
Abstract
Co-injection of zygotes with Cas9 mRNA and sgRNA has been proven to be an efficient gene-editing strategy for genome modification of different species. Genetic engineering in pigs holds a great promise in biomedical research. By co-injection of one-cell stage embryos with Cas9 mRNA and Npc1l1 sgRNA, we achieved precise Npc1l1 targeting in Chinese Bama miniature pigs at the efficiency as high as 100%. Meanwhile, we carefully analyzed the Npc1l1 sgRNA:Cas9-mediated on- and off-target mutations in various somatic tissues and ovaries and demonstrated that injection of zygotes with Cas9 mRNA and sgRNA is an efficient and reliable approach for generation of gene-modified pigs.
Similar content being viewed by others

Introduction
Pigs are considered as one of the best animals for generating models of human diseases and for providing organs for xenotransplantation, because they share similar features with humans in physiology, anatomy and lifespan. Genetically modified pigs have been demonstrated to hold a great promise in biomedical research1,2. However, precise gene modifications of the pig genome are challenging, because traditional gene modifications by homologous recombination (HR) have only been demonstrated to be efficient in authentic embryonic stem cells (ESC) that are not available in pigs3. Historically, specific gene knockout strategies in pigs were based on DNA homologous recombination (HR) in somatic cells, followed by somatic cell nuclear transfer (SCNT)4.
However, the efficiency of HR in somatic cells is extremely low, which is one of the main barriers in gene targeting in pigs5,6. Recently, programmable endonucleases, such as Zinc Finger Nucleases (ZFNs), Transcription activator-like effector nucleases (TALENs) and especially the CRISPR, the recently developed clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) 9 system, were shown to be a kind of revolutionary technologies for targeted genome editing7. Indeed, these technologies greatly advanced the gene targeting in pigs by HR-based SCNT8,9,10,11,12,13,14,15. Nevertheless, SCNT is limited by the technical challenges, such as that the porcine fetal fibroblasts used as donors for SCNT have a finite proliferative capacity16, SCNT sometimes results in abnormal animals and the efficiencies of producing offspring by SCNT are low5,17. Hence, more efficient and reliable strategies are required for gene targeting in pigs. Genome modifications by direct injection of zygotes with DNA or mRNA of ZFNs and TALENs opened a new avenue for precise gene modification in different species18,19,20. This strategy has been successfully used in pigs21.
Compared to ZFNs and TALENs, CRISPR/Cas9-mediated genome engineering is easy to handle, highly specific, efficient and multiplexable22. Taking the advantages of CRISPR/Cas9, we achieved efficient gene targeting in mice, rats and monkeys by co-injection of one-cell stage embryos with Cas9 mRNA and sgRNAs23,24,25, which encouraged us to extend the application of this strategy to gene targeting in pigs and especially, to further characterize this strategy by thorough analysis of the Cas9/sgRNA-mediated target modification in different tissues, especially the gonads, although using CRISPR/Cas9 to produce gene-modified pigs via direct injection of zygotes has been proved26,37.
To do this, the Chinese Bama miniature pig was chosen as the model animals. Chinese Bama miniature pig is one of the widely used large model animals for bio-medical research in China, especially for pharmacological experiments27,28,29, because of its genetic stability, high degree of inbreeding, small size, easiness to handle and it shares anatomical, metabolic and physiological similarities with humans. Npc1l1 (Niemann-Pick C1-Like 1) was chosen as the target gene. The results showed, through co-injection of early stage embryos of Chinese Bama miniature pigs with Npc1l1 sgRNA and Cas9 mRNA, target bi-allelic modifications of pig Npc1l1 were achieved at the efficiency as high as 100%. Meanwhile, we first characterized the Cas9/sgRNA-mediated on- and off-target mutation in various somatic tissues and ovaries and first provided detailed information about injection of zygotes with Cas9 mRNA and sgRNA. We demonstrated that this CRISPR/Cas9-based approach is efficient and reliable for generation of gene-modified pigs.
