
Abstract
Chronic exposure to neonicotinoid insecticides has been linked to reduced survival of pollinating insects at both the individual and colony level, but so far only experimentally. Analyses of large-scale datasets to investigate the real-world links between the use of neonicotinoids and pollinator mortality are lacking. Moreover, the impacts of neonicotinoid seed coatings in reducing subsequent applications of foliar insecticide sprays and increasing crop yield are not known, despite the supposed benefits of this practice driving widespread use. Here, we combine large-scale pesticide usage and yield observations from oilseed rape with those detailing honey bee colony losses over an 11 year period and reveal a correlation between honey bee colony losses and national-scale imidacloprid (a neonicotinoid) usage patterns across England and Wales. We also provide the first evidence that farmers who use neonicotinoid seed coatings reduce the number of subsequent applications of foliar insecticide sprays and may derive an economic return. Our results inform the societal discussion on the pollinator costs and farming benefits of prophylactic neonicotinoid usage on a mass flowering crop.
Similar content being viewed by others


Introduction
Insects underpin the seed and fruit formation of pollinator-dependent crops that make up a critical fraction of the human diet1,2. Since 1961, the global cultivated area of insect-dependent crops has trebled3, while at the same time several key insect pollinator groups have declined4,5. This has led to concerns that pollination deficits may limit crop production6. Agricultural intensification reduces the diversity of food plants upon which insects depend7 and increases pollinators’ exposure to multiple pesticides8 that may act synergistically to increase insecticidal activity9. Neonicotinoids are potent nerve stimulants with a high affinity for insect nicotinic acetylcholine receptors10 and include the N-nitroguanidine group (clothianidin, imidacloprid or thiamethoxam) used as seed coatings and the less toxic11 N-cyanoamidine group (acetamiprid and thiacloprid) used as foliar insecticide sprays. Neonicotinoids are readily absorbed by plants, which transport them systemically providing pest protection throughout all plant tissues. The versatility of application and favourable pest control properties have contributed to neonicotinoids becoming the most widely used insecticides in the world12, with over 90% of usage in the form of seed coatings of clothianidin, imidacloprid or thiamethoxam13.
Oilseed rape (OSR; Brassica napus) is the most widely planted oilseed crop in Europe14 and provides a mass-flowering pollen and nectar resource highly attractive to many species of pollinator, including honey bees15,16. Neonicotinoid seed coating has become common practice on OSR across the UK in recent years13, offering the farmer a new class of insecticide active against important pests such as the cabbage stem flea beetle (Psylliodes chrysocephala) and the peach potato aphid (Myzus persicae; Fig. 1).

Neonicotinoid insecticides are widely used on oilseed rape in the United Kingdom to combat pests like the peach potato aphid
(Myzus persicae; A), which can carry damaging viruses such as Turnip yellows virus (TuYV; B) and cabbage stem flea beetle (Psylliodes chrysocephala) whose larvae cause stem damage (C). Oilseed rape represents a mass flowering nectar and pollen resource in the UK landscape for important pollinators such as the honey bee (D and E). Images A, C and D – Courtesy Food and Environment Research Agency; Image B – Courtesy Dr Mark Stevens, Rothamsted Research; Image E created using ESRI ArcGIS 10.1 from the ArcGIS Online World Imagery service.
Prophylactic application of neonicotinoids as a seed coating helps to protect the crop against pests for up to 10 weeks post planting17, reducing the need for subsequent weather-dependent applications of foliar insecticide sprays using older chemistries such as organophosphates, pyrethroids and carbamates, to which some pests have developed resistance18,19. However, neonicotinoid residues persist in plant tissues long enough to be detectable in OSR pollen and nectar20,21, providing a potential route for mass exposure to pollinators. Indeed the N-nitroguanidine neonicotinoids have been linked experimentally to changes in pollinator foraging behaviour22, reduced survival of individual insects23,24, decelerated colony growth25,26 and in the case of bumble bees, colony failure27. Concern that neonicotinoid seed coatings may be harming pollinators has led to a two year restriction on their use for mass flowering crops across the EU, starting in December 201328. However, real-world datasets that link the usage of neonicotinoid seed coatings to pollinator mortality are lacking at the agroecosystem level.
There is little doubt that pests such as cabbage stem flea beetle and the peach potato aphid and its associated viruses (Fig. 1) cause significant reductions in OSR yield in untreated crops17,29,30 and that neonicotinoid seed coatings provide a level of control for these pests30,31. However, applying a neonicotinoid as a seed coating costs farmers over three times more than a single foliar insecticide spraying32, yet the additional yield benefit of neonicotinoid seed coating — in comparison to pest control regimes that only use foliar insecticide applications — has never been quantified. Statements on the benefits of neonicotinoid usage on OSR seed assume that the protective benefits of neonicotinoid seed coatings post-planting avert the need for a farmer to apply a spray of foliar insecticides in the Autumn32. However, the foliar pesticide application behaviours of farmers who use neonicotinoid seed coatings have never been assessed. The honey bee (Apis mellifera) can be regarded as the most important commercial pollinator globally, responsible for at least 90% of commercial pollination33. Honey bees are the most frequent flower visitors to OSR6,15 and their pollinating activities contribute to seed pod weight and pod set6. Our study fuses large datasets that describe seed and foliar pesticide usage, land use, OSR yield, meteorological conditions and honey bee colony losses to provide a unique opportunity to investigate the potential costs to pollinators and benefits to farmers of neonicotinoid seed coatings on OSR at a national level.
