
Abstract
True predators are characterised by capturing a number of prey items during their lifetime and by being generalists. Some true predators are facultative specialists, but very few species are stenophagous specialists that catch only a few closely related prey types. A monophagous true predator that would exploit a single prey species has not been discovered yet. Representatives of the spider family Ammoxenidae have been reported to have evolved to only catch termites. Here we tested the hypothesis that Ammoxenus amphalodes is a monophagous termite-eater capturing only Hodotermes mossambicus. We studied the trophic niche of A. amphalodes by means of molecular analysis of the gut contents using Next Generation Sequencing. We investigated their willingness to accept alternative prey and observed their specific predatory behaviour and prey capture efficiency. We found all of the 1.4 million sequences were H. mossambicus. In the laboratory A. amphalodes did not accept any other prey, including other termite species. The spiders attacked the lateral side of the thorax of termites and immobilised them within 1 min. The paralysis efficiency was independent of predator:prey size ratio. The results strongly indicate that A. amphalodes is a monophagous prey specialist, specifically adapted to feed on H. mossambicus.
Similar content being viewed by others


Introduction
Predators can be classified into herbivores, parasites, parasitoids and true predators1. These categories differ not only in the type of food consumed but also in the width of their trophic niche. While some herbivores, many parasites and parasitoids frequently feed only on a few host species and are often specialists, true predators typically capture a wide variety of prey. As true predators capture a number of prey items during their lifetime they are usually generalists2. Some species of true predators, such as snakes3 or spiders4, only capture a few prey types. These are, however, examples of facultative stenophagy or local specialisation, with exploitation of a locally abundant prey5. Such predators lack specialised capture adaptations6.
Stenophagous specialised true predators (sensu6) that capture only certain prey and possess specialised adaptations are rather rare. The trophic width of these predators is restricted yet it includes a few representatives of a family or a few species of one genus. For example, acarophagous mites catch several other mite species7, araneophagous spiders catch spiders from several families8, aphidophagous coccinellids catch aphids of a single genus9, lepidopterophagous bolas spiders capture male moths of a few species belonging to distinct families10 and myrmecophagous spiders feed on several species of a single subfamily11. A theoretical monophagous true predator, one in which all individuals are adapted to exclusively exploit a single prey species throughout their life and ignore even prey closely related to the focal species, has not been discovered yet.
Spiders are the largest (most diversified) order of terrestrial true predators12. Most species are generalist predators, some are oligophagous, while a few are stenophagous13. Stenophagous specialised spiders have been found to catch other spiders, woodlice, dipterans, lepidopterans, ants and termites6. Although termites are locally abundant and highly palatable, they are only exploited by a few termite-eating spiders: Salticidae14, Theridiidae15, Zodariidae16 and Ammoxenidae17. This is probably because the majority of termites live underground or in wood and have highly unpredictable and brief activity on the surface, restricting their accessibility to spiders18.
All representatives of the family Ammoxenidae appear to be adapted to catch termites19. Some Ammoxenidae have been reported to feed only on the harvester termite, Hodotermes mossambicus (Hagen) (Isoptera: Hodotermitidae), which is the only species of the genus occurring in southern and eastern Africa20. Hodotermes mossambicus inhabits subterranean nests and forage on grass upon the surface during short activity periods21,22. Anecdotal observations suggest that some Ammoxenus species may be specialists of this termite species17,21,22,23,24,25.
Rigorous analysis of the trophic niche of a predator can be based on direct observations of prey consumption or on the morphological analysis of prey remnants. The latter can be successfully used only in sedentary species, such as web-building spiders, that store remnants in their web26. In cursorial species it is difficult to investigate the trophic niche using any of these methods, as they feed cryptically (e.g. during the night or under vegetation) and do not store prey remnants. Molecular approaches provide useful tools for determining the dietary breadth of such predators. Prey DNA sequences in the gut content of predators have been successfully analysed in many studies27,28,29,30. Next generation sequencing (NGS), in particular, allows reliable, rapid and simultaneous amplification and subsequent identification, of thousands of prey DNA sequences from the guts of many individual predators separately31,32. It is necessary to target short prey amplicons (to ensure they survived digestion in the predator27,33), amplify part of the barcoding region of the cytochrome c oxidase I gene (to maximise the chances of identification from databases such as GenBank and BOLD, the Barcoding of Life Database), and, where possible, design primers that will amplify the DNA of a range of prey but not that of the predator28. Such primers must amplify sequences that are variable enough to distinguish taxa to the highest level possible34,35.
Here we focused on Ammoxenus amphalodes Dippenaar & Meyer, in which preliminary observations suggest that it may only capture H. mossambicus harvester termites21. We tested the hypothesis that A. amphalodes are monophagous, feeding on a single termite species. Ammoxenus amphalodes is a common, widespread species that is endemic to South Africa. It mainly inhabits open plains in grassland and savannah habitats and usually lives in sandy soils near to H. mossambicus nests. We investigated the natural prey of these spiders by means of NGS. Then we investigated their willingness to accept alternative sympatrically-occurring prey types under standardised laboratory conditions. Finally, we observed their predatory behaviour and prey capture efficiency to reveal their behavioural and venomic adaptations.