Results and Discussion
The Design and Validation of sgRNA Targeting Pigs
NPC1L1 is highly expressed in small intestine and plays a critical role in both dietary cholesterol absorption and biliary cholesterol reabsorption30,31,32. In mice, NPC1L1 is exclusively expressed in small intestine and gallbladder, but human NPC1L1 is highly expressed in liver, intestine and perhaps other tissues. The tissue distribution of NPC1L1 expression in pigs was unknown. Considering similarities in physiology and metabolism between pigs and humans, we generated NPC1L1-deficient pig models for future definition of how NPC1L1 influences cardiovascular and metabolic diseases. One sgRNA targeting the exon 2 of Npc1l1 (Figure 1a) was designed as described previously22.

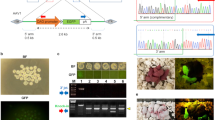
Evaluation of Npc1l1 sgRNA:Cas9-mediated modifications of Npc1l1 in pig parthenogenetic embryos.
(a) Schematic diagram of pig Npc1l1 partial protein coding region and the targeting locus of Npc1l1 sgRNA:Cas9. Red arrow indicates the targeting site of Npc1l1 sgRNA:Cas9. (b) Sequencing results of the modified Npc1l1 alleles detected in injected parthenogenetic embryos. Sequences complementary to sgRNA are labeled in red and PAM sequences are in green; mutations, blue, lower case; deletions, (−); insertions, (+). (c) The chromatographs of sequencing modified Npc1/1 alleles in parthenogenetic embryos in which overlapped peaks were observed. Con denotes wild-type sequence. Red arrow indicates the targeting site of Npc1l1 sgRNA:Cas9.
Although Cas9 mRNA and sgRNA have been successfully applied to embryos of several species33, they had not been tested in pigs when this work was initiated. But the fact that microinjection of zygotes with ZFN or TALEN generated knockout pigs21, suggested it was highly possible that Cas9 mRNA and sgRNA would work in pig embryos. To determine if Cas9 mRNA and sgRNA system works in pig embryos, we transcribed the Cas9 and Npc1l1 sgRNA in vitro using T7 RNA polymerase as described previously23. Twenty nanograms of Cas9 mRNA and 10 nanograms of sgRNA were pooled and microinjected into cytoplasm of 195 pig parthenogenetic embryos as described by Wang et al. 39. (Table 1). A total of 28 embryos normally developed to blastocyst at even a slightly higher efficiency of cleavage (the experimental group: 52.8% (103/195) v.s. the control group: 42.4% (91/225); P = 0.014). The blastocyst development rate in the experimental group was comparable to that of the control group (14.4% (28/195) v.s. 13.3% (30/225); P = 0.778; Table 1), suggesting that Cas9 mRNA and Npc1l1 sgRNA were not toxic to the embryos of pigs.
We isolated the pig genomic DNA samples from a total of 8 individual embryos at 144 hr post microinjection and screened for the presence of site-specific gene modification by PCR amplification of the region around the target site followed by T7EN1 cleavage assays using the PCR products as substrates. The cleavage bands were visible in the target gene (Data not shown). Further characterization of the cleavage by sequencing the PCR products revealed that 3 out of 8 samples displayed overlapped peaks in the sequencing chromatographs (test embryos #1, #7 & #8) and 4 out of 8 samples exhibited distinguishable insertions or deletions (indels) with variable mutation sizes (−11 ~ +29 bp) (test embryos #2, #3, #4 & #6) at the target site (Figure 1b & c and Table 2). These data demonstrated that the selected sgRNA worked well with Cas9 on the target gene in pig embryos.
Cas9/sgRNA Enables One-Step Gene Modification in Pigs
With the success in pig parthenogenetic embryos, we set out to generate knockout Chinese Bama miniature pigs. A total of 110 porcine early embryos (1-cell stage) were surgically collected from 20 mated sows. The Cas9 mRNA and Npc1l1 sgRNA mixtures were injected into cytoplasm of embryo cells as described above. A total of 105 out of 110 injected embryos were transferred into 4 surrogate females. Of the 4 recipient mothers, 2 pregnancies were established (50%; 2 out of 4). After full-term (~114 days) pregnancy, 2 litters (Litter 1: C1-1 ~ C1-5; Litter 2: C2-1 ~ C2-6) of 12 piglets were successfully delivered alive, but one died immediately after birth (Figure 2a and Table 3).