Results
Exploring factors to explain honey bee colony losses
Datasets that described pesticide usage, land use, OSR yield, meteorological conditions and honey bee colony losses were successfully linked for each of nine regions across England and Wales from 2000 to 2010 (see Methods). Agricultural data from the June survey showed that the total cropped area of OSR for England and Wales doubled between 2000 (293,378 ha) and 2010 (602,270 ha; Fig. 2). National pesticide usage data indicated a steady increase in the use of neonicotinoid seed coatings, which began replacing the organochlorine Gamma HCH with <1% of the planted OSR area in 2000 and increased to cover over 75% of the planted OSR area in 2010 (Fig. 2).

Total area of oilseed rape cropped across England and Wales (ha) including the area treated with insecticide seed coatings.
More recent seed coated insecticide usage is dominated by the neonicotinoids clothianidin, imidacloprid and thiamethoxam, which replaced Gamma HCH.
Pesticide usage data, from between 6,000 ha and 18,000 ha of OSR annually, were the most spatially and temporally restricted, so all datasets were considered at the same regional, biennial scale (see Methods). Two region/year combinations (West Midlands in 2000 and Wales in 2006) were excluded due to missing pesticide usage data, leaving 52 region/year combinations (Supplementary Table S1). The proportion of in-season honey bee colony losses was derived from colony health inspections across England and Wales after excluding apiaries that contained notifiable disease (see Methods). In total, there were 126,220 colony observations, of which 10,725 honey bee colonies were found to be dead (8.5% colony losses; Supplementary Table S1).
First we used the proportion of honey bee colony losses as the response variable and region, meteorological conditions, summed neonicotinoid usage on OSR seed (to include clothianidin, imidacloprid and thiamethoxam; kg/m2) and density of OSR grown (m2/m2) as the explanatory variables in a quasi-binomial generalized linear model (see Methods). The model output indicated that honey bee colony mortality differed significantly due to region, SummerTempmin, SummerSun and SpringTempmax (Table 1). Neonicotinoid usage on OSR seed (p = 0.41) and density of OSR grown (p = 0.11) did not explain additional variation for honey bee colony mortality and were not added to the final model. A single value with high leverage (Wales in 2008) was removed from the model to investigate whether this point influenced the reported relationships, but the results after removal remained unchanged (data not shown).
Given the dominance of imidacloprid usage as an insecticide seed coating on OSR between 2000 and 2008 (Fig. 2), we repeated the above quasi-binomial generalized linear model substituting summed neonicotinoid usage on OSR seed (kg/m2) with that of only imidacloprid. Region remained the strongest predictor of honey bee colony loss, however once the differences between regions had been accounted for, imidacloprid usage on OSR (kg/m2) had a positive relationship with honey bee colony losses such that increased regional usage was linked to higher honey bee colony losses (Table 2; Fig. 3). A single value with a high residual (East Midlands in 2008) was removed from the model, but again the results remained unchanged (data not shown). Given the lack of clothianidin and thiamethoxam usage prior to 2010 (Fig. 2), it was not possible to analyse for an individual effect of these neonicotinoid seed coatings.

Predicted relationship between regional imidacloprid usage on oilseed rape (kg/m2) assessed biennially from 2000 to 2010 and the proportion of dead honey bee colonies after region had been taken into account (black line; n = 52; p = 0.001).
Red lines represent 95% confidence intervals.
We then checked for spurious results due to the possible correlation between the OSR density (m2/m2) and imidacloprid usage on OSR seed by forcing the model described in Table 2 to include OSR density before imidacloprid usage. The results indicated that honey bee colony loss was not significantly related to the density of OSR (deviance ratio = 2.43; df = 1, 41; p = 0.13) and the relationship between imidacloprid usage and honey bee colony losses remained significant (deviance ratio = 9.36; df = 1,41; p = 0.004), suggesting that our datasets contained sufficient temporal spread to decouple imidacloprid usage on OSR seed from the density of OSR grown.
Quantifying foliar insecticide sprays
Pesticide usage survey data were used to identify the number of foliar insecticide sprays associated with winter OSR crops between 2000 and 2010 (see Methods). Over 99% of all foliar insecticide applications were pyrethroids, including Cypermethrin (47%) Lambda-cyhalothrin (19%), Alpha-cypermethrin (12%), Tau-fluvalinate (9%), Zeta-cypermethrin (8%), Deltamethrin(4%) and Bifenthrin (1%). Carbamates (pirimicarb; 0.5%), organophosphates (Chlorpyrifos and Dimethoate; 0.1%) and neonicotinoids from the N-cyanoamidine group (Thiacloprid; 0.1%) were rarely used as foliar insecticide sprays. Pesticide usage records indicated that no neonicotinoids from the N-nitroguanidine group (clothianidin, imidacloprid or thiamethoxam) had been used as foliar insecticide sprays.
