant female (TA-F). *P < 0.5; **P < 0.001. (E) The comparative analyses of oocytes maturation between WT-F and TA-F. (F) Histological analyses of gonad tissue. CN, chromatin nucleolar oocytes; PO, perinucleolar oocytes; CA, cortical alveolar oocytes; EVO, early vitellogenic oocytes; VO, vitellogenic oocytes; LVO, late vitellogenic oocytes; MO, mature oocytes.
Transcriptomic analysis of TALENs-induced DMY- mutants and WT
To better explore the transcriptional regulation function of DMY and identify DMY-related downstream factors that affect the generation, development and maturation of testis or ovary, we used RNA-seq to analyze the transcriptome of WT_M_TE, WT_F_OV and TA_F_OV after 90 days. For both the WT and DMY-mutants, ten samples of gonadal tissues were mixed for library construction. RNA-seq generated 157 million raw reads comprising 10333, 8746 and 8621 transcripts, respectively (Fig. 6A). Using the Ensembl medaka genome database as the reference, 75.8% of the raw reads matched medaka genomic sequences (File S6). Twelve genes were selected randomly from hundreds of different expression transcripts between the gonad of WT and that of DMY mutants. The result of real-time quantitative PCR showed that the trends of these genes expression were the same as in the RNA-seq data (File S7). Thus, the RNA-seq information was accurate and reliable.

Bioinformatic analyses of RNA-seq data.
(A) The number of novel transcripts in the RNA-seq data the WT female (WT_F_OV), the wild-type male (WT_M_Te) and XYDMY− female medaka (TA_F_OV). (B) The differentially expressed transcripts between TA_F_OV and WT_F_OV/WT_M_Te. (C) Correlation of gene expression between WT_M_Te and TA_F_Ov. The up- and downregulated genes are shown in red and green, respectively. Non-differentially expressed genes are shown in blue. (D) Correlation of gene expression between WT_F_Ov and TA_F_Ov. The up- and downregulated genes are shown in red and green, respectively. Non-differentially expressed genes are shown in blue. (E) The cluster of testis-specific expressing transcripts. Cluster analyses of differentially expressed genes among WT_F_Ov, WT_M_Te and TA_F_Ov. The high- and low-expressed genes are shown form yellow to blue, corresponded to the expression level from negative 2 fold to positive 2 fold. (F) The differentially expressed transcripts among TA_F_Ov, WT_F_Ov and WT_M_Te. The up- and downregulated genes in WT_M_Te were shown in Fuchsia and yellow, respectively. The up- and downregulated genes in WT_F_Ov are shown in green and purple, respectively. TA_F_Ov ≫ WT_F_Ov means the genes were upregulated in TA_F_Ov. TA_F_Ov ≪ WT_F_Ov means the genes were downregulated in TA_F_Ov. TA_F_Ov ≫ WT_M_Te means genes were upregulated in TA_F_Ov. TA_F_Ov ≪ WT_M_Te means the genes were downregulated in TA_F_Ov. (G) SRY binding sites. (H) The SRY binding sites analyses of differentially expressed genes among TA_F_Ov, WT_F_Ov and WT_M_Te.
To isolate DMY-related gonad developmental-regulated genes, we conducted a comparative analysis of the three transcriptomic groups. Comparing WT_M_TE with TA_F_OV, there were 9407 differentially expressed transcripts (Data A, Dataset 1). The only difference between WT and TA medaka is the absence of the DMY gene, more or less, these differentially expressed genes were attributed to the loss of the DMY gene. There were 4629 upregulated genes and 4778 downregulated genes in WT-M-Te (Fig. 6B,C). The functions of these genes were mainly related to cilium morphogenesis (191), cilium organization (152), cilium assembly (152), spermatid differentiation and development, wnt signal pathway (77) (Dataset 1). The master male SD marker, DMY12,13 (Dmrt1Y, Uingene20290/Unigene19692), did not expressed in TA_F_OV (Dataset 1), supporting previous observations that DMY gene was successfully disrupted in XYDMY− mutants (Fig. 4C). The male sex-differentiation or testes maintaining marker Dmrt1a38 (Unigene42535/Unigene30831), Gsdf39 (Unigene44032), Sox9b40 (Unigene18419) were also significantly down-expressed in TA_F_OV (Dataset 1). Interestingly, the majority of genes in wnt signal pathway were differentially expressed between WT_M_TE and TA_F_OV. Wnt1 (Unigene54610), Wnt5a (Unigene50823), Wnt5b (Unigene25302), Wnt9 (Unigene32218), Wnt11 (CL6243), PRCK (CL13707, PREDICTED: Oryzias latipes PRKC apoptosis WT1 regulator protein-like), DKK (Unigene58048) and PRKCA (Unigene17297) were up-expressed in TA_F_OV; Wnt2 (Unigene 27916), Wnt8a (Unigene51222), FZD6 (CL9539) and FZD8 (Unigene49832) were down-expressed in TA_F_OV (Dataset 1). These genes also contribute significantly to the molecular supporting the sex-reversal female phenotype of XYDMY− mutants (Fig. 5A,B).