Detection of Npc1l1sgRNA:Cas9-mediated modifications of NPC1L1 in founder pigs.
(a) A photo showing 38-day-old pigs carrying Npc1l1 mutations. (b) PCR products of the targeted region of Npc1l1 from founder pigs. The first litter of five live pigs were named C1-1 to C1-5. The second litter of six live pigs were named from C2-1 to C2-6. Con denotes wild-type pig as a control. Notably, the PCR products of C1-3, C1-4, C2-1, C2-4 and C2-6 differ from the control in size, suggesting a large fragment deletion or insertion. (c) Detection of Npc1l1 sgRNA:Cas9-mediated on-target cleavage of Npc1l1 by T7EN1 cleavage assays. All PCR products from (b) were subjected to T7EN1 cleavage assays. All the samples were digested by T7EN1, suggesting that all founders carry Npc1l1 mutations. (d) Sequencing results of modified Npc1l1 alleles detected in founder pigs. At least 12 TA clones of the PCR products were analyzed. Sequences complementary to sgRNA are labeled in red and PAM sequences in green. Mutations, blue, lower case; deletions, (−); insertions, (+). N/N indicates positive colonies out of total sequenced samples. See also Figure S1.
To genotype the piglets, the ear punch tissues of the eleven live infant pigs were collected and the genomic DNA was isolated. The Npc1l1 sgRNA:Cas9-mediated genome modifications were first screened by PCR and T7EN1 cleavage assay using the genomic DNA as described above. The additional bands were observed by PCR amplification of the target region in 5 infants, including C1-3, C1-4, C2-1, C2-4 and C2-6 (Figure 2b), suggesting that the genomic modification occurred in these founder animals with large indels. Then, the PCR products of all the founders were subjected to the T7EN1 cleavage assay (Figure 2c). Very excitingly, the cleavage products were observed in all the 11 infants, indicating efficient genomic modifications in the founder piglets. As expected, different kinds of indels were detected by sequencing the PCR products (Figure 2d, Table 4 and Supplementary Figure 1), further confirming the efficient genomic modifications. Incredibly, no wild-type sequence was detected in all of the founders, indicating that CRISPR/Cas9 induced bi-allelic mutations at an extremely high efficiency in pig embryos.
Cas9/sgRNA-Mediated Genome Targeting Extensively Integrates into Different Tissues of Pigs
All of the targeting results described above were from the noninvasively available ear tissues. The dead founder pig provided us an opportunity to evaluate the integration of the Cas9/sgRNA-mediated Npc1l1 targeting into the derivatives of three germ layers, which generated detailed information for CRISPR/Cas9-mediated genome targeting in pigs via injection of zygotes with Cas9/sgRNA mixture. By PCR amplification and T7EN1 cleavage assays, we first performed extensive analysis of the target mutagenesis in 7 different somatic tissues, including heart, liver, spleen, lung, kidney, skin and muscle from the dead founder. While the PCR bands did not differ between experimental and control groups (Figure 3a), but the T7EN1 assays showed that the cleavage bands existed in every reactions (Figure 3b), indicating that the Cas9/sgRNA-mediated mutations occurred in all tissues examined. The target mutations were further confirmed by sequencing the samples from ear, heart and liver, which are derived from ectoderm, mesoderm and endoderm, respectively and the results showed that identical genetic modifications extensively integrated into all three germ layers (Figure 3c and Table 5). Again, no wild type allele was detected, indicating that CRISPR/Cas9-mediated bi-allelic mutations existed in all tissues examined. These results further substantiate the high efficiency of CRISPR/Cas9 system in pigs.

Detection of Npc1l1 sgRNA:Cas9-mediated targeting in different tissues.