The number of seasonally applied foliar insecticide sprays (either autumn or during flowering) was used as the response variable in a Poisson regression model with a log link function with insecticide seed coating (none, clothianidin, imidacloprid or thiamethoxam) as an explanatory variable and region as a fixed effect (see Methods). Data were excluded for 2000 because of the low number of observations for imidacloprid (n = 2) and data on the spring planting of OSR were removed to focus on foliar insecticide sprays on the more common winter sown OSR. The number of foliar insecticide sprays was assessed for 4,230 fields totalling 73,051 ha of winter OSR crops across England and Wales (Table 3). Regression models predicted a consistent negative effect of imidacloprid seed treatment on the number of insecticide sprays applied in autumn in each year (Fig. 4A). This was supported by a mixed effect Poisson model combining data from across all years for imidacloprid seed treated crops (p < 0.001; Fig. 4A). A positive effect was noted for imidacloprid seed treatment on the number of insecticide sprays during flowering in 2004 (Fig. 4B), however a mixed effect Poisson model combining data from across all years for imidacloprid seed treated crops suggested no consistent increase in the number of foliar insecticide applications during flowering (p = 0.430: Fig. 4B).

Estimated effect of imidacloprid (grey), clothianidin (clear) and thiamethoxam (hatched) seed treatments on the number of foliar insecticide sprays used during autumn
(A) and flowering (B) on winter oilseed rape when compared to crops that received no insecticide seed treatment. Estimates, p-values and 95% confidence intervals are derived from Poisson models for each individual year or mixed effect Poison models for all years (imidacloprid only;*).
Yield benefits of neonicotinoid seed coatings
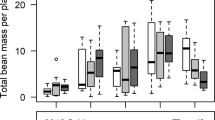
Yield (t ha−1) was used as the response variable in a linear regression with insecticide seed coating (none, clothianidin, imidacloprid or thiamethoxam) as an explanatory variable and region, the proportion of farm-saved seed and the proportion of hybrid seed as fixed effects (see Methods). Not all fields assessed for the number of foliar insecticide applications had associated yield data, leading to a slightly reduced dataset for yield comparisons. The yield (t ha−1) was assessed for 2801 fields totalling 47,120 ha of winter OSR crops across England and Wales (Table 3). Linear models predicted a positive effect of neonicotinoid seed treatment on yield (t ha−1) for 2004, 2006 and 2010 (thiamethoxam only; Fig. 5). However, a negative effect on yield was predicted for imidacloprid seed treated crops in 2008 when compared to crops that received no insecticide seed treatment (Fig. 5). A linear mixed model combining data from across all years found no overall effect of imidacloprid seed treatment on yield (p = 0.248; Fig. 5).

Estimated effect of imidacloprid (grey), clothianidin (clear) and thiamethoxam (hatched) seed treatments on oilseed rape yield (t ha−1) when compared to crops that received no insecticide seed treatment.
Estimates, p-values and 95% confidence intervals are derived from linear models for each individual year or a mixed effect linear model for all years (imidacloprid only;*).
Partial budget analysis of neonicotinoid seed coatings
We completed a partial budget analysis for using prophylactic neonicotinoid seed coatings by combining our new data on yield and foliar insecticide usage with industry figures on insecticide treatment costs for farmers (see Methods)32. The results of the budget analysis mirrored the yield, with positive of neonicotinoid seed treatment on profit (£ ha−1) for 2004, 2006 and 2010 (thiamethoxam only) and a negative effect in 2008 (Fig. 6). A linear mixed model combining data from across all years found no overall effect of imidacloprid seed treatment on farming profits (p = 0.248; Fig. 6).

Estimated effect of imidacloprid (grey), clothianidin (clear) and thiamethoxam (hatched) seed treatments on farming profit (£ ha−1) when compared to crops that received no insecticide seed treatment.
Estimates, p-values and 95% confidence intervals are derived from linear models for each individual year or a mixed effect linear model for all years (imidacloprid only;*).
Discussion
The farming benefit of using neonicotinoid seed coatings to reduce subsequent foliar insecticide sprays and increase crop yield for OSR was not known prior to this study, despite this practice being widespread. We combined pesticide usage and yield observations from over 76,000 ha of OSR over five replicate years to provide the first evidence that farmers who used imidacloprid seed coatings consistently reduced the number of foliar insecticide sprays used in the autumn, but not during flowering (Fig. 4). Our observations fit well with the known efficacy of such seed coatings against cabbage stem flea beetle (up to 4–5 weeks34) and peach potato aphid (up to 10 weeks17) and demonstrate for the first time that farmers do alter their foliar pesticide application practices as a result of using imidacloprid seed coatings.