Theoretically, a number of differentially expressed genes between the testis (WT_M_TE) and ovary (WT_F_OV) (Data B) might confound Data A. More accurately, Data B must be excluded from Data A. The intersection of Data A and Data B identified the real differentially expressed genes between the WT XY male testis and XYDMY− female ovary (Data C, Dataset 2), which were the sum of 309 transcripts, 276 transcripts, 144 transcripts, 293 transcripts, 62 transcripts and 78 transcripts, which are DMY -related or affected genes (Fig. 6F). Relative to WT_F_OV, 144 ovary-specific transcripts were downregulated and 293 ovary-specific transcripts were upregulated in male-to-female reversed gonads (Fig. 6F). 62 transcripts in XYDMY− female ovary were downregulated relative to WT_M_TE and were upregulated relative to WT_F_OV. 78 transcripts in XYDMY− female ovary were upregulated relative to WT_M_TE and were downregulated relative to WT_F_OV. Relative to WT_F_OV and WT_M_TE, 309 transcripts were upregulated in TA_F_OV and 276 transcripts were downregulated in TA_F_OV (Fig. 6F and Dataset 2). Seventy-three testis-specific transcripts were identified (Fig. 6E and File S8). GO analysis and homologous annotation with human genes is a traditional way to further predict the functions of the genes in Data C (Dataset 2). According to the annotation of Blast2GO, genes of Data C were associated with the regulation of ubiquitination and fertilization. Moreover, a large number of genes, such as ENSORLT00000000529, have not been previously reported.
GO analysis is not a straightforward way to predict the relationships among genes. Sry and DMY are transcription factors; theoretically, their downstream genes should have the direct binding regions for Sry or DMY. The predicted binding site of human SRY is shown in Fig. 6G. Human homologous genes of data C were scanned for SRY binding sites (Dataset 3). If SRY binding sites could be found in the human homologous genes, we could speculate that their medaka homologous genes may have the DMY binding sites or be a directly affected gene. There were 9844 unique genes in medaka that were analyzed: 7440 of which had homologous genes in the human genome. There were 4644 human homologous transcripts that may have more than one potential SRY binding site (Fig. 6H and Dataset 3). A number of potential DMY regulated genes, such as SLC25A38, had a potential SRY binding site in the upstream region of its human homolog; and a novel transcript, ENSORLT00000000529, had six potential SRY binding sites in the upstream region of its human homolog. These genes are significant for investigating the transcriptional function of the DMY gene.
When we compared TA_F_OV with WT_F_OV, 1163 differentially expressed transcripts (Data D) were found in XYDMY− female (Dataset 4). There were 515 upregulated genes and 647 downregulated genes in WT-F-Ov (Fig. 6B,D). Using Blast2GO, the differentially expressed genes were blasted and annotated on biological process (BP), cell components (CC) and molecular function (MF). The genes of Data D are associated with the wnt receptor signaling pathway (Predicted: syntabbulin-like, axin-2-like), ovarian follicle development (Forkhead box O5, Predicted: beta-arrestin-1-like, adenomatous polyposis coli protein-like, ubiquitin-protein ligase E3A-like, bone morphogenetic protein receptor type-1B-like) and the follicle-stimulating hormone signaling pathway (luteinizing hormone receptor, lhr, Predicted: beta-arrestin-1-like) (Dataset 4). In terms of the number and repetitions of the genes, the genes of the wnt signaling pathway were the most significant proportion of Data D. PRCKA (CL423) and DKK (Unigene31972) were up-expressed in TA_F_OV; Wnt1 (CL10671), MAPK8 (CL4166), FZD6 (CL9539) and PRCK (CL13707) were down-expressed in TA_F_OV. These genes, especially Wnt1 and PRCK (CL13707), were down-expressed in XYDMY− mutants, compared with WT XX females (Dataset 4). Therefore, we reasonably believed that the wnt signaling pathway is one of the vital pathways on regulating the development and mature of oocytes in XYDMY− mutants, why the ovarian function of XYDMY− mutants are significant lower than that of WT females. Comprehensive Data A, B, C and D, greatly promote research into the transcriptional function of the DMY gene in medaka, not only in sex reversal, but also in the normal development and maturation of gonads.