(a) PCR products of the targeted region of Npc1l1 from 7 different tissues. (b) Detection of Npc1l1 sgRNA:Cas9-mediated on-target cleavage of Npc1l1. All PCR products from (a) were subjected to T7EN1 cleavage assays. (c) Sequencing results of modified Npc1l1 alleles detected in ear, heart and liver. Sequences complementary to sgRNA are labeled in red and PAM sequences in green. Mutations, blue, lower case; deletions, (−); insertions, (+). N/N indicates positive colonies out of total sequenced.
Cas9/sgRNA-Mediated Genome Targeting Efficiently Transmits into Ovaries of Pigs
To establish genetically modified animals requires germline transmission of genetic mutations. Because the gene-modified Bama miniature pigs have not reached sexual maturity, we currently cannot perform breeding to determine germline transmission. But the highly efficient occurrences of the target mutations in different tissues strongly suggest it is very possible that the Cas9/sgRNA-mediated genome targeting will integrate into the pig gonads. To prove the possibility, the founder C2-5 was sacrificed and the ovaries were isolated. The examination of the target mutagenesis was performed as above. The PCR and T7EN1 assays revealed that the cleavage of Npc1l1 did occur in the ovaries (Figure 4a & b). This was confirmed by sequencing of the target mutations, which were identical to those detected in the somatic tissues, such as a 1-bp insertion, 9-, 13- and 24-bp deletions (Figure 4c and Table 6) and absence of wild-type sequence, demonstrating the Cas9/sgRNA-mediated genome targeting successfully integrated into the gonads too. It is notifying, the gonads are composed of germ cells and somatic cells. The sequencing results of the ovaries showed that the efficiency of Cas9/sgRNA-induced targeting was 100% in the ovaries, strongly suggesting that the Cas9/sgRNA-mediated targeting was most likely transmitted into the pig germ cells. Nevertheless, breeding results will provide direct evidence of germline transmission in the future.

Npc1l1 sgRNA:Cas9-mediated modifications in the ovaries of founder C2-5.
(a) PCR products of the targeted region of Npc1l1 in the ovaries of founder C2-5. (b) Detection of Npc1l1 sgRNA:Cas9-mediated cleavage of Npc1l1 in ovaries. All PCR products from (a) were subjected to T7EN1 cleavage assays. (c) Sequencing results of modified Npc1l1 alleles detected in the ovaries of founder C2-5. PCR products of the targeted region of Npc1l1 were amplified from the ovaries and sequenced. The PAM sequences are highlighted in green; the targeting sequences in red; the mutations are shown in blue, lower case. Deletions, (−); and Insertions, (+). N/N indicates positive colonies out of total sequenced.
Mosaicism Analysis
The sequence data of both live and dead founder animals revealed multiple genotypes (Figure 2d, 3c and 4c; Table 4 ~ 6 and Supplementary Figure 1), suggesting that the CRISPR/Cas9-mediated cleavage had occurred multiple times and resulted in mosaicism of the modification as seen in other species23,24,25.
Further characterization of the mosaicism showed that a total of 18 kinds of indels with mutation sizes ranging from −412 bp to +535 bp were detected in different founders. Surprisingly, the 9 bp deletions were detected in 8 of the 11 founders, suggesting the possible preference of the Cas9/sgRNA-mediated Npc1l1 targeting. Of the total 11 founder piglets, 6 (C1-1, C1-2, C1-3, C1-5, C2-2 and C2-4) harbored with 2, 3 (C1-4, 2-1 and C2-3) with 3 and 2 (C2-5 and C2-6) with 4 different modifications (Table 4). Furthermore, the same modifications were detected in different tissues (Figure 3 and Table 4 & 5, Supplementary Figure 1) and ovaries (Figure 4c; Table 6 and Supplementary Figure 1) of the same founder, indicating that the modification occurred at very early embryogenesis.