Previous work has demonstrated the efficacy of imidacloprid seed coating against peach potato aphids in OSR, however the resulting yield of seed-treated plots was not found to be different from the untreated controls29. Indeed, previous studies have failed to explore the yield benefit of neonicotinoid seed coating in comparison to pest control regimes that only use foliar insecticide applications. For the first time we tested whether farmers using neonicotinoid seed treatments increase yield (t ha−1) or profit (£ ha−1) when compared to OSR crops grown with no seed treatment but with foliar insecticides. Our results demonstrate, that whilst farmers did experience significant yield increases in 2004 and 2006 for imidacloprid and in 2010 for thiamethoxam, there was no consistent effect of imidacloprid seed coating on yield over all years (Fig. 5). Indeed the effects of imidacloprid seed treatment on the yield of oilseed rape were negative in 2008, perhaps suggesting reduced pest control compared to crops that received no insecticide seed treatment. This curious result could be explained by a failure of the prophylactic neonicotinoid seed treatment to control pests, perhaps due to leaching after unprecedented heavy rainfall across many parts of the UK during late summer 200735. Thiamethoxam has been shown to leach from soil with simulated high rainfall36 and increased irrigation of seed treated potatoes was shown to reduce imidacloprid residual levels in leaf tissues37. It therefore seems plausible that the efficacy of imidacloprid seed treatments may have been compromised in 2008. Interestingly, significant reductions in autumnal foliar insecticide sprays in crops treated with neonicotinoid seed coatings did not appear to greatly influence farming profit, which instead appeared to be driven by OSR yield (compare Figs 5 and 6).
We also present the first evidence of a relationship between increasing imidacloprid usage on OSR seed and escalating honey bee colony losses at a landscape level. Given the generalised nature of our datasets and the multitude of unaccounted variables that are known to impact honey bee colony mortality38, it is surprising that our approach detected a link between imidacloprid usage and honey bee colony loss. Our data were not derived from a controlled experiment and so may be influenced by confounding factors not accounted for in our models, such as honey bee pests. However, controlled studies have severe logistical constraints because the required manipulations focus at the landscape scale39. For example, honey bees have been observed flying between control and neonicotinoid treated sites over 10 km apart in agricultural landscapes40, placing significant practical restrictions on site selection. Our observational data include colony health measures from 126,220 colonies over 11 years and predicted differences in colony mortality of 10% between low and high field exposure of imidacloprid (Fig. 3; Table 2). Our data would support the suggestion of Cresswell41 that published experiments attempting to link neonicotinoid usage with poor honey bee health40,42 lack the statistical power to discover similar population level effects on colony mortality. Our observational data inform Hill’s epidemiological criteria43 of consistency (repeatable association between the putative cause and its consequence over space and time) and provide evidence of a biological gradient44. Taken together with a growing body of evidence that imidacloprid alters the foraging behaviour and survival23,24 of insects, we provide important new supporting evidence that highlights the need to investigate causal relationships between imidacloprid usage at the landscape level and honey bee decline.
If the correlation between imidacloprid seed treatment on OSR and honey bee colony losses is a result of a causal link, then the direct exposure of honey bees after foraging on the nectar and pollen from treated OSR offers a likely mechanism. Nectar taken directly from treated OSR have been shown to contain imidacloprid in the range of 0.6–2.0 ppb20 and pollen from honey bees in the USA has been shown to have near ppm levels8. Furthermore, feeding choice experiments recently suggested that honey bees may prefer to consume sucrose solutions containing nectar-realistic concentrations of imidacloprid, leading to concerns that preferential foraging on toxin containing nectar could lead to higher colony exposure45. Estimated dissipation times (DT50) for imidacloprid in soil vary widely13 making it difficult to predict whether repeated applications in successive years could result in an accumulation of product in soil. Nevertheless, typically more than 90% of the seed coating enters the soil13 and an analyses of soil from UK field detected imidacloprid up to 10.7 ppb, despite no record of growing imidacloprid treated crops for three years prior to sampling46. Pollinator-friendly flowering plants growing in field margins of neonicotinoid treated crops have also been shown to contain 1–10 ppb levels in flowers47, suggesting another likely route of oral exposure.
More recently imidacloprid usage has declined (Fig. 2), replaced by second generation neonicotinoids with different metabolic fates in honey bees. Whilst clothianidin, thiamethoxam and imidacloprid are all toxic to honey bees, metabolites of imidacloprid, such as olefin, have higher toxicity than the parent compound and the timing of their production has been linked to honey bee mortality48. Toxic metabolites of clothianidin and thiamethoxam have not been reported in honey bees (with the exception of conversion of thiamethoxam to clothianidin49) and such differences mean that further investigations are required to determine whether the risks for second generation neonicotinoids are significantly different to imidacloprid – parity of risk cannot be assumed. The fact that summed usage of clothianidin, imidacloprid and thiamethoxam failed to explain additional variation in honey bee colony loss could be an artefact of a single year’s sampling (2010; Fig. 2) and this analysis would benefit from the inclusion of more recent usage data. Our model suggested that honey bee colony losses differed significantly between regions and were linked negatively to changes in summer sunshine and spring temperature, but positively to summer temperature. Honey bee colony losses are known to differ between regions in a single year50, however our data suggest certain regions, such as Wales, have consistently high losses over time (Tables 1 and 2). Temperature and light intensity have a clear impact on the ability of honey bees to forage and sexually reproduce. Worker honey bees do not begin to forage for pollen and nectar in the spring until temperatures rise above 9 °C and foraging activity increases as temperature and light intensity rises51. Drone (male) honey bees are less likely to depart on mating flights with low light intensity52 and honey bee queens mated in poor weather mate with fewer drones53 and are more frequently superseded54. Reports on weather influencing honey bee colony losses are rare, but colony losses have been associated with unseasonably cool and wet weather in early spring55, supporting our observed link between low spring temperatures and honey bee colony losses (Table 1).