Discussion
Nanos3UTR- TALENs effectively generated a targeted gene mutant line in Medaka
Germline transmission of the knockout genotypes is critical to obtain homozygous gene knockout animals. Generally, a 46.8% mutation rate in the F0 is not ideal for gene disruption. Perhaps this low efficiency should be attributed to the DMY gene on the Y chromosome rather than our optimized TALENs. In general, there should be equal numbers of male and female embryos in a generation. This means that the targeting efficiency of DMY-TALENs could not beyond be greater than 50%. Indeed, in our results, the efficiency was never more than 50%, even if the concentration of TALENs was increased to 800pg (Fig. 2I). To improve the germline integration efficiency, we incorporated the zebrafish nanos-3′UTR into the TALEN construct (Fig. 1A), which was reported to protect mRNA from degradation in primordial germ cells and improve the germ cell targeting efficiency28,41,42. Compared with 9.02% DMY-TALEN-induced F1 embryos carrying mutations; there were 37.56% DMY-nanos3UTR-TALEN-induced F1 embryos that carried mutations in the DMY gene (Fig. 2I and File S2). These results indicated that DMY-nanos3UTR-TALEN induced a high proportion, possible a majority, of mutant gametes in the F0 medaka. Thus, the incorporation of the nanos-3′UTR into the TALEN construct improved the germ cell targeting efficiency in medaka, permitting the generation of medaka knockout lines.
A novel program to identify potential off-target sites of TALENs
Specificity is essential to establish precisely targeted gene knockout lines. TALENs have become an accepted tool for targeted mutagenesis, but undesired off-targets, in addition to the targeted genomic region, remain an important issue15,21,43,44,45,46,47. Unfortunately, using e-PCR to perform BLAST searches, potential off-target sites were identified in several studies46. Using Primer3 and BLAST, potential off-target sites of the DMY gene in medaka were identified (File S5). According to this data, BLAST emphasized sequence similarity, but there were few base pair sites of TALENs that could match to potential off-target sites of the genomes. In fact, not only the similarity of sequence, but also that fact that the 0 position of EBEs must be T is very important to TALENs binding site48. Using the novel program, 55 candidates’ off-target sites were identified in the medaka genome. We amplified the top predicted potential off-target locus by PCR, sequenced it and found no mutations (File S4). Using our data, our program was more efficient than ePCR to predict TALENs off-target sites. Recently, tools for predicting TALEN off-targets have been developed, such as idTALE (http://idtale.kaust.edu.sa), Paired Target Finder (https://tale-nt.cac.cornell.edu) and TALENoffer49. Compared with these, the advantage of our program is its simplicity and reliability. However, a more detailed investigation on the possible off-target effects of TALENs and more accurate program will be needed in the future.
Mutant phenotypes
Gonadal dysgenesis in 46, XY patients was first noted by Drash et al.4. Phenotypic female patient with an XY karyotype were initially reported by Kaplan5. Several phenotypic female patients with the XY karyotype were evaluated clinically, cytogenetically, hormonally, endoscopically and histologically6. The functional study of gonadal dysgenesis in phenotypic female patients with an XY karyotype has been hindered by a lack of animal models with specific mutations, except for the mouse sry mutant3,31. Using TALENs, two different Y-linked genes were efficiently manipulated in mouse embryonic stem cells (mESCs)31 and an Sry knockout mouse was generated3. The mutant mice are almost completely infertile, although the Sry knockout mouse is similar to humans in terms of their physiological phenotype. The phenotype of the DMY knockout medaka is also similar to that human XY female syndrome. In particular, a majority of fertile individuals were found in the population. XYDMY− female medakas could help in studies of the mechanism of human XY female syndrome in genetics and reproductive biology. Benefiting from the number of embryos or offspring from the parents, fish are good for large sample analysis and for investigating individual differences of human XY female syndrome in populations.
DMY, a duplicated copy of DMRT1, is identified as the master male SD gene and shows all features of a SD gene in medaka12. In this study, we generated both insertions (+16) and deletions (−11) of the TALEN-induced DMY mutant line, which developed into females with the XY karyotype (Fig. 5A–C,F). Sex reversal was also reported very early in medaka50, which could be induced by steroid hormones or high temperature51,52. To evaluate whether the sex reversal observed in XYDMY− female medaka was caused by DMY gene mutation, the best method is rescue of the phenotypes of the XYDMY− female medaka. In the former case, the expected mutant DMY lacks functionally important motifs of DMY; this mutated Y chromosome could pair with the WT X and Y chromosome to generate females and males, respectively. Crossing with the WT medaka (Fig. 3), produced YDMY−Y male mutants in the next generation, which meant that dysfunction of the DMY gene was the unique difference between XYDMY− female and YDMY−Y male. This result confirmed that the sex reversal observed in XYDMY− female medakas was caused by a DMY gene mutation.