Off-Target Detection
Off-target mutagenesis is of a major concern for CRISPR/Cas9 system33,34,35. The CRISPR/Cas9-induced off-target mutation is heritable both in mice and rats23,24. We thus ask whether off-target mutation occurred in our genetic modified pigs. To this end, we screened the pig genome using the open SeqMap tool and predicted a total of 8 most potential off-target sites (OT 1 ~ 8) (Supplementary Table 1). Using the primers listed in Supplementary Table 4, all the selected OTs were subjected to PCR amplification and T7EN1 cleavage assay. Results showed that except the OTs 2 and 4, no genetic modification was detected at the selected OTs (Figure 5a & b, Supplementary Figure 2). At the OTs 2 and 4, distinguishable cleavage bands of unexpected size were detected in 7 reactions of founders C1-1, C1-2, C1-5, C2-4, C2-6, including 2 (Founders C2-4 and C2-6) for OT2 and 5 (Founders C1-1, C1-2, C1-5, C2-4 and C2-6) for OT 4 (Figure 5b), indicating that the Npc1l1 sgRNA:Cas9 might have induced non-specific mutations at these two sites.

Detection of the Npc1l1 sgRNA:Cas9-mediated off-target cleavages in vivo.
(a) PCR products of the potential off-target sites 2 and 4 (OTs 2 and 4) of Npc1l1 sgRNA:Cas9 from founder pigs. (b) Detection of Npc1l1 sgRNA:Cas9-mediated off-target cleavage at the OTs 2 and 4. All PCR products from (a) were subjected to T7EN1 cleavage assays. *, different cleavage band patterns compared to the control. The size of the digested bands does not match that expected. (c) Sequencing results of PCR products. No off-target indel mutation was observed in the T7EN7 cleavage bands. The two red arrows indicate potential off-target sites of OT2 and OT4, respectively. Con denotes the corresponding sequence of each retrieved from Ensembl.
To confirm the Npc1l1 sgRNA:Cas9-mediated off-target cleavage events, PCR products from C1-1 for OT2 and C2-4 for OT 4 were randomly chosen for sequencing. Surprisingly, no real off-target mutations were detected and the unexpected cleavages came from SNPs (Figure 5c). These results demonstrated that CRISPR/Cas9 did not induce detectable off-target mutations in this study. Nevertheless, more comprehensive study may be required to clarify the off-target effects of these pigs. Considering the off-target mutagenesis is site-dependent and more specific strategies for organisms using mutated Cas9 has already been established36, the off-target mutagenesis can be minimized by optimizing the procedure.
In summary we show here that the site-specific pig gene modification can be efficiently achieved by co-injection of Cas9 mRNA and sgRNAs into the one-cell embryos. Furthermore, by careful characterization of the Npc1l1 sgRNA:Cas9-mediated on- and off-target effects, the mosacism, et al., we first provide detailed information about the Cas9/sgRNA-mediated target mutations via direct injection of zygotes in various somatic tissues, as well as the gonads, confirming the CRISPR/Cas9 system is versatile for heritable pig genome targeting without detectable off-target mutagenesis. During preparation of this manuscript, a similar study was reported37, which supports our conclusion that direct injection of zygotes with Cas9/RNA is an efficient and reliable approach for pig genome targeting.
Methods
Animals
The animals used in this study were regularly maintained in the Laboratory Animal Centre of the Third Military Medical University. All the protocols involving the use of animals were in accordance with approved guidelines of the Institutional Animal Care and Use Committee of the Third Military Medical University (Approval ID: SYXK-PLA-2007036).
Vector constructs and in vitro transcription
To construct the recombinant vector for preparation of sgRNA by in vitro transcription, the two complementary DNA oligos shown in Supplementary Table 2 were annealed to be double-stranded and subcloned into pUC57-T7-gRNA vector as described23. Using the constructed recombinant vector that was completely linearized by the endonuclease DraI as the templates, sgRNAs were produced via in vitro transcription using MEGAshortscript kit (Ambion) and purified using MEGAClear kit (Ambion) as described in the manuals. Using the Cas9 mRNA in vitro transcription vector (Addgene No. 44758) as templates, Cas9 mRNAs were produced and purified as described previously by Shen et al.23.