Results
Trophic niche
The primers (AMF1 and AMR1) successfully amplified DNA from all potential prey taxa. The new primers were specific enough to amplify DNA from all the potential prey, but not that of A. amphalodes. Altogether, prey DNA fragments were successfully amplified from 46 females, 34 males and seven juveniles. In four females and three males (7.5%) no fragments were amplified. Ion Torrent sequencing gave us, after filtering, 1,755,965 sequences (70% of all reads). 1,371,138 sequences were used in further analyses after removing rare haplotypes and sequences with indels changing the reading frame. The number of sequences per spider varied from 5 to 101,242. Generally, fewer sequences were obtained from samples that were extracted from agarose gels (due to presence of very strong dimers during PCRs) than from samples that were purified without cutting from the gels. MOTU analysis showed 16 variants, ranging from 1 to 4 “MOTUs” (groups of haplotypes differing by 4 bp) per spider. Such variation was probably caused by sequencing errors; all but one “MOTUs” contained only 2–4 sequences.
The great majority of sequences were assigned to H. mossambicus (99.8% of valid sequences) when compared to GenBank and BOLD databases. Almost all those sequences were similar to H. mossambicus reference sequences above 99%. Odontotermes species differed from H. mossambicus only by 4 bases in the amplified fragment. When searching for the specific mutations, no base combinations corresponding to the Odontotermes sp. were found. Only 32 valid sequences were less than 99% similar to H. mossambicus (according to BOLD database criterion) but were not assigned to any other species. Two identical sequences were not assigned to any arthropod taxon with a match higher than 95%.
There was a significant difference in the number of sequences (from DNA not cut from gels, N = 46) obtained from females, males and juveniles (GLM-p: F2,43 = 32204, P < 0.0001) (Fig. 1). The highest number of termite sequences was found in juveniles.

Comparison of the mean numbers of Hodotermes mossambicus termite sequences found in the guts of females (N = 17), males (N = 23) and juveniles (N = 7) of Ammoxenus amphalodes spiders.
Seven arthropod orders were found as potential prey of A. amphalodes spiders (Table 1) in the two field plots. Hodotermes mossambicus was not a frequent prey item on the soil surface, where A. amphalodes foraged. The majority of potential prey were ants (91%, N = 701). Comparison of the potential and actual prey showed a significant difference (χ29 = 459.6, P < 0.0001). Smith’s index of trophic niche is 0.07 indicating very high level of stenophagy.
Prey acceptance
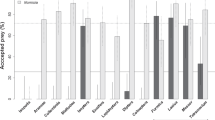
All 32 female spiders accepted H. mossambicus as prey but none even attempted to catch other termite species or any other prey. They did not even emerge from the soil to make contact with the alternative prey offered to them.
Predatory behaviour
The hunting sequence was composed of the following events: emerge (the spider emerged from the sand), chase (chasing the moving termite), attack (the spider aimed to attack the termite), bite (the spider bit the termite and held it) and dig in (the spider dragged the immobilized termite and dug with it into the soil) (see Supporting information). The flow diagram (Fig. 2) shows that 72% (N = 32) of spiders needed more than one attempt to catch the termite, repeatedly digging themselves in and out of the soil and making sequential attacks. The remaining individuals (28%) successfully captured the termite at the first attempt. A successful bite was inflicted behind the termite’s head on the lateral side of the thorax. On average (1.96 ± 0.22 (SE)), spiders captured the termite on the second attempt. The termite struggled hard after being bitten, but the spider held it firmly while lying on its back. Then the spider dug itself into the soil together with the immobilized termite and fed on the termite while hidden in the soil.

Flow diagram of hunting sequence of Ammoxenus amphalodes spiders (N = 32) when hunting Hodotermes mossambicus termites.
Percentages signify the proportion of individuals transferring from previous step to the next one. Pictures show particular behaviours (attack, holding and carrying, burying beneath prey). Photographs made by Stano Pekár.
It took a mean of 82 s (SE = 10.5) to immobilize a termite completely. The paralysis latency did not differ between male and female spiders (N = 40; GLM-g: χ21 = 0.05, P = 0.83) and there was no relationship between termite/spider size ratio and the paralysis latency (N = 40; GLM-g: χ21 = 0.05, P = 0.84, Fig. 3).

Relationship between paralysis latency after final bite and termite/spider body size ratio in female and male Ammoxenus spiders (N = 40).
The GLM revealed no relationship (GLM-g: χ21 = 0.05, P = 0.84).
Discussion
This paper appears to be the first where invertebrate predators have been individually screened using NGS. It is also one of very few papers to use NGS to analyze the diet of an invertebrate species36. The combination of prey DNA analysis with behavioural experiments allowed us to obtain more reliable data on a species’ diet than any single approach. The results of the DNA analyses were in close concordance with data from the behavioural experiments.
