
Abstract
This study investigated a typical secondary Betula platyphylla forest in the Ziwuling Mountains, Loess Plateau, China. In the sample plot, the DBH (diameter at breast height) class structure of B. platyphylla was bimodal. Individuals with small and large DBH values were abundant. The DBH structures of Quercus wutaishanica and Pinus tabulaeformis were close to that of the logistic model, thus suggesting the increasing population of these species. B. platyphylla and Populus davidiana showed random spatial distributions at almost all scales. However, Q. wutaishanica and P. tabulaeformis were significantly clumped at small scales. B. platyphylla had a negative spatial relation with Q. wutaishanica at small spatial scales. P. tabulaeformis and Q. wutaishanica showed negative spatial correlations at small scales, but they had positive correlations at large scales. These results suggest that P. tabulaeformis and Q. wutaishanica shared habitat preferences at these scales. In the future, the secondary B. platyphylla forest in the Ziwuling Mountains in the Loess Plateau will probably change into a multi-species mixed forest (Quercus–Pinus mixed forest). Assisted restoration strategies must be employed to improve the regeneration dynamics of the forest in the long term.
Similar content being viewed by others

Introduction
The Loess Plateau of China (LPC) faces the worst soil erosion problems in the world, given that approximately 72.3% of the total 6.5 × 105 km2 area is eroded. The serious soil erosion in the LPC has hampered local development because of the resulting low land productivity, frequent droughts and floods, deteriorating ecological condition and fast species decline1,2. Moreover, the erosion area is currently increasing at a rate of 900 km2 to 1100 km2 a year1.
The destruction of vegetation is considered the main cause of the erosion problems in the LPC3,4,5,6. The forest coverage of the LPC has decreased from 53% to 6.1% (mid-20th) over the last 25 centuries because of land reclamation7. Therefore, forest conservation and restoration research in the LPC are necessary to address the soil erosion problems.
As the largest natural secondary forest-covered region in the LPC, the Ziwuling Mountains play an important role in the control of soil erosion and climatic regulation in Northwest China. Betula platyphylla, which is a species of deciduous hardwood, is widely distributed in mid-high mountains of warm temperate regions in the world, including northern China, Russian Far East, Siberia, Mongolia, Northern Korea and Japan. B. platyphylla is also the most important pioneer woody species for the recovery of LPC vegetation. Some previous studies have explained the succession patterns in the LPC based on observations of similar communities in other areas8,9. However, these studies could not accurately predict the development of secondary forests in the LPC because of the endemic climate and habitats.
The spatial distribution pattern and population structure of trees are the direct results of comprehensive interactions among species characteristics, environmental factors and intraspecific and interspecific plant–plant interactions over a long period4,10. The spatial distribution patterns of trees depend on the plant's biological characteristics at a small scale and on environmental heterogeneity, such as soil, water, light and terrain, at large scales1,10,11. Considering the dual attributes (temporal and spatial scales) of spatial distribution pattern and population structure, we study a typical B. platyphylla community (B. platyphylla forests are one of the most common forests in the Ziwuling Mountains) to understand the biological characteristics of a secondary forest community and the interspecific relationship among the tree species in such community with spatial distribution and diameter class structure. The intrinsic reasons for the pattern formation and fluctuating population are also discussed. This study aims to provide a scientific basis for vegetation dynamics that aid in the interpretation of the secondary communities in the LPC and in the development of restoration strategies for other arid regions in the world characterized by plant disturbance and regeneration.
Results
Species composition of the forest and population structure of main tree species
The forest in the study area had a distinct layer system of trees, shrubs and grasses. The following tree species were identified in the sample plot: such as B. platyphylla, Quercus wutaishanica, Pinus tabulaeformis, Populus davidiana, Acer ginnala, Carpinus turczaninowii, Swida walteri, Robinia pseudoacacia and Toxicodendron vernicifluum (Table 1). In addition, 19 shrub and 35 grass species were found in sample plots, with the most abundant species being Rosaceae, Gramineae and Compositae (Table 1).
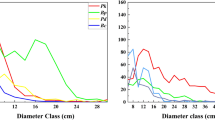
A total of 937 individuals were recorded in study plots. The dominant tree species were B. platyphylla, Q. wutaishanica, P. davidiana and P. tabulaeformis, which accounted for 49.62%, 36.18%, 3.84% and 7.90%, respectively, of the total number of individuals. According to the polynomial fitting result (Figure 1, polynomial order = 6), B. platyphylla showed a bimodal DBH distribution and young (DBH < 8 cm) and large trees (16 cm ≤ DBH < 26 cm) were more abundant than medium trees (10 cm ≤ DBH < 16 cm). The DBH distribution of P. davidiana approximated a Gaussian model (R2 = 0.92). P. davidiana trees thrived in the middle diameter classes (10 cm ≤ DBH < 16 cm), thereby suggesting a stable population. The DBH of Q. wutaishanica and P. tabulaeformis showed reverse J distributions, which suggested compatibility with an expanding trend in population size10.

DBH structures of main tree species in sample plots.
BP: Betula platyphylla; QW: Quercus wutaishanica; PD: Populus davidiana; PT: Pinus tabulaeformis.
Spatial pattern analysis of main tree species
An isogram of the main tree species was obtained by Kriging interpolation analysis using the tree coordinates (Figure 2). The distribution of B. platyphylla trees was continuous. Q. wutaishanica trees had slightly low continuity. P. davidiana and P. tabulaeformis had a patchy distribution.