Cas9/sgRNA efficacy test via pig parthenogenetic embryo injection
Pig cumulus-oocyte complexes (COCs) were harvested from ovaries collected from slaughter houses. The COCs were cultured for in vitro maturation as described5,38,39. The pig oocytes were freed of cumulus by treating the cultured COCs with hyaluronidase and the matured oocytes with extruded polar body were selected out and subjected to cytoplasmic microinjection with Cas9/sgRNA mixture as described39. The injected oocytes were activated by direct current electrical pulses (1.2 KV/cm, 30 μs, two times, 1 sec interval) and the activated oocytes (parthenogenetic embryos) were cultured in PZM-3 media as described by Wang et al.39. The cleavage rate of parthenogenetic embryos was counted at 48 hr post activation and blastocystes harvested at 144 hr post activation. Pig genomic DNA was extracted from single parthenogenetic blastocysts by incubating individual embryos in lysis buffer as described39. Using the genomic DNAs as templates, a primer pair set (pNpc1l1-exon2-sgR-F: 5-GACCTACGAGTCCTGCAGC-3; pNpc1l1-exon2-sgR-R: 5-GAAGACCGAGCAGAGGATGA-3; product size: 386 bp) were used to amplify modified Npc1l1 alleles in injected embryos by PCR and the amplification products were subjected to T7EN1 analysis or sequencing after purification using gel extraction kit (Qiagen).
Production of gene-modified pigs via zygote injection with Cas9/sgRNA
Cas9 mRNA and sgRNA were mixed at the final concentrations of 20 and 10 ng/μL, respectively. Pig zygotes were surgically collected from mated sows as described40. The collected zygotes were subjected to cytoplasmic microinjection with the Cas9/sgRNA mixture in the same way as that for parthenogenetic embryos described above. Shortly after injection, the injected zygotes were transferred into synchronized foster mother sows as described40. Pregnancy was investigated by observing the oestrus behaviors of recipient sows at every ovation circle.
T7EN1 cleavage assay and sequencing
Different samples were collected and digested in a lysis buffer (0.4 M NaCl, 2 μM EDTA, 1% SDS, 10 μM Tris-HCl and 100 μg/ml Proteinase K). The genomic DNA of the sample was extracted from lysate by phenol-chloroform and recovered by alcohol precipitation. T7EN1 cleavage assay was performed as described by Shen et al23. Briefly, the targeted fragments were amplified by PrimerSTAR HS DNA polymerase (TaKaRa, DR010A) from the genomic DNA, then purified with a PCR cleanup kit (Axygen, AP-PCR-50). The primers for amplifying Npc1l1 targeted fragments were listed in Supplementary Table 3. The purified PCR product was denatured and re-annealed in NEBuffer 2 (NEB) using a thermocycler. The PCR products were digested with T7EN1 (NEB, M0302L) for 30 min at 37°C and separated on a 2.5% agarose gel. The PCR products with mutations detected by T7EN1 cleavage assay were sub-cloned into T vector (Takara, D103A). For each sample, the colonies were picked up randomly and sequenced by M13F (−47) primer (M13F (−47): 5′-CGC CAG GGT TTT CCC AGT CAC GAC-3′).
Off-target assay
To determine the site-specific cleavage of the CRISPR-Cas9 system in vivo, the potential off-target loci were searched by an open tool, SeqMap41. The mismatch parameter of target sequence was set as described25. ‘NGG’ and ‘NAG’ were chosen as PAM. The sites that have conserved 7 bp proximal to PAM with total mismatches < 5 and the sites with total mismatches < 4 were chosen as potential off-target sites for subsequent test. The selected potential off-target sites were PCR amplified using genomic DNA as templates. The PCR products were first subjected to T7EN1 cleavage assay. The potential off-target sites yielding typical cleavage bands were considered as candidates. The PCR products of the candidates were cloned and sequenced to confirm the off-target effects. The primer pairs used were listed in Supplementary Table 4.
Statistical analysis
Chi-square Test was performed in two-tailed manner to evaluate the difference of parthenogenetic embryo cleavage rate and blastocyst development rate between the Cas9/sgRNA injection and untreated groups.
References
Fahrenkrug, S. C. et al. Precision genetics for complex objectives in animal agriculture. J. Anim. Sci. 88, 2530–2539 (2010).