Unlike all other papers29,31,32,35,36,37,38 using NGS to analyse diet to date only one prey species was detected and identified (99.8% of valid sequences). Although different “MOTUs” were isolated, all had their closest match to one termite species, H. mossambicus. Although some MOTU could have belonged to different termite haplotypes, or possibly nuclear copies of the mtDNA sequences within the nuclear genome were present, we think it more likely that most of the differences were caused by sequencing errors. Evidence for this comes from the very low sequence numbers (<4) of different MOTUs found within individual spider predators. Several factors have quantitative affects on DNA amplification during gut content analysis, particularly primer efficiency, DNA copies number per cell in different prey species, the effect of temperature and predator activity on DNA degradation and differences in the digestibility of different prey29,39 (e.g. soft-bodies vs. chitinous invertebrate prey). Given that (1) we used the same primers throughout, (2) there was only one prey species and (3) all spiders and termites were living under the same microclimatic conditions (temperature), we can make an approximate comparison of the relative quantities of ingested prey. The highest number of sequences per individual was found in juvenile spiders. This is likely because juveniles may catch prey more often than adults due to their investment in ontogenetic development40,41.
Our results confirm that A. amphalodes is a monophagous termite-eater. However, we expected that it would catch and exploit several termite species. Although sequences for H. mossambicus and Odontotermes sp. collected at the same site were so similar, when compared to the databases no sequence was assigned to any Odontotermes sp. (among 34 sequences of the barcoding region available in the BOLD database), not even with a lower percentage similarity. Therefore, the likelihood that the sequences were incorrectly assigned to H. mossambicus is very low. The two termite species differ considerably in their ecology: H. mossambicus forages upon the soil surface and is thus available to A. amphalodes, whereas Odontotermes sp. inhabits wood and is thus inaccessible. Furthermore, in the laboratory A. amphalodes spiders did not accept other termite species. There are two other harvester termite species occurring in South Africa, Trinervitermes trinervoides and Microhodotermes viator (Latreille 1804), which have been reported to be prey of other Ammoxenus species19,21,24. However, we have not found any sequence that would correspond to either of those two species. Microhodotermes viator does not occur in the Free State Province where this study was conducted20. Thus, all our evidence shows that the population of A. amphalodes at our study site is a monophagous termite-eating true predator.
If A. amphalodes was a facultative stenophage then it would accept alternative prey when available, under laboratory conditions. Yet, our results show that not only were different prey types refused, but other sympatrically-occurring termite species were rejected. The experiments were conducted with experienced adult spiders, which might have developed a preference for H. mossambicus during their ontogenetic development. A previous study showed that even freshly hatched spiderlings capture H. mossambicus25, supporting suggestions for an obligate preference.
Previous continuous pitfall-trap sampling over two years revealed that both A. amphalodes and H. mossambicus are active throughout the year. However, A. amphalodes has two seasonal peaks—it is a bivoltine species with one reproductive period in September and the other in March. This coincides with seasonal activity of H. mossambicus termites but not with other sympatrically occurring termite species, namely T. trinervoides (Haddad, unpublished). In some places A. amphalodes was even observed to hide in the mounds of the latter termite species but results of our study, both field and laboratory, show that A. amphalodes does not feed on it. Additionally, this study found that there is a synchronised spatial co-occurrence of A. amphalodes and H. mossambicus.
In a monophagous predator all populations and all developmental stages would have to catch a single prey species. Our data were obtained from a single A. amphalodes population occurring in the central South Africa. Previous work on the trophic ecology of A. amphalodes conducted in the north-eastern part of South Africa provided evidence that this species fed exclusively on H. mossambicus there as well17,21,25. This supports our view that not only is the population we studied monophagous, but that the species as a whole is a monophagous termite-eater. Furthermore, the distributional overlap of A. amphalodes and H. mossambicus supports this view20,42.
Specialist predators must be adapted to deal with extreme predator-prey size differences, with tiny juveniles capable of capturing prey as large as those attacked by adults. Prey specialists seem to be adapted to catch extremely large prey. For example, ant-eating Zodarion cyrenaicum Denis that occurs in the Negev Desert captured mainly large Messor arenarius Fabricius ants43. Surprisingly, even the first instar juveniles captured this ant species, even though the ants are gigantic when compared to the body size of the spiders44. All ontogenetic stages of A. amphalodes were found to catch H. mossambicus21,25. The body size of workers of this termite species is sexually dimorphic45, so it might be possible that tiny spiderlings select the smaller female morphs while adult spiders select the larger male morphs. A similar trend has been observed in ant-eating Zodarion spiders44.
Although several ant-eating spiders of the genus Zodarion appear to be monophagous, as they only captured one prey species in the wild44, in the laboratory they accepted other ant species, though the capture was less efficient. In A. amphalodes, other termites did not even elicit an attack. The difference stems from the fact that prey selection in specialists can take place at various levels during the predation sequence: encounter—detection—recognition—immobilisation—capture—processing46,47. At each step of the predation sequence, specific adaptations may function as filters that exclude some of the available prey types, so that fewer prey types are left at the next step. A high level of specificity will correspond to the filtering at earlier stages in the predatory sequence. Specifically, in A. amphalodes alternative prey did not even elicit the hunting behaviour, suggesting that the filter is at the level prior to direct encounter, possibly through the detection of vibrations of foraging termites in the substrate24. This again points to monophagy of A. amphalodes.
It is not known which cues produced by H. mossambicus are used to elicit the predatory behaviour in A. amphalodes. The cues must be very specific and different from those produced by sympatrically occurring alternative prey. Alarm, trail or sex pheromones produced by prey have been reported for prey-specialists48,49, but these are used at a greater distance. During inactivity, Ammoxenus spiders hide singly in a silk cell buried in soil mounds around the entrance to the termite nest17. As Hodotermes termites begin their foraging activity their movement upon the soil surface may produce vibratory cues that subsequently elicit the predatory behaviour of A. amphalodes24.