For the first time we are able to present the costs and benefits of prophylactic neonicotinoid use on OSR for the farmer alongside an accompanying link to landscape level honey bee colony loss. Neonicotinoid seed coatings provide only partial control of pests and viruses56 and resistance in some pest groups57 will reduce their efficacy. Risk assessments assuming total control in the presence of a seed coating versus apocalyptic yield losses in their absence are simplistic and perhaps over-state the benefits32,58. Our data provide numerical evidence on the potential costs (lost honey bee colonies and sometimes lower yield) relative to benefits (reduced number of foliar insecticide sprays and sometimes yield increase) associated with using neonicotinoid seed coatings on OSR. It appears that the economic justification for using neonicotinoid to treat OSR seed in our model system is dynamic and sometimes financially beneficial to farmers. Our data contribute to the growing body of evidence highlighting the need for a large scale field-based experiment to determine the real-world impacts on pollinators of the use of neonicotinoid seed coatings on mass flowering crops. As long as field-applied acute toxins remain the basis of agricultural pest control practices, society will repeatedly be forced to weigh the benefits of pesticides against their collateral environmental damage. Nowhere is this tension more evident than in the system we describe here with the world’s most widely used insecticide, the world’s most widely used managed pollinator and Europe’s most widely grown mass flowering crop.
Methods
Exploring factors to explain honey bee colony losses
Pesticide usage surveys of arable crops were completed biennially between 2000 and 2010 by visiting selected holdings stratified from across nine regions of England and Wales (East Midlands, Eastern, London & South East, North East, North West, South West, Wales, West Midlands, Yorkshire & the Humber)59. Total neonicotinoid usage on OSR seed (kg/m2) was calculated by summing clothianidin, imidacloprid and thiamethoxam seed treatments from national pesticide usage surveys59. Regional cropped OSR area and OSR density (m2/m2) were derived from the June Agricultural Survey biennially between 2000 and 201060. National Bee Unit (NBU) Inspectors assessed the health of honey bee colonies belonging to registered beekeepers across England and Wales between April and September from 2000 to 2010. NBU inspectors operate a combined prioritised, non-random, risk-based inspection protocol for the control of notifiable brood diseases and surveillance for exotic pest incursions61. Whilst surveillance visits for exotic pests comprise a random element, apiary visits for notifiable disease can be spatially clustered and diseased colonies are sometimes destroyed. As such, visits where notifiable brood diseases (American or European foulbrood) were discovered were removed from the dataset (approximately 8% of observations). Honey bee hives that contained no live adult honey bees upon inspection were classed as dead colonies. A range of different factors can influence honey bee colony losses, including pests and pathogens, for a recent review see62. Whilst the ectoparasitic mite Varroa destructor and its associated viruses have been shown to cause honey bee colony losses (for review see63), no records of these or any other honey bee pests and diseases were gathered systematically for the period of interest. Honey bee colony losses can be influenced by poor weather conditions during foraging periods55, we therefore purchased quarterly meteorological data that covered the majority of the foraging period for honey bees from the Met Office archive of gridded climate data created by interpolating monthly station data as described previously64. Measurements of meteorological conditions included average daily mean (Tempmean), minimum (Tempmin) and maximum (Tempmax) temperature (°C), total rainfall (Rain; mm); and total sunshine (Sun; h) for Spring (Apr, May, Jun) and Summer (Jul, Aug, Sep) for each region/year combination.
A quasi-binomial generalized linear model with a logit link function was constructed using the proportion of honey bee colony losses as the response variable and region, meteorological conditions (SpringTempmax, SpringTempmean, SpringTempmin, SpringSun, SpringRain, SummerTempmax, SummerTempmean, SummerTempmin, SummerSun, SummerRain), neonicotinoid usage on OSR seed (kg/m2) and density of OSR grown (m2/m2) as the explanatory variables using Genstat V17.1 (VSN International).
Quantifying foliar insecticide sprays
The number of foliar insecticide spray applications and active ingredients for winter sown OSR crops were derived from biennial pesticide usage surveys from 2000 to 2010. Timing of insecticide spray application was separated into Autumn (Sept-Nov) and Flowering (Mar-Jul). The number of seasonally applied foliar insecticide sprays (either autumn or during flowering) was used as the response variable in a Poisson regression model with a log link function with insecticide seed coating (none, clothianidin, imidacloprid or thiamethoxam) as an explanatory variable and region as a fixed effect. In addition, a mixed effect Poisson regression model was constructed based on data from all years for imidacloprid as a single explanatory variable. The mixed model included a random intercept as well as a random treatment effect grouped by year in order to account for variation in foliar insecticide applications between years. Results were expressed as the effect size between neonicotinoid seed treatment and seed receiving no insecticide seed treatment. Analyses were run using R version 3.0.2 for Windows61 and mixed effect models fitted using the lme4 package for R version 1.0.762.