Tan, W. S., Carlson, D. F., Walton, M. W., Fahrenkrug, S. C. & Hackett, P. B. Precision editing of large animal genomes. Adv. Genet. 80, 37–97 (2012).
Capecchi, M. R. Altering the genome by homologous recombination. Science 244, 1288–1292 (1989).
Clark, A. J. & Whitelaw, C. B. A. A future for transgenic livestock. Nat. Rev. Genet. 4, 825–833 (2003).
Lai, L. et al. Production of α-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science 295, 1089–1092 (2002).
Rogers, C. S. et al. Disruption of the CFTR gene produces a model of cysticfibrosis in newborn pigs. Science 321, 1837–1841 (2008).
Kim, H. & Kim, J. S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 15, 321–334 (2014).
Bao, L. et al. Generation of GGTA1 biallelic knockout pigs via zinc-finger nucleases and somatic cell nuclear transfer. Sci. China Life Sci. 57, 263–268 (2014).
Carlson, D. F. et al. Efficient TALEN-mediated gene knockout in livestock. Proc. Natl. Acad. Sci. USA 109, 17382–17387 (2012).
Hauschild, J. et al. Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases. Proc.Natl. Acad. Sci. USA 108, 12013–12017 (2011).
Kwon, D. N. et al. Production of biallelic CMP-Neu5Ac hydroxylase knock-out pigs. Sci. Rep. 3, 1981; 10.1038/srep01981 (2013).
Tan, W. S. et al. Efficient nonmeiotic allele introgressionin livestock using custom endonucleases. Proc. Natl. Acad. Sci. USA 110, 16526–16531 (2013).
Xin, J. et al. Highlyefficient generation of GGTA1 biallelic knockout inbred minipigs with TALENs. PLoS ONE 8, e84250 (2013).
Yang, D. et al. Generation of PPARγ mono-allelic knockout pigs via zinc-finger nucleases and nuclear transfer cloning. Cell Res. 21, 979–982 (2011).
Zhou, X. et al. Generation of CRISPR/Cas9-mediated gene-targeted pigs via somatic cell nuclear transfer. Cell Mol. Life Sci. s00018-014-1744-7; 10.1007/s00018-014-1744-7 (2014).
Zhu, H. et al. Influence of tissue origins and external microenvironment on porcine foetal fibroblast growth, proliferative life span and genome stability. Cell Prolif. 37, 255–266 (2004).
Carter, D. B. et al. Phenotyping of transgenic cloned piglets. Cloning Stem Cells 4, 131–145 (2002).
Bogdanove, A. J. & Voytas, D. F. TAL effectors: customizable proteins for DNA targeting. Science 333, 1843–1846 (2011).
Carroll, D. et al. Gene targeting in Drosophila and Caenorhabditis elegans with zinc-finger nucleases. Methods Mol. Biol. 435, 63–77 (2008).
Urnov, F. D., Rebar, E. J., Holmes, M. C., Zhang, H. S. & Gregory, P. D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 11, 636–646 (2010).
Lillico, S. G. et al. Live pigs produced from genome edited zygotes. Sci. Rep. 3, 02847; 10.1038/srep02847 (2013).
Mali, P., Esvelt, K. M. & Church, G. M. Cas9 as a versatile tool for engineering biology. Nat. Methods 10, 957–963 (2013).
Shen, B. et al. Generation of gene-modified mice via Cas9/RNA-mediated gene targeting. Cell Res. 23, 720–723 (2013).
Ma, Y. et al. Generating rats with conditional alleles using CRISPR/Cas9. Cell Res. 24, 122–125 (2014).
Niu, Y. et al. Generation of gene-modified cynomolgus monkey via Cas9/RNA-mediated gene targeting in one-cell embryos. Cell 156, 836–843 (2014).
Whitworth, K. M. et al. Use of the CRISPR/Cas9 system to produce genetically engineered pigs from in vitro-derived oocytes and embryos. Biol. Reprod. 91, 78 (2014).
Puccinelli, E., Gervasi, P. G. & Longo. V. Xenobiotic metabolizing cytochrome P450 in pig, a promising animal model. Current Drug Metabolism 12, 507–525 (2011).