Ammoxenus amphalodes seem to possess very effective venom to paralyse large individuals of H. mossambicus. The prey was paralysed within approx. 1 min and the paralysis efficiency was independent of predator-prey body size ratio, even for extreme ratios, when the termite was ten times larger than the spider prosoma. A paralysis latency of 30–35 s was reported for adult A. pentheri feeding on H. mossambicus25, while 2nd and 3rd instars took approximately 1 min to paralyze prey. In a study on paralysis efficiency of myrmecophagous spider specialists we found that the efficiency decreases with an increasing body size ratio44. This is likely because larger prey requires a larger volume of venom to take its effect. Absence of a relationship here suggests that either A. amphalodes spiders adjusted the amount of injected venom perfectly according to the prey size, or they possess very effective venom compounds. A similarly high venom efficacy has been found in ant-eating Zodarion spiders44.
Ammoxenus amphalodes seems to consume one termite per week, which amounts to dozens of individuals during their life cycle. The termites have, however, unpredictable foraging activity, often staying inactive for several days at a time. How does the spider deal with their absence? The silken retreats of A. amphalodes containing up to four immobile H. mossambicus termites were observed22. Even more surprisingly, they were not dead, but only paralyzed. The authors suggested that they could serve as food storage and be used during periods when termites are inactive.
We conclude that both the DNA analyses and laboratory experiments support the hypothesis that A. amphalodes is a specialized predator of a single termite species, H. mossambicus. The evidence suggest that this is the first case of a monophagous true predator, although we cannot entirely exclude the possibility that in other sites in Africa, where different sympatric termite species are found, they may take other species too. As in specialised herbivores, parasites and parasitoids, monophagy in true predators seems to evolve when the prey is considerably larger than the predator. Other ammoxenid species may show similar trophic ecology. Indeed, there is anecdotal evidence that A. pentheri Simon is a specialised predator of H. mossambicus23, whereas Rastellus sabulosus Platnick & Griffin feeds only on Psammotermes allocerus Silvestri (Isoptera: Rhinotermitidae)50.
Methods
Potential prey
Fieldwork was conducted in a grassland at the Amanzi Private Game Reserve near Brandfort (S 28° 35’ 53″ E 26° 25’ 04″), South Africa in March 2013. Ammoxenus spiders are rare free-living soil-dwellers hiding in the termite mounds25, thus are difficult to catch. Observations from previous years revealed that in March there is a seasonal peak of adult occurrence (and reproduction) of A. amphalodes (Haddad, unpublished). At this time females were expected to be maximising prey capture, therefore the chances to observe prey capture should be high. Ammoxenus amphalodes were found in a strip of short grass and bare soil, about 20 m wide and 200 m long, between two fields of cultivated Pangola grass (Digitaria eriantha). Previous observations revealed that A. amphalodes are active during the day (Haddad, pers. observ.). To investigate the potential prey of A. amphalodes, two square plots, each 10 × 10 m, were marked in the morning when the spiders were active. These plots were at least 20 m from each other. Arthropods occurring in the plots with body size between 3–15 mm were counted and recorded by visual census of the entire area which lasted for a period of 2 hours. The survey provided not complete but basic idea of the potential prey. Most arthropods were determined to order level, but ants were determined to genus level. Three representatives of each arthropod taxon found were stored in 100% ethanol. These included Hippodamia sp. (Coleoptera: Coccinellidae), Zonocerus sp. (Orthoptera: Acrididae), Hodotermes mossambicus, Odontotermes sp. (Isoptera: Termitidae: Termitinae), Trinervitermes trinervoides (Sjöstedt) (Isoptera: Termitidae: Nasutitermitinae), Ligariella sp. (Mantodea: Mantidae), Pheidole sp. (Hymenoptera: Formicidae: Myrmicinae), Lygaeidae (Hemiptera: Heteroptera), Nysius sp. (Hemiptera: Orsilidae) and Thanatus vulgaris Simon (Araneae: Philodromidae). DNA from these specimens was later used as non-target species for sequence alignment during primer design and for primer testing (see below).
Actual prey
To investigate the actual prey of A. amphalodes, spiders were hand-collected with tubes in the strip of short grass over the course of a few days, at different times, at Amanzi Private Game Reserve. A total of 130 spiders were collected and immediately placed in ethanol. Six A. amphalodes individuals were starved to death which occurred after at least one month. The DNA of these individuals and DNA from potential prey was extracted using the salt precipitation method.