Yield benefits of neonicotinoid seed coatings
Yields (t ha−1) were derived directly from a random subset of farmers during pesticide usage surveys. Linear models (for each individual year) and mixed effect linear models (imidacloprid use across all years) were constructed in the same way as described above for the number of foliar insecticide sprays, except that the proportion of farm-saved seed and the proportion of hybrid seed were added as additional fixed effects.
Partial budget analysis of neonicotinoid seed coatings
A partial budget analysis was conducted by multiplying the yield (t ha−1) by the fixed sales price of £327.13 t using the July 2007 to mid-April 2013 average delivered Erith OSR price32. The costs of applying neonicotinoid seed coatings was subtracted assuming treatment costs of £16.50, £12.90 and £9.55 per ha respectively for farm-saved, conventional and hybrid seed32. The cost of applying an exemplar foliar insecticide (deltamethrin) was taken to be £4.75 per spray per ha32. Fuel and time costs were assumed to be fixed due to the normal practice of using tank mixes that contain herbicides and/or fungicides when applying foliar insecticides32. Linear models (for each individual year) and a mixed effect linear model (imidacloprid use across all years) were constructed in the same way as described above for yield.
Additional Information
How to cite this article: Budge, G.E. et al. Evidence for pollinator cost and farming benefits of neonicotinoid seed coatings on oilseed rape. Sci. Rep. 5, 12574; doi: 10.1038/srep12574 (2015).
Change history
20 August 2015
An uncorrected version of this article was inadvertently made available on the Scientific Reports website on 13 August 2015. The corrected, final version was published 20 August 2015.
References
Ollerton, J., Winfree, R. & Tarrant, S. How many flowering plants are pollinated by animals? Oikos 120, 321–326 (2011).
Eilers, E. J., Kremen, C., Greenleaf, S. S., Garber, A. K. & Klein, A. M. Contribution of Pollinator-Mediated Crops to Nutrients in the Human Food Supply. Plos One 6, 10.1371/journal.pone.0021363 (2011).
Aizen, M. A. & Harder, L. D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr Biol 19, 915–918 (2009).
Forister, M. L. et al. Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. PNAS 107, 2088–2092 (2010).
Biesmeijer, J. et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313, 351–354 (2006).
Garratt, M. et al. The identity of crop pollinators helps target conservation for improved ecosystem services. Biol Conserv 169, 128–135 (2014).
Barr, C. J. et al. Countryside Survey 1990: Main Report (Countryside 1990 vol. 2). (London, 1993).
Mullin, C. A. et al. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Honey Bee Health. Plos One 5, 10.1371/journal.pone.0009754 (2010).
Bingham, G. et al. Temporal synergism can enhance carbamate and neonicotinoid insecticidal activity against resistant crop pests. Pest Manag Sci 64, 81–85 (2008).
Tomizawa, M. & Casida, J. E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu Rev Pharmacol 45, 247–268 (2005).
Laurino, D., Porporato, M., Patetta, A. & Manino, A. Toxicity of neonicotinoid insecticides to honey bees: laboratory tests. Bull Insectology 64, 107–113 (2011).
Jeschke, P., Nauen, R., Schindler, M. & Elbert, A. Overview of the Status and Global Strategy for Neonicotinoids. J Agr Food Chem 59, 2897–2908 (2011).
Goulson, D. REVIEW: An overview of the environmental risks posed by neonicotinoid insecticides. J Appl Ecol 50, 977–987 (2013).
Carre, P. & Pouzet, A. Rapeseed market, worldwide and in Europe. OCL - Oilseeds and Fats, Crops and Lipids 21, D102–D102 (2014).
Hayter, K. E. & Cresswell, J. E. The influence of pollinator abundance on the dynamics and efficiency of pollination in agricultural Brassica napus: implications for landscape-scale gene dispersal. J Appl Ecol 43, 1196–1202 (2006).
Westphal, C., Steffan-Dewenter, I. & Tscharntke, T. Mass flowering crops enhance pollinator densities at a landscape scale. Ecol Lett 6, 961–965 (2003).
Jones, R., Coutts, B. & Hawkes, J. Yield-limiting potential of Beet western yellows virus in Brassica napus. Aust J Agr Res 58, 788–801 (2007).
Bass, C. et al. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem Molec 51, 41–51 (2014).
Zimmer, C. T., Mueller, A., Heimbach, U. & Nauen, R. Target-site resistance to pyrethroid insecticides in German populations of the cabbage stem flea beetle, Psylliodes chrysocephala L. (Coleoptera: Chrysomelidae). Pestic Biochem Phys 108, 1–7 (2014).
Pohorecka, K. et al. Residues of Neonicotinoid Insecticides in Bee Collected Plant Materials from Oilseed Rape Crops and Their Effect on Bee Colonies. J Apic Sci 56, 115–134 (2012).
EFSA. Statement on the findings in recent studies investigating sub-lethal effects in bees of some neonicotinoids in consideration of the uses currently authorised in Europe. EFSA Journal 10, 2752 (2012).
Feltham, H., Park, K. & Goulson, D. Field realistic doses of pesticide imidacloprid reduce bumblebee pollen foraging efficiency. Ecotoxicology 23, 317–323 (2014).