Shang, H., Yang, J., Liu, Y. & Wei, H. Tissue distribution of CYP3A29 mRNA expressionin Bama miniature pig by quantitative reversetranscriptase-polymerase chain reaction (RT-PCR). Xenobiotica. 39, 423–429 (2009).
Liu, Y., Zeng, B. H., Shang, H. T., Cen, Y. Y. & Wei, H. Bama miniature pigs (Sus scrofa domestica) as a model for drug evaluation for humans: comparison of in vitro metabolism and in vivo pharmacokinetics of lovastatin. Comp. Med. 58, 580–587 (2008).
Altmann, S. W. et al. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 303, 1201–1204 (2004).
Temel, R. E. et al. Hepatic Niemann-Pick C1-like 1 regulates biliary cholesterol concentration and is a target of ezetimibe. J. Clin. Invest. 117, 1968–1978 (2007).
Davis, H. R., Jr et al. Niemann-Pick C1 Like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 279, 33586–33592 (2004).
Sander, J. D. & Joung, J. K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 32, 347–355 (2014).
Hsu, P. D. et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 31, 827–32 (2013).
Pattanayak, V. et al. High-throughput profiling of off-target DNA cleavage reveals RNA-programmed Cas9 nuclease specificity. Nat. Biotechnol. 31, 839–843 (2013).
Shen, B. et al. Efficient genome modification in mice by CRISPR/Cas9 nickase without off-target effects. Nat. Methods 11, 399–402 (2014).
Hai, T., Teng, F., Guo, R., Li, W. & Zhou, Q. One-step generation of knockout pigs by zygote injection of CRISPR/Cas system. Cell Res. 24, 372–375 (2014).
Betthauser, J. et al. Production of cloned pigs from in vitro system. Nat. Biotechnol. 18, 1055–1059 (2000).
Wang, Y. et al. The meganuclease I-sceI containing nuclear localization Signal (NLS-I-sceI) efficiently mediated mammalian germline transgenesis via embryo cytoplasmic microinjection. PLoS ONE 9, e108347 (2014).
Whitelaw, C. B. et al. Efficient generation of transgenic pigs using equine infectious anaemia virus (EIAV) derived vector. FEBS Letter 571, 233–236 (2004).
Jiang, H. & Wong, W. H. SeqMap: mapping massive amount of oligonucleotides to the genome. Bioinformatics 24, 2395–2396 (2008).
Acknowledgements
The authors also thank the entire Huang Lab and Wei Lab for their support and advice. Y.W., H.W. and J.L. are supported by National Natural Science Foundation of China (31171280, 81173126 & 31201081), Y.L. is supported by National Science & Technology Support Program (2011BAI15BO2) and National High-tech R&D Program of China (863 Program) (2012AA020603). L.Y. is supported by Award Number R01DK085176 from the National Institute of Health, USA.
Author information
Authors and Affiliations
Contributions
H.W., X.H., J.G. and L.Y. designed the experiments and wrote the manuscript. Y.W., Y.D., B.S., X.Z., J.L., Y.L., J.W., J.Z., B.H. and N.K. performed the experiments.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Wang et al., Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Wang, Y., Du, Y., Shen, B. et al. Efficient generation of gene-modified pigs via injection of zygote with Cas9/sgRNA. Sci Rep 5, 8256 (2015). https://doi.org/10.1038/srep08256
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep08256
This article is cited by
-
One-step base editing in multiple genes by direct embryo injection for pig trait improvement
Science China Life Sciences (2022)
-
Comparison of the effects of introducing the CRISPR/Cas9 system by microinjection and electroporation into porcine embryos at different stages
BMC Research Notes (2021)
-
Strategies to reduce genetic mosaicism following CRISPR-mediated genome edition in bovine embryos
Scientific Reports (2019)
-
Genome mutation after introduction of the gene editing by electroporation of Cas9 protein (GEEP) system in matured oocytes and putative zygotes
In Vitro Cellular & Developmental Biology - Animal (2019)
-
Cattle with a precise, zygote-mediated deletion safely eliminate the major milk allergen beta-lactoglobulin
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