As the DNA in the gut of spiders is progressively degrading due to digestion51, it was necessary to use primers that amplified fragments <300 bp27,52 which are still variable enough to distinguish between taxa34,35. In the starved spiders and the potential prey, the barcoding region of the COI gene was first amplified using the LCO and HCO primers53 and Go Taq G2 Flexi DNA Polymerase (Promega) under the following conditions: initial denaturation at 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 48 °C for 30 s as an annealing temperature, 72 °C for 1 min; and a final extension at 72 °C for 7 min. The reaction mixture total volume of 20 μL consisted of 8.3 μL nuclease-free water, 4 μL of 5x Green GoTaq buffer, 2.5 μL of 25 mM MgCl2, 1 μL of 10 mM dNTP’s, 1 μL of 10 μM forward and 1 μL of reverse primer, 0.2 μL of GoTaq G2 Flexi DNA polymerase (5 u/μL) and 2 μL of DNA. PCR products were detected by electrophoresis in 2% SafeView-stained agarose gels. Amplified products were sequenced on an ABI Prism 3130 Genetic Analyzer (Applied Biosystems). Sequences were aligned using Mega 5.154. Additional sequences downloaded from the GenBank database, representing another African termite and spiders (accession numbers NC_018122.1, JF302834.1, JF302835.1, JF302833.1, JF302832.1, JF302836.1, JF923296.1, JX023555.1), were included in the alignment. Two primer pairs, which would amplify DNA of potential prey, but not the spiders, were designed using Amplicon.b0855. The primers were tested to determine PCR conditions for successful amplification of all the potential prey. The primer pair AMF1: 5′- AGCAGGAATAGTAGGAACAT-3′ and AMR1: 5′-CCWCTTTCWACTATTCTTC-3′, which amplified a 250 bp fragment, was chosen for subsequent analyses and modified with MID identifiers (10 bp tags) and Ion Torrent adaptors. We used 10 MIDs added to the forward primers and 10 different MIDs added to the reverse primers to assign prey sequences to individual predators. This gave us the capacity to separate sequences derived from the gut contents of up to 100 individual spiders31.
DNA was extracted from 94 spider abdomens using the DNeasy Blood & Tissue Kit (Qiagen) following the manufacturer’s protocol (purification of DNA from animal tissues) with a change in a final elution step (only 80 μL of AE buffer were used). In total, seven juveniles and 87 adults (37 males and 50 females) were screened. PCR reactions were performed using the Multiplex PCR kit (Qiagen) under the following conditions: initial denaturation at 95 °C for 15 min; 43 cycles of 94 °C for 30 s, 47.2 °C for 90 s (annealing temperature), 72 °C for 90 s; and a final extension at 72 °C for 10 minutes. Reaction mixture total volume of 20 μL consisted of 10.6 μL of Multiplex PCR Master Mix, 1.8 μL of Q-Solution, 3 μL of RNase-free water, 0.8 μL of 10 μM forward and 0.8 μL of reverse primers and 3 μL of DNA. PCR products were detected by electrophoresis in 2% SafeView-stained agarose gels. Samples which did not form visible dimers (or only very weakly) were purified using QIAquick PCR Purification Kit (Qiagen). In cases where dimers were strong, DNA was cut from the agarose gel and extracted using the QIAquick Gel Extraction Kit (Qiagen). Both extractions and purifications were performed according to the manufacturer’s protocols. Concentration of all PCR products was assessed by comparison with the 100 bp ladder (BioLabs). Then, 5 μL of 50 μg/μL PCR products was pooled into the same sterile vial and sent for sequencing. Enrichment (emPCR) and one-directional sequencing on an Ion Torrent PGM with a 316 chip was performed at the Centre de Recerca en Agrigenòmica (Bellaterra - Barcelona, Spain).
The sequences were processed using the Galaxy platform (https://usegalaxy.org/)56 and BioEdit 7.2.557. Reads were split according to their MIDs (with two mismatches and two deletion thresholds allowed), resulting in files corresponding to individual spiders. Sequences were filtered according to their length (<200 bp) to remove dimers or too short reads. The sequences were collapsed and rare haplotypes (containing <2 identical sequences) were removed as well as sequences with stop codons and indels changing the reading frame to eliminate sequencing errors. The remaining haplotypes were clustered into MOTUs (= molecular operational taxonomic units) using jMOTU 4.158 with a 4-bp cut-off (corresponding to 1.6% sequence divergence), following31,32,37. The variation among species is greater than 2% (or even more than 4% in some taxa) when using COI34. We decided to use a lower threshold because H. mossambicus and Odontotermes sp. differed by only 4 bp in the targeted COI region. Each MOTU was compared to the GenBank database (http://blast.ncbi.nlm.nih.gov/Blast.cgi) using megablast, the BOLD database (http://www.boldsystems.org/) and also to sequences obtained from the potential prey specimens. In BOLD, sequences are assigned to species level when their similarity is higher than 99%. Some sequences (0.2%) did not match with any known sequence. One of them (0.06% of all analyzed sequences) did not appear to be a valid COI sequence; another two (0.13%), that looked valid, could not be assigned to any known taxonomic group and were probably caused by sequencing errors.
Predatory behaviour
Forty adult A. amphalodes were collected at Amanzi in order to investigate the prey-capture behaviour of the spider. After transfer to the laboratory, adult spiders of both sexes were placed separately into an arena consisting of 250 ml plastic bottles (8 cm in diameter, 15 cm tall) containing a 3 cm layer of sand in the bottom. They were left for three days to settle down at room temperature (~25 °C) and a natural LD (12:12) regime, during which time they were starved. Spiders usually dug themselves into the sand immediately after being released into the arena. Hodotermes mossambicus termites from one nest were collected in a suburban grassland in Langenhoven Park, Bloemfontein and kept together in plastic containers (40 cm in diameter) filled with soil. Trials began when a termite was introduced to the arena occupied by a spider. The hunting sequence was observed and recorded using handycam Canon Legria HF G10. If the spider did not start hunting within one hour, the prey was removed. We measured the paralysis latency as the time between successful attack and termite immobilization. After the trial the whole body length of termites and prosoma length of the spider were measured with a Nikon SMZ800 stereomicroscope.