Adults of two phenotypically female mutant lines were evaluated through mutation confirmation, phenotype diagnostic (Fig. 5A) and histological analysis (Fig. 5B,F). Using genomic PCR (Fig. 4B) and FISH of DMY gene (Fig. 5C) showing the specific hybridization signal of the Y chromosome, the two phenotypically female mutant lines were identified as genetic males with the XY karyotype. RT-PCR analysis (Fig. 4C) and RNA-seq (Dataset 1) confirmed that XYDMY− mutants did not express DMY gene, which were different from WT XY individuals. Gsdf, co-localized with DMY in the same somatic cells in the XY gonads, was expressed exclusively in primordial gonads of only the genetic males39. Both the increased number of germ cells (Fig. 4D) and the significantly down-expression of Gsdf gene (Unigene44032) implied sex reversal of XYDMY− mutants took place in early developmental stages, attributed to the disruption of DMY gene. As expected, the TALEN-induced DMY knockout medakas had female external and internal specificities. Unlike the Sry KO mouse, which did not produce any offspring3, the majority of TALEN-induced DMY knockout medakas were fertile. Twelve of 15 XYDMY− mutants developed functional ovaries; and the ovaries of three females showed incomplete development. There were significant differences among XYDMY− mutants, WT female and WT male in GSI (Fig. 5D) in terms of their GSI scores. Histological analysis showed that the ovary of XYDMY− mutants displayed a reduced number of oocytes (Fig. 5F). In three of the 15 mutants, no eggs could develop beyond CA oocytes. The other 12 could generate mature eggs, however, the mature oocytes number of XYDMY− mutants are significantly fewer than that of WT XX females. Therefore, the ovary function of XYDMY− mutants was lower than that of WT females, not only in quality, but also in the quantity (Fig. 5F). To investigate whether there is significant difference in the fertilization of eggs and between XYDMY− female and WT female; test crosses were continuously and systemically recorded. During generation of offspring from the XYDMY− female testcrosses, we found that it was always difficult to obtain a sufficient number of offspring. After 10 days of continuous monitoring, 6 eggs matured per day from XYDMY− mutants; 20 eggs matured per day from WT XX females (Fig. 5E). The results from F2 phenotypes revealed that a reduced number of mature oocytes were one of the most plausible explanations for the reduced fertility in the DMY KO medakas. Thus, the XYDMY− mutants could develop functional ovaries; however, the development, maturation and fertilization capacity of their eggs were significantly lower than those of the WT female.
Transcriptomic analysis of TALENs induced DMY- mutants and the WT
Natural sex reversal has been reported in medaka50 and mutation of the DMY gene was identified in several artificial mutants14. However, the molecular mechanism of sex reversal, inducing by loss of DMY, has not been resolved. RNA-Seq is a recently developed approach for transcriptome profiling that is based on deep-sequencing32. RNA-seq could quantify the changing expression levels of each transcript during development and under different conditions. XYDMY− mutant female from the F3 generation were used for RNA-seq analysis, which minimized the off-target effects of DMY-TALENs. Using the RNA-seq approach, we obtained transcriptome information of XYDMY− mutants and through a comparative analysis of them, revealed the transcriptional function of the DMY gene in medaka.
Using RNA-seq, 157 million raw reads were generated from WT_M_TE, WT_F_OV and TA_F_OV libraries. However, only 75.8% of the processed reads were mapped to the reference genome of medaka in the Ensemble database (File S6). This indicated that the medaka genome information requires improvement and transcriptomic sequencing could revise and promote the improvement of medaka genome. In addition, it implied that it would be better to use independent analysis with no reference genome in the analysis of medaka transcriptome, which we used in this study. The problem with the medaka genomic information had little bearing on the fact that medaka is a good model for human diseases. Although only 7481 of 9844 transcripts could match the medaka genome, 7440 transcripts had homologous genes in the human genome (Fig. 6H). This suggested that medaka might be similar to humans in terms of sex determination and regulation; the medaka transcriptome data could play a role in the analysis of human sex reversal patients.