Schneider, C. W., Tautz, J., Gruenewald, B. & Fuchs, S. RFID Tracking of Sublethal Effects of Two Neonicotinoid Insecticides on the Foraging Behavior of Apis mellifera. Plos One 7, 10.1371/journal.pone.0030023 (2012).
Henry, M. et al. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 336, 348–350 (2012).
Whitehorn, P. R., O’Connor, S., Wackers, F. L. & Goulson, D. Neonicotinoid Pesticide Reduces Bumble Bee Colony Growth and Queen Production. Science 336, 351–352 (2012).
Sandrock, C. et al. Impact of Chronic Neonicotinoid Exposure on Honeybee Colony Performance and Queen Supersedure. Plos One 9, 10.1371/journal.pone.0103592 (2014).
Gill, R. J., Ramos-Rodriguez, O. & Raine, N. E. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 491, 105–108 (2012).
EU. EU-485/2013: Amending implementing regulation (EU) No 540/2011. http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32013R0485 (2013) (Date of access: 04/11/2014).
Dewar, A., Tait, M. & Stevens, M. Efficacy of thiamethoxam seed treatment against aphids and turnip yellows virus in oilseed rape. Asp Appl Biol, 195–202 (2011).
Soroka, J. J., Grenkow, L. F. & Irvine, R. B. Impact of decreasing ratios of insecticide-treated seed on flea beetle (Coleoptera: Chrysomelidae, Phyllotreta spp.) feeding levels and canola seed yields. J Econ Entomol 101, 1811–1820 (2008).
Birch, P. A. & Nicholson, T. A new insecticidal seed treatment for oilseed rape. In: Seed treatment: challenges & opportunities, British Crop Protection Council, 27–32 (2001).
Nicholls, C. J. Implications of the restriction on the neonicotinoids: imidacloprid, clothianidin and thiamethoxam on crop protection in oilseeds and cereals in the UK. Research Review No. 77. (2013).
Allsopp, M. H., de Lange, W. J. & Veldtman, R. Valuing Insect Pollination Services with Cost of Replacement. Plos One 3, 10.1371/journal.pone.0003128 (2008).
Kazda, J., Baranyk, P. & Nerad, D. The implication of seed treatment of winter oilseed rape. Plant Soil Environ 51, 403–409 (2005).
Prior, J. & Beswick, M. The exceptional rainfall of 20 July 2007. Weather 63, 261–267, 10.1002/wea.308 (2008).
Gupta, S., Gajbhiye, V. T. & Gupta, R. K. Soil dissipation and leaching behavior of a neonicotinoid insecticide thiamethoxam. Bul Environ Contam Tox 80, 431–437, 10.1007/s00128-008-9420-y (2008).
Prager, S. M., Vindiola, B., Kund, G. S., Byrne, F. J. & Trumble, J. T. Considerations for the use of neonicotinoid pesticides in management of Bactericera cockerelli (Sulk) (Hemiptera: Triozidae). Crop Prot 54, 84–91, 10.1016/j.cropro.2013.08.001 (2013).
vanEngelsdorp, D. et al. A national survey of managed honey bee 2010-11 winter colony losses in the USA: results from the Bee Informed Partnership. J Apicult Res 51, 115–124 (2012).
vanEngelsdorp, D. et al. Standard epidemiological methods to understand and improve Apis mellifera health. J Apicult Res 52, 10.3896/ibra.1.52.4.15 (2013).
Cutler, G. C., Scott-Dupree, C. D., Sultan, M., McFarlane, A. D. & Brewer, L. A large-scale field study examining effects of exposure to clothianidin seed-treated canola on honey bee colony health, development and overwintering success. Peer J 2, e652–e652 (2014).
Cresswell, J. E. A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology 20, 149–157 (2011).
Pilling, E., Campbell, P., Coulson, M., Ruddle, N. & Tornier, I. A Four-Year Field Program Investigating Long-Term Effects of Repeated Exposure of Honey Bee Colonies to Flowering Crops Treated with Thiamethoxam. Plos One 8, 10.1371/journal.pone.0077193 (2013).
Hill, A. B. The Environment and disease: Association or Causation? P Roy Soc Med 58, 295–300 (1965).
Cresswell, J. E., Desneux, N. & vanEngelsdorp, D. Dietary traces of neonicotinoid pesticides as a cause of population declines in honey bees: an evaluation by Hill’s epidemiological criteria. Pest Manag Sci 68, 819–827 (2012).
Kessler, S. C. et al. Bees prefer foods containing neonicotinoid pesticides. Nature 521, 74–76 (2015).
Jones, A., Harrington, P. & Turnbull, G. Neonicotinoid concentrations in arable soils after seed treatment applications in preceding years. Pest Manag Sci 70, 1780–1784, 10.1002/ps.3836 (2014).
Krupke, C. H., Hunt, G. J., Eitzer, B. D., Andino, G. & Given, K. Multiple Routes of Pesticide Exposure for Honey Bees Living Near Agricultural Fields. Plos One 7, 10.1371/journal.pone.0029268 (2012).
Suchail, S., Debrauwer, L. & Belzunces, L. P. Metabolism of imidacloprid in Apis mellifera. Pest Manag Sci 60, 291–296 (2004).