Prey acceptance
The acceptance of different prey by A. amphalodes spiders was investigated in the lab. We used only adult females (N = 32) collected in the Amanzi with a mean body size of 5 mm as these are generally more voracious than males. The following prey that occurred syntopically with the spiders were used: termites (H. mossambicus workers, 9 mm body length, T. trinervoides workers, 4 mm, Reticulitermes sp., 3.5 mm), ants (Messor sp. workers, 7 mm; Anoplolepis custodiens (F. Smith) workers, 5 mm) and Tenebrionidae beetles (Zophosis boei Solier, 8 mm). Spiders were placed separately into the arenas (as above) and starved for two days. The prey were collected one day prior to running the trials. The prey was released into the container occupied by the spider and the result was recorded. In each trial we recorded whether the spider attacked and consumed the prey. When the spider did not attack the prey after an hour, the prey was removed from the dish and was replaced with a different prey. Each spider was offered all prey types in a randomised order. When the spider captured the prey the next prey was offered two days later. The design used was a complete block. To control for motivation to hunt the prey, H. mossambicus termites were offered to individuals that refused alternative prey (as a control). Only individuals that subsequently captured H. mossambicus termites were included in the analysis.
All analyses were performed within an R environment59. The composition of potential prey (i.e. relative frequency of each prey species in the field plots) was compared with the composition of the actual prey (i.e. relative frequency of spider with DNA of each prey species found in the gut) using the χ2 goodness of fit test. Smith’s index60 was used to estimate the width of the trophic niche. To compare the paralysis latency and the relationship between body size ratio and paralysis latency, we used General Linear Model (GLM) with Gamma error structure (GLM-g). GLM with Poisson error structure (GLM-p) was used to compare the number of sequences among juveniles, females and males.
Additional Information
Accession codes: The sequences used for primer design are available via GenBank (Accession numbers: KP748184-KP748191).
How to cite this article: Petráková, L. et al. Discovery of a monophagous true predator, a specialist termite-eating spider (Araneae: Ammoxenidae). Sci. Rep. 5, 14013; doi: 10.1038/srep14013 (2015).
References
Begon, M., Townsend, C. R. & Harper, J. L. Ecology: From Individuals to Ecosystems. 4th edn. (Blackwell Publishing, Oxford, UK, 2006).
Thompson, J. N. The coevolutionary process (University of Chicago Press, Chicago, USA, 1994).
Britt, E. J., Hicks, J. & Bennett, A. F. The energetic consequences of dietary specialisation in populations of the garter snake. Thamnophis elegans. J. Exp. Biol. 209, 3164–3169 (2006).
Nyffeler, M., Breene, R. G. & Dean, D. A. Facultative monophagy in the jumping spider, Plexippus paykulli (Audouin) (Araneae: Salticidae). Peckhamia 2(6), 92–96 (1990).
Líznarová, E., Sentenská, L., García, L. F., Pekár, S. & Viera, C. Local trophic specialisation in a cosmopolitan spider (Araneae). Zoology 116(1), 20–26 (2013).
Pekár, S. & Toft, S. Trophic specialisation in a predatory group: the case of prey-specialised spiders (Araneae). Biol. Rev. 90(3), 744–761 (2015).
McMurtry, J. A. & Croft, B. A. Life-styles of the Phytoseid mites and their roles in biological control. Annu. Rev. Entomol. 42, 291–321 (1997).
Li, D. & Jackson, R. R. Influence of diet on survivorship and growth in Portia fimbriata, an araneophagic jumping spider (Araneae: Salticidae). Can. J. Zool. 75, 1652–1658 (1997).
Slogget, J. J. & Majerus, M. E. N. Habitat preferences and diet in the predatory Coccinellidae (Coleoptera): an evolutionary perspective. Biol. J. Linnean Soc. 70, 63–88 (1999).
Yeargan, K. V. Ecology of a bolas spider, Mastophora hutchinsoni: phenology, hunting tactics and evidence for aggressive chemical mimicry. Oecologia 74, 524–530 (1988).
Pekár, S., Toft, S., Hrušková, M. & Mayntz, D. Dietary and prey-capture adaptations by which Zodarion germanicum, an ant-eating spider (Araneae: Zodariidae), specialises on the Formicinae. Naturwissenschaften 95(3), 233–239 (2008).
Coddington, J. A. & Levi, H. W. Systematics and evolution of spiders (Araneae). Annu. Rev. Ecol. Syst. 22, 565–592 (1991).
Pekár, S., Coddington, J. A. & Blackledge, T. A. Evolution of stenophagy in spiders (Araneae): evidence based on the comparative analysis of spider diets. Evolution 66(3), 776–806 (2012).
Wesołowska, W. & Cumming, M. S. Mashonarus guttatus, gen. and sp. n., the second termitivorous jumping spider from Africa (Araneae: Salticidae). Bull. British Arachn. Soc. 12, 165–170 (2002).
Eberhard, W. G. Chrosiotes tonala (Araneae, Theridiidae), a web-building spider, specializing on termites. Psyche 98, 7–19 (1991).