Medaka sex is primarily determined by the presence or absence of DMY gene12,13. Several studies show that Dmrt116,38, Gsdf39 and Sox9b40 is essential to maintain testis differentiation or regulate testis development. Un-expression of DMY (Uingene20290) and lower expression of Dmrt1a (Unigene42535), Gsdf (Unigene44032) and Sox9b (Unigene18419) in RNA-seq analysis provided the molecular basis to the failure of male sex determination, male-to-female sex reversal and the failure of testis differentiation or development in XYDMY− mutants (Dataset 1). Among the differentially expressed genes in TA_F_OV compared with WT_F_OV and TA_F_OV compared with WT_M_TE (Fig. 6F and Dataset 2), we found several potential factors may directly bind SRY and DMY, such as SLC25A38 and ENSORLT00000000529. This validated the transcriptional function of the DMY gene. Theoretically, the DMY gene, a SD gene, might have a relationship with the sperm production and cilia assembly. However, from our comparative analysis, it was apparent that differences in the expressions of genes involved in these processes represented background differences between the WT ovary and WT testis tissue. This is an essential problem in the transcriptional function of DMY, but it beyond the scope of this study. Further, we may investigate developmental stages of gonad or different tissue of the HPG axis to explain the problem.
The main differentially expressed genes between WT_F_OV and TA_F_OV, were genes in the wnt receptor signaling pathway (Predicted: syntabbulin-like, axin-2-like) (Dataset 4). In mammals, beta-arrestin-1-like gene regulated IGF-1 affects human reproductive endocrinology. The medaka homolog of beta-arrestin-1-like gene was detected 10 times in the RNA-seq data. The high expression of the beta-arrestin-1-like gene could provide some clues to the mechanism of the degradation of the testis and the development of the ovary in XY DMY mutant females. Lhr and Forkhead box O5 factor were also differentially expressed between TA_F_OV and WT_F_OV. Lhr is the luteinizing hormone receptor and Forkhead box O5 factor plays an important role in metabolism and cell differentiation. The low expression of lhr and Forkhead box O5 factor might affect the incomplete ovary function of the XY DMY mutant female. The majority of genes in wnt signal pathway, such as Wnt genes, PRCK genes, FZD genes and DKK were differentially expressed between WT_M_TE and TA_F_OV (Dataset 1), it implied that the wnt signaling pathway is the main regulation pathway during male-to-female sex reversal process. In addition, the genes in wnt signaling pathway, especially Wnt1 and PRCK, were up-expressed in XYDMY− mutants compared with WT XY males and down-expressed compared with WT XX females (Dataset 1 and 4). These results implied that the wnt signaling pathway also contributed primarily to the ovarian development, reduced fertility and ovarian maturation in the XYDMY− mutants. Interestingly, PRCKA and DKK were up-expressed in TA_F_OV, whether compared with WT_M_TE (Dataset 1) or compared with WT_F_OV (Dataset 4). In summary, these results implied that the wnt signaling pathway is the root of sex reversal, the incomplete ovarian function and reduced fertility in the XYDMY− mutants.
Methods
Ethics Statement on the Use of Animals
The research animals are provided with the best possible care and treatment and are under the care of a specialized technician. All procedures were approved by the Institute of Hydrobiology, Chinese Academy of Sciences and were conducted in accordance with the Guiding Principles for the Care and Use of Laboratory Animals.
Medaka husbandry
The Orange strain of medaka was used in this study (Originally from National University of Singapore). Medakas were maintained in aquaria under an artificial 14-h light/10-h dark photoperiod at 26 °C in the Institute of Hydrobiology, Chinese Academy of Science30.
Construction of TALENS
TALEN were assembled and transferred into vectors pCS2-KKR and pCS2-ELD28. To improve the germline integration efficiency, the zebrafish nanos-3′UTR was separately inserted into the 3′ end of the pCS2-ELD/KKR vector to replace the SV40 UTR using NotI and XbaI.
Manipulation of medaka embryos
The final TALEN plasmids were linearized using Not1 and the mMessage mMachine SP6 kit (Ambion) was used to synthesize mRNAs. TALENs mRNAs (half left and half right monomer mRNAs) were microinjected into medaka embryos at the one cell stage.
Detection of mutations in TALEN-targeted medaka embryos
72 hours after microinjection, TALEN targeted embryos were pooled for genomic DNA extraction (10 embryos for each pool). PCR was performed using primers DMY F and DMY R; PCR products were purified from the agarose gel using a gel extraction kit (QIAGEN). Amplicons harboring the targeted gene fragments were sub-cloned into pMD-18T using TA cloning (Takara) and single colonies were examined by PCR using primers DMY F, DMY F1 and DMY R. The PCR conditions were as follows: 5 min at 95 °C; followed by 30 cycles of 15 sec at 95 °C, 20 sec at 52 °C, 30 sec at 72 °C; and a final step of 5 min at 72 °C. PCR products were electrophoresed in a 2% agarose gel and verified by DNA sequencing.