Nauen, R., Ebbinghaus-Kintscher, U., Salgado, V. L. & Kaussmann, M. Thiamethoxam is a neonicotinoid precursor converted to clothianidin in insects and plants. Pestic Biochem Phys 76, 55–69 (2003).
Steinhauer, N. A. et al. A national survey of managed honey bee 2012-2013 annual colony losses in the USA: results from the Bee Informed Partnership. J Apicult Res 53, 1–18, 10.3896/ibra.1.53.1.01 (2014).
Burrill, R. M. & Dietz, A. The response of honey bees to variations in solar-radiation and temperature. Apidologie 12, 319–328, 10.1051/apido:19810402 (1981).
Neves, E. F., Faita, M. R., Gaia, L. D. O., Alves Junior, V. V. & Antonialli-Junior, W. F. Influence of Climate Factors on Flight Activity of Drones of Apis mellifera (Hymenoptera: Apidae). Sociobiology 57, 107–113 (2011).
Woyke, J. Causes of repeated mating flights by queen honeybees. J Apicult Res 3, 7 (1964).
Winston, M. The biology of the honey bee. (Harvard University Press, 1991).
Wilson, W. & Menapace, D. Disappearing disease of honey bees: a survey of the United States. Am Bee J 119, 118–217 (1979).
Coutts, B., Webster, C. & Jones, R. Control of Beet western yellows virus in Brassica napus crops: infection resistance in Australian genotypes and effectiveness of imidacloprid seed dressing. Crop Pasture Sci 61, 321–330 (2010).
Puinean, A. M. et al. Amplification of a Cytochrome P450 Gene Is Associated with Resistance to Neonicotinoid Insecticides in the Aphid Myzus persicae. Plos Genetics 6, 10.1371/journal.pgen.1000999 (2010).
Noleppa, S. & Hahn, T. The value of neonicotinoid seed treatment in the European Union: a socioeconomic, technological and environmental review, HFFA working paper 01/2013 (2013).
Garthwaite, D. G., Thomas, M. R., Parrish, G., Smith, L. & Barker, I. Pesticide usage survey report 224: Arable crops in Great Britain. (2008).
Defra. June Census of Agriculture and Horticulture (Land Use and Livestock on Agricultural Holdings at 1 June 2010) UK - Final Results. (2010) (Date of access: 01/02/2012).
Wilkins, S., Brown, M. A. & Cuthbertson, A. G. S. The incidence of honey bee pests and diseases in England and Wales. Pest Manag Sci 63, 1062–1068 (2007).
Vanbergen, A. J. & Initiative, T. I. P. Threats to an ecosystem service: pressures on pollinators. Front Ecol Environ 11, 251–259 (2013).
Rosenkranz, P., Aumeier, P. & Ziegelmann, B. Biology and control of Varroa destructor. J invertebr pathol 103 Suppl 1, S96–119, 10.1016/j.jip.2009.07.016 (2010).
Perry, M. & Hollis, D. The generation of monthly gridded datasets for a range of climatic variables over the UK. Int J Climatol 25, 1041–1054 (2005).
Acknowledgements
The authors were funded jointly by grants from BBSRC, Defra, NERC, the Scottish Government and the Wellcome Trust, under the Insect Pollinators Initiative (grants BB/I000119/1, BB/I000429/1 and BB/I000801/1). Long-term datasets on honey bee colony health and the June Agricultural Survey were funded by Defra and the Welsh Government. Collection of Pesticide Usage Survey data were funded by Chemicals Regulation Directorate. The authors would also like to thank Simon Kightley from NIAB for providing a list of hybrid and conventional OSR cultivars.
Author information
Authors and Affiliations
Contributions
G.E.B., S.P. and N.D.B. conceived the project; A.C. provided oilseed rape coverage data from the June Agricultural Survey; D.G. contributed the Pesticide Usage Survey data; G.E.B., M.A.B. contributed the honey bee loss data; S.P. and H.H.T. completed all analyses in discussion with the remaining authors; G.E.B. and K.S.D. wrote the first draft of the manuscript and all authors contributed to the final version.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Budge, G., Garthwaite, D., Crowe, A. et al. Evidence for pollinator cost and farming benefits of neonicotinoid seed coatings on oilseed rape. Sci Rep 5, 12574 (2015). https://doi.org/10.1038/srep12574
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep12574
This article is cited by
-
Imidacloprid seed treatment in soybean-associated arthropod food webs: Reason for concern, or justifiable neglect?
Journal of Pest Science (2023)
-
Honeybee colonies compensate for pesticide-induced effects on royal jelly composition and brood survival with increased brood production
Scientific Reports (2021)
-
An update of the Worldwide Integrated Assessment (WIA) on systemic insecticides. Part 3: alternatives to systemic insecticides
Environmental Science and Pollution Research (2021)
-
An update of the Worldwide Integrated Assessment (WIA) on systemic insecticides. Part 2: impacts on organisms and ecosystems
Environmental Science and Pollution Research (2021)
-
Late effect of larval co-exposure to the insecticide clothianidin and fungicide pyraclostrobin in Africanized Apis mellifera
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