Jocqué, R. & Dippenaar-Schoeman, A. S. Two new termite-eating Diores species (Araneae: Zodariidae) and some observations on unique prey immobilization. J. Nat. Hist. 26, 1405–1412 (1992).
Van den Berg, A. & Dippenaar-Schoeman, A. S. Ground-living spiders from an area where the harvester termite Hodotermes mossambicus occurs in South Africa. Phytophylactica 23, 247–253 (1991).
Traniello, J. F. A. & Leuthold, R. H. Behavior and ecology of foraging in termites in Termites: Evolution, Sociality, Symbioses, Ecology (eds. Abe, T., Bignell, D. E., Higashi, M. ) 141–168 (Kluwer Academic Publishers, Dordrecht, Netherlands, 2000).
Dean, W. R. J. & Milton, S. The Karoo: Ecological Patterns and Proceses. (Cambridge University Press, Cambridge, UK, 1999).
Uys, V. A Guide to the Termite Genera of Southern Africa in Plant Protection Research Institute Handbook No. 15. (Agricultural Research Council, Pretoria, 2002).
Dippenaar-Schoeman, A. S., De Jager, M. & Van den Berg, A. Ammoxenus species (Araneae: Ammoxenidae)-specialist predators of harvester termites in South Africa. Afr. Plant Prot. 2, 103–109 (1996).
Dippenaar-Schoeman, A. S. & Harris, R. Food storage by a wandering ground spider (Araneae, Ammoxenidae, Ammoxenus). J. Arachnol. 33(3), 850–851 (2005).
Wilson, D. S. & Clark, A. B. Above ground predator defence in the harvester termite, Hodotermes mossambicus (Hagen). J. Entomol. Soc. South Africa 40, 271–282 (1977).
Dean, W. R. J. Spider predation on termites (Hodotermitidae). J. Entomol. Soc. South Africa 51, 147–148 (1988).
Dippenaar-Schoeman, A. S., De Jager, M. & Van den Berg, A. Behaviour and biology of two species of termite-eating spiders, Ammoxenus amphalodes and A. pentheri (Araneae: Ammoxenidae), in South Africa. Afr. Plant Prot. 2, 15–17 (1996).
Richardson, M. L. & Hanks, L. M. Partitioning of niches among four species of orb-weaving spiders in a grassland habitat. Environ. Entomol. 38(3), 651–656 (2009).
Symondson, W. O. C. Molecular identification of prey in predator diets. Mol. Ecol. 11, 627–641 (2002).
King, R. A., Read, D. S., Traugott, M. & Symondson, W. O. C. Molecular analysis of predation: a review of best practice for DNA-based approaches. Mol. Ecol. 17, 947–963 (2008).
Pompanon, F. et al. Who is eating what: diet assessment using next generation sequencing. Mol. Ecol. 21, 1931–1950 (2012).
Chapman, E. G., Schmidt, J. M., Welch, K. D. & Harwood, J. D. Molecular evidence for dietary selectivity and pest suppression potential in an epigeal spider community in winter wheat. Biol. Control 65, 72–86 (2013).
Brown, D. S. et al. Dietary competition between the alien Asian Musk Shrew (Suncus murinus) and a re-introduced population of Telfair’s Skink (Leiolopisma telfairii). Mol. Ecol. 23(15), 3695–3705 (2014).
Clare, E. L. et al. The diet of Myotis lucifugus across Canada: assessing foraging quality and diet variability. Mol. Ecol. 23(15), 3618–3632 (2014).
Zaidi, R. H., Jaal, Z., Hawkes, N. J., Hemingway, J. & Symondson, W. O. C. Can multiple-copy sequences of prey DNA be detected amongst the gut contents of invertebrate predators? Mol. Ecol. 8, 2081–2087 (1999).
Herbert, P. D. N., Ratnasingham, S. & deWaard, J. R. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B Biol. Sci. 270, S96–S99 (2003).
Zeale, M. R. K., Butlin, R. K., Barker, G. L. A., Lees, D. C. & Jones, G. Taxon specific PCR for DNA barcoding arthropod prey in bat faeces. Mol. Ecol. Res. 11, 236–244 (2011).
Piñol, J., San Andrés, V., Clare, E. L., Mir, G. & Symondson, W. O. C. A pragmatic approach to the analysis of diets of generalist predators: the use of next-generation sequencing with no blocking probes. Mol. Ecol. Res. 14(1), 18–26 (2014).
Razgour, O. et al. High-throughput sequencing offers insight into mechanisms of resource partitioning in cryptic bat species. Ecol. Evol. 1(4), 556–570 (2011).
Kruger, F. et al. An integrative approach to detect subtle trophic niche differentiation in the sympatric trawling bat species Myotis dasycneme and Myotis daubentonii. Mol. Ecol. 23(15), 3657–3671 (2014).
Deagle, B. E., Chiaradia, A., McInnes, J. & Jarman, S. N. Pyrosequencing faecal DNA to determine diet of little penguins: is what goes in what comes out? Conserv. Genet. 11, 2039–2048 (2010).
Huseynov, E. F. O. The prey of the lynx spider Oxyopes globifer (Araneae, Oxyopidae) associated with a semidesert dwarf shrub in Azerbaijan. J. Arachnol. 34, 422–426 (2006).