Off-target analysis
The criteria of the novel program for determining off-target sites were that the 0 position of the EBEs must be T, from 1 to 10 mismatch bases occur in the pairs of EBEs and the spacer between the two putative EBE regions is less than 100 bp, because it has been suggested that longer spacers interfere with Fok I dimerization.
Founder Screening
TALEN-injected medaka embryos were raised to sexual maturity. F0 DMY mutated females were crossed with wild-type males; and F1 embryos were collected at 72 hours post fertilization. Genomic DNA was extracted from ten randomly pooled embryos to assess mutagenesis at the TALEN-targeted site by PCR and sequencing. F1 embryos were individually collected at 7 days post fertilization (dpf) and genomic DNA was extracted from each individual embryo to assess mutagenesis at the TALEN-targeted site by PCR and sequencing.
Fluorescence in situ Hybridization
FISH was performed using a DMY fragment labelled by PCR (Dmy Probe F: 5′-TGCCCAAGTGCTCCC-3′; Dmy Probe R: 5′-CCCCTTTTGTCTGTCCTCT-3′) with digoxigenin-11-dUTP (Boehringer). Standard nick translation using biotin-16-dUTP and digoxigenin-11-dUTP12 separately labeled the probe. Before hybridizing with denatured medaka mitotic chromosomes, the probe was denatured and preannealed in the presence of excess genomic DNA. Rhodamine-conjugated avidin and antidigoxigenin (monoclonal)-conjugated fluorescein (Sigma) were used to detect the hybridization sites for the probe simultaneously. 4, 6-diamidino-2-phenylindole (DAPI) was used to counterstain the chromosomes. EASY FISH 1.0 software (Applied Spectral Imaging, Mannheim, Germany) was used to capture and display digitized images of the rhodamine signals on DAPI-stained chromosomes.
Expression Analysis
The TRIZOL reagent (Invitrogen) was used to extract total RNA from pooled organs of three adult medaka fish, according to the supplier’s recommendation. After DNase treatment, reverse transcription was performed with 2 or 4 mg RNA by using Superscript II reverse transcriptase (Invitrogen) and random primers. cDNA from 10 ng (actin) to 200 ng (adult organs) of total RNA was used for PCR with gene-specific primers: Ola Actin, Ola DMY (Dmy F1 and PG17.89 primers12).
Histological Analysis
For the histological analysis, identified mutants from the F2 generation of medaka were dissected into head and body segments (N = 15). Each dissected head was used to determine the genetic sex. The body portions were fixed overnight in Bouin’s fixative solution and then embedded in paraffin. Cross-sections were cut serially at 5 μm thickness and after hematoxylin and eosin (H&E) staining; the images were obtained under a microscope (Eppendorf).
Library construction and high-throughput sequencing
Medaka gonads (divided into WT-F, WT-M and TA-F, 10 samples pooled in each group, respectively) were collected for RNA extraction. TRIZOL (Invitrogen) performed the total RNA extraction, following the manufacturer’s instructions. RNA library construction was then performed by BGI- Shenzhen, Shenzhen, China then constructed the RNA library. Before library construction, the integrity of RNA samples was confirmed using an Agilent 2100 Bioanalyzer; 10 μg of total RNA was used for isolation of mRNA with Sera-mag Magnetic Oligo (dT) beads from Illumina.
Quantitative Real-Time PCR
The TRIZOL reagent (Invitrogen) was used to isolate total RNA from embryos. The SYBR Green PCR Master Mix (Applied Biosystems) and the Real-Time PCR System (ABI 7900) measured gene transcription. The actb1 gene was used as the endogenous control gene for normalizing expression of the target gene. Triplicate technical replicates were performed for duplicate cultures.
Functional Annotation, Classification and Enrichment Analysis
Blast2GO software (http://www.blast2go.de) was used to perform BLAST searching, mapping and annotation of proteins differentially expressed53. BINGO 2.4454 plug-in in the Cytoscape platform55 was used to perform functional enrichment analysis of differentially expressed proteins to determine the significantly enriched GO terms and relevant proteins. Enrichment analysis of GO term assignment was performed in reference to the entire annotated P. tricornutum proteome. The corrected P values were derived from a hypergeometric test followed by Benjamini and Hochberg false discovery rate correction. A corrected P < 0.05 was regarded as significant.
Additional Information
Accession codes: All RNA-seq data submited to GEO, GEO accession: GSE62700.
How to cite this article: Luo, D. et al. Direct production of XYDMY- sex reversal female medaka (Oryzias latipes) by embryo microinjection of TALENs. Sci. Rep. 5, 14057; doi: 10.1038/srep14057 (2015).
References
Sinclair, A. H. et al. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 346, 240–244 (1990).
Hawkins, J. R. et al. Mutational analysis of SRY: nonsense and missense mutations in XY sex reversal. Hum Genet 88, 471–474 (1992).
Kato, T. et al. Production of Sry knockout mouse using TALEN via oocyte injection. Sci Rep 3, 3136 (2013).
Drash, A., Sherman, F., Hartmann, W. H. & Blizzard, R. M. A syndrome of pseudohermaphroditism, Wilms’ tumor, hypertension and degenerative renal disease. J Pediatr 76, 585–593 (1970).
Kaplan, E. Gonadal dysgenesis in a phenotypic female with an XY chromosomal constitution. S Afr Med J 53, 552–553 (1978).
Portuondo, J. A. et al. Management of phenotypic female patients with an XY karyotype. J Reprod Med 31, 611–615 (1986).
Conover, D. O. & Kynard, B. E. Environmental sex determination: interaction of temperature and genotype in a fish. Science 213, 577–579 (1981).
Bulmer, M. Evolution: sex determination in fish. Nature 326, 440–441 (1987).
Conover, D. O. & Heins, S. W. Adaptive variation in environmental and genetic sex determination in a fish. Nature 326, 496–498 (1987).
Loukovitis, D. et al. Quantitative trait loci for body growth and sex determination in the hermaphrodite teleost fish Sparus aurata L. Anim Genet 43, 753–759 (2012).
Piferrer, F., Ribas, L. & Diaz, N. Genomic approaches to study genetic and environmental influences on fish sex determination and differentiation. Mar Biotechnol (NY) 14, 591–604 (2012).
Nanda, I. et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc Natl Acad Sci USA 99, 11778–11783 (2002).
Matsuda, M. et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 417, 559–563 (2002).
Matsuda, M. et al. DMY gene induces male development in genetically female (XX) medaka fish. Proc Natl Acad Sci USA 104, 3865–3870 (2007).
Otake, H. et al. The medaka sex-determining gene DMY acquired a novel temporal expression pattern after duplication of DMRT1. Genesis 46, 719–723 (2008).
Masuyama, H. et al. Dmrt1 mutation causes a male-to-female sex reversal after the sex determination by Dmy in the medaka. Chromosome Res 20, 163–176 (2012).
Esvelt, K. M. & Wang, H. H. Genome-scale engineering for systems and synthetic biology. Mol Syst Biol 9, 641 (2013).
Bibikova, M., Golic, M., Golic, K. G. & Carroll, D. Targeted chromosomal cleavage and mutagenesis in Drosophila using zinc-finger nucleases. Genetics 161, 1169–1175 (2002).
Kim, Y. G., Cha, J. & Chandrasegaran, S. Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci USA 93, 1156–1160 (1996).
Kim, S. et al. Preassembled zinc-finger arrays for rapid construction of ZFNs. Nat Methods 8, 7 (2011).
Hockemeyer, D. et al. Genetic engineering of human pluripotent cells using TALE nucleases. Nat Biotechnol 29, 731–734 (2011).
Huang, P. et al. Heritable gene targeting in zebrafish using customized TALENs. Nat Biotechnol 29, 699–700 (2011).
Li, T. et al. TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res 39, 359–372 (2011).
Christian, M. et al. Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 186, 757–761 (2010).
Ishino, S. et al. Nucleotide sequence of the meso-diaminopimelate D-dehydrogenase gene from Corynebacterium glutamicum. Nucleic Acids Res 15, 3917 (1987).
Hwang, W. Y. et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotechnol 31, 227–229 (2013).
Cong, L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823 (2013).
Liu, Y. et al. Inheritable and Precise Large Genomic Deletions of Non-Coding RNA Genes in Zebrafish Using TALENs. PLoS One 8, e76387 (2013).
Liu, Y. et al. A highly effective TALEN-mediated approach for targeted gene disruption in Xenopus tropicalis and zebrafish. Methods 69, 58–66 (2014).
Qiu, C. et al. Efficient knockout of transplanted green fluorescent protein gene in medaka using TALENs. Mar Biotechnol (NY) 16, 674–683 (2014).
Wang, H. et al. TALEN-mediated editing of the mouse Y chromosome. Nat Biotechnol 31, 530–532 (2013).
Wang, Z., Gerstein, M. & Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10, 57–63 (2009).
Ansai, S. et al. Efficient targeted mutagenesis in medaka using custom-designed transcription activator-like effector nucleases. Genetics 193, 739–749 (2013).
Miller, J. C. et al. A TALE nuclease architecture for efficient genome editing. Nat Biotechnol 29, 143–148 (2011).