Huseynov E. F. O. Natural prey of the jumping spider Menemerus taeniatus (Araneae: Salticidae). Eur. J. Entomol. 102, 797–799 (2005).
Dippenaar-Schoeman, A. S. et al. First Atlas of the Spiders of South Africa (ARC-Plant Protection Research Institute, Pretoria, 2010).
Pekár, S., Král, J. & Lubin, Y. Natural history and karyotype of some ant-eating zodariid spiders (Araneae: Zodariidae) from Israel. J. Arachnol. 33(1), 50–62 (2005).
Pekár, S., Šedo, O., Líznarová, E., Korenko, S. & Zdráhal, Z. David and the Goliath: potent venom of an ant-eating spider (Araneae) enables capture of a giant prey. Naturwissenschaften 101(7), 533–540 (2014).
Luamba, J. L. N. Recherches sur le polymorphisme et aperçu sur l’influence de l’analogue de l’hormone juvénile sur le développement d’un termite Hodotermes mossambicus (Isoptera, Hodotermitidae). Biol. Ecol. Médit. 7, 169–171 (1980).
Endler, J. A. Interactions between predators and prey in Behavioural Ecology - An Evolutionary Approach (eds J. R., Krebs & N. B., Davies ) 169–196 (Blackwell Scientific Publications, Oxford, UK, 1991).
Ferry-Graham, L. A., Bolnick, D. I. & Wainwright, P. C. Using functional morphology to examine the ecology and evolution of specialization. Integr. Comp. Biol. 42, 265–277 (2002).
Allan, R. A., Elgar, M. A. & Capon, R. J. Exploitation of an ant chemical alarm signal by the zodariid spider Habronestes bradleyi Walckenaer. Proc. R. Soc. Lond. B Biol. Sci. 263, 69–73 (1996).
Cárdenas, M., Jiroš, P. & Pekár, S. Selective olfactory attention of a specialised predator to intraspecific chemical signals of its prey. Naturwissenschaften 99(8), 597–605 (2012).
Henschel, J. R. Psammophily in Namib Desert spiders. J. Arid Environ. 37, 695–707 (1997).
Sint, D., Raso, L., Kaufmann, R. & Traugott, M. Optimizing methods for PCR-based analysis of predation. Mol. Ecol. Res. 11(5), 795–801 (2011).
Hajibabaei, M. et al. A minimalist barcode can identify a specimen whose DNA is degraded. Mol. Ecol. Notes 6(4), 959–964 (2006).
Folmer, O., Black, M., Hoeh, W., Lutz, R. & Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299 (1994).
Tamura, K. et al. MEGA5: Molecular Genetics Analysis using Maximum Likelihood, Evolutionary Distance and Maximum Parsimony Methods. Mol. Biol. Evol. 28, 2731–2739 (2011).
Jarman, S. N. Amplicon: software for designing PCR primers on aligned DNA sequences. Bioinformatics 20, 1644–1645 (2004).
Giardine, B. et al. Galaxy: A platform for interactive large-scale genome analysis. Genome Res. 15, 1451–1455 (2005).
Hall, T. A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98 (1999).
Jones, M., Ghoorah, A. & Blaxter, M. jMOTU and Taxonerator: turning DNA barcode sequences into annotated operational taxonomic units. PLoS ONE 6, e19259 (2011).
R Development Core Team R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, Vienna, Austria, 2010). Available at http://www.R-project.org (Accessed: 15th December 2014).
Smith, E. P. Niche breadth, resource availability and inference. Ecology 63(6), 1675–1681 (1982).
Acknowledgements
We would like to thank S. Korenko and V. Butler for their help to collect spiders and termites in the field and Beth Clare for her advice with NGS data analysis. Field work for this study was funded through a grant from the National Research Foundation of South Africa through its Knowledge Interchange and Collaboration programme (Grant UID #83231). The lab study was funded by the Czech Science Foundation (GA15-14762S).
Author information
Authors and Affiliations
Contributions
S.P. and W.O.C.S. designed the study; S.P., E.L., L.S. and C.R.H. collected material at the study site; L.P. performed molecular analyses supervised by W.O.C.S.; E.L. and L.S. did behavioural experiments; S.P., E.L. and L.P. analysed data; all authors contributed to writing the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Petráková, L., Líznarová, E., Pekár, S. et al. Discovery of a monophagous true predator, a specialist termite-eating spider (Araneae: Ammoxenidae). Sci Rep 5, 14013 (2015). https://doi.org/10.1038/srep14013
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep14013
This article is cited by
-
Tracing the evolution of trophic specialisation and mode of attack behaviour in the ground spider family Gnaphosidae
Organisms Diversity & Evolution (2020)
-
Nest usurpation: a specialised hunting strategy used to overcome dangerous spider prey
Scientific Reports (2019)
-
Comparison of the capture efficiency, prey processing, and nutrient extraction in a generalist and a specialist spider predator
The Science of Nature (2018)
-
Making weed biological control predictable, safer and more effective: perspectives from New Zealand
BioControl (2018)
-
Landuse Change in Savannas Disproportionately Reduces Functional Diversity of Invertebrate Predators at the Highest Trophic Levels: Spiders as an Example
Ecosystems (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