Kobayashi, T. et al. Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the medaka, Oryzias latipes. Dev Dyn 231, 518–526 (2004).
Otake, H. et al. Wild-derived XY sex-reversal mutants in the Medaka, Oryzias latipes. Genetics 173, 2083–2090 (2006).
Kagawa, H., Young, G. & Nagahama, Y. Relationship between seasonal plasma estradiol-17 beta and testosterone levels and in vitro production by ovarian follicles of amago salmon (Oncorhynchus rhodurus). Biol Reprod 29, 301–309 (1983).
Hornung, U., Herpin, A. & Schartl, M. Expression of the male determining gene dmrt1bY and its autosomal coorthologue dmrt1a in medaka. Sex Dev 1, 197–206 (2007).
Shibata, Y. et al. Expression of gonadal soma derived factor (GSDF) is spatially and temporally correlated with early testicular differentiation in medaka. Gene Expr Patterns 10, 283–289 (2010).
Nakamura, S. et al. Sox9b/sox9a2-EGFP transgenic medaka reveals the morphological reorganization of the gonads and a common precursor of both the female and male supporting cells. Mol Reprod Dev 75, 472–476 (2008).
Koprunner, M., Thisse, C., Thisse, B. & Raz, E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev 15, 2877–2885 (2001).
Mishima, Y. et al. Differential regulation of germline mRNAs in soma and germ cells by zebrafish miR-430. Curr Biol 16, 2135–2142 (2006).
Mussolino, C. & Cathomen, T. On target? Tracing zinc-finger-nuclease specificity. Nat Methods 8, 725–726 (2011).
Mussolino, C. et al. A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity. Nucleic Acids Res 39, 9283–9293 (2011).
Tesson, L. et al. Knockout rats generated by embryo microinjection of TALENs. Nat Biotechnol 29, 695–696 (2011).
Lei, Y. et al. Efficient targeted gene disruption in Xenopus embryos using engineered transcription activator-like effector nucleases (TALENs). Proc Natl Acad Sci USA 109, 17484–17489 (2012).
Osborn, M. J. et al. TALEN-based gene correction for epidermolysis bullosa. Mol Ther 21, 1151–1159 (2013).
Boch, J. et al. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509–1512 (2009).
Grau, J., Boch, J. & Posch, S. TALENoffer: genome-wide TALEN off-target prediction. Bioinformatics 29, 2931–2932 (2013).
Yamamoto, T. Artificial induction of functional sex-reversal in genotypic females of the medaka (Oryzias latipes). J Exp Zool 137, 227–263 (1958).
Paul-Prasanth, B., Shibata, Y., Horiguchi, R. & Nagahama, Y. Exposure to diethylstilbestrol during embryonic and larval stages of medaka fish (Oryzias latipes) leads to sex reversal in genetic males and reduced gonad weight in genetic females. Endocrinology 152, 707–717 (2011).
Hattori, R. S. et al. Temperature-dependent sex determination in Hd-rR medaka Oryzias latipes: gender sensitivity, thermal threshold, critical period and DMRT1 expression profile. Sex Dev 1, 138–146 (2007).
Conesa, A. et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676 (2005).
Maere, S., Heymans, K. & Kuiper, M. BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21, 3448–3449 (2005).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13, 2498–2504 (2003).
Acknowledgements
We thank Ms. Li Ming (Institute of Hydrobiology, Chinese Academy of Sciences) for her kind help during the feeding of medaka and microinjection of medaka embryos. This work was supported by the National Science Foundation for Distinguished Young Scholars of China (31325026 to WH), the National High Technology Research and Development Program of China (2011AA100404 to WH), the National Science Foundation of China (31472263 to DL) and the Chinese Academy of Sciences (XDA08010106 and 2011FBZ19 to WH).
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: D.L. and W.H. Performed the experiments: D.L., Y.L., J.C., M.C., B.C., X.W., C.Q. and Y.Z. Analyzed the data: D.L., X.X., W.G., C.H.K.C., Z.Z. and W.H. Wrote the paper: D.L., C.H.K.C., Z.Z. and W.H. Contributed reagents/materials/analysis tools: D.L. and W.H.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Luo, D., Liu, Y., Chen, J. et al. Direct production of XYDMY− sex reversal female medaka (Oryzias latipes) by embryo microinjection of TALENs. Sci Rep 5, 14057 (2015). https://doi.org/10.1038/srep14057
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep14057
This article is cited by
-
Functional analysis of the promoter of the dmrt1 gene in Chinese tongue sole, Cynoglossus semilaevis
Journal of Oceanology and Limnology (2019)
-
Autosomal gsdf acts as a male sex initiator in the fish medaka
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