Kriging interpolation analysis of individual distributions of main tree species.
BP: Betula platyphylla; QW: Quercus wutaishanica; PD: Populus davidiana; PT: Pinus tabulaeformis. Isogram generated using ArcGIS 10.0.
The spatial distribution and intraspecific spatial relations among the size classes of each tree species were analyzed by single-variable and double-variable O-ring functions (Figure 3, 4). B. platyphylla had a random distribution at almost all scales and no significant correlations were observed among size classes. Q. wutaishanica individuals were clumped at small scales (less than 7 m and 10 m to 13 m), especially at scales of less than 7 cm. The sapling of Q. wutaishanica had positive correlations with the middle-aged and adult trees at small scales. Meanwhile, Q. wutaishanica dominated the second layer in the canopy of the sample plots. P. davidiana had a random distribution at almost all scales. P. tabulaeformis was apparently clumped at small scales (less than 15 m). The sapling of P. tabulaeformis had positive correlations with the middle-aged and adult trees at small scales. Middle-sized P. tabulaeformis had positive correlations with adult individualities at scales of 5 m to 12 m.

Pattern analysis by Single variable O-ring functions of main tree species.
BP: Betula platyphylla; QW: Quercus wutaishanica; PD: Populus davidiana; PT: Pinus tabulaeformis.

Intraspecific spatial relations between different size classes (Double variable O-ring functions was used.
Populus davidiana was abandoned because the quantity of each size class was too scarce for a credible outcome) of main tree species. BP: Betula platyphylla; QW: Quercus wutaishanica; PT: Pinus tabulaeformis; S: Saplings; M: Middle-aged trees; A: Adult trees.
Interspecific correlations between main tree species
The interspecific correlation analysis (Figure 5) revealed that B. platyphylla had a negative correlation with Q. wutaishanica at a scale of less than 3 m and a positive correlation with Q. wutaishanica at a scale of 6 m to 12 m. B. platyphylla had positive correlations with P. tabulaeformis at scales of 8 m to 12 m and 30 m to 35 m. Q. wutaishanica had positive correlations with P. tabulaeformis at a scale of 16 m to 34 m. No significant correlations were observed among the other species.

Spatial distribution correlation of main tree species based on double variable O-ring function.
BP: Betula platyphylla; QW: Quercus wutaishanica; PD: Populus davidiana; PT: Pinus tabulaeformis.
Discussion
Population structure
As a pioneer tree species in the LPC, B. platyphylla achieved its predominance at the early successional stage12,13,14. In this study, the B. platyphylla population was found to have a bimodal DBH structure. Previous publications have suggested that as a shade-intolerant tree species, the Betulaceae species suffers from high seedling mortality under the canopy of mature forests and recruitment difficulties15,16. However, our results indicate that B. platyphylla saplings were abundant in the Ziwuling Mountains. The abundance of saplings was most likely the result of the low density of canopy trees (746/ha) caused by the special ecotype in the LPC. In general, seedling mortality under the canopy of Betulaceae trees increases with the aging of the forest17,18. Therefore, B. platyphylla recruitment is likely to stop with the subsequent increase in canopy density. The Gaussian model population structure of P. davidiana showed that the population had stable quantity distribution. The logistic model-based population structures of Q. wutaishanica and P. tabulaeformis were similar (with a reverse J shape) and showed a possible increasing trend. Some studies have suggested that Q. wutaishanica seedlings have high mortality at the early rooting stages but could develop vigorous populations after being planted in suitable habitats4. Q. wutaishanica is a zonal vegetation species that can reach the climate climax in arid and semi-arid regions13,19,20,21.
Spatial pattern
The seed of B. platyphylla is small and samara-shaped. These characteristics slow down the falling speed and enhance the dispersal capacity of seeds by wind. In a young secondary forest, light conditions are usually favorable for the recruitment of B. platyphylla seedlings. Such scenario can help explain the random distributions or negative correlations of B. platyphylla and P. davidiana. Distribution patterns and spatial relationships among oak recruits and resident pines have been investigated in Mediterranean environments; a random distribution of Pinus halepensis and a positive correlation between oak seedlings and pine trees have been found20. In the present study, the heavy seeds of Q. wutaishanica and P. tabulaeformis were clumped at small scales surrounding their mother trees. At large scales, Q. wutaishanica had a random distribution and P. tabulaeformis showed a discrete pattern of distribution. ZhangXiaojing et al.22 studied a secondary P. tabulaeformis forest in the Taiyue Mountains in China and reported that the spatial pattern gradually changed from clumped to random with the development of the P. tabulaeformis forest. The aforementioned findings suggest that the spatial distribution patterns of tree species depend on their biological characteristics and intraspecific relations at small scales and change with different periods (such as the change in the relationships between middle-aged trees and adult trees in this study). The spatial pattern of tree species generally changes not only with scales but also along the developmental stages23,24. The degree of the intraspecific aggregation of Q. wutaishanica andP. tabulaeformis in the present data decreased from the saplings to the adult individuals. Therefore, the increasing competition as the forest grows results in the death of members with low ability levels. This condition refers to the self-thinning rule for all probabilities. Finally, the tree species in this work showed a random or uniform spatial pattern.
Interspecific correlations
Interspecific correlations vary with forest development. At the beginning of succession, no significant correlation exists among lesser species because of adequate resource supply and minimal competition25,26. With forest development, the interspecific relationship tends to be positive or lack correlation25,26. In the Ziwuling Mountains, B. platyphylla had a negative correlation with Q. wutaishanica at small scales, thereby suggesting that the establishment of Q. wutaishanica reduced the recruitment of B. platyphylla. However, this phenomenon disappeared with the increase in scale. Local plant–plant interactions (competition/facilitation) would usually operate and be reflected at small scales (neighborhoods of plants). By contrast, habitat heterogeneity effects, such as soil, water, light and terrain, on the distribution patterns of different species could operate at large spatial scales. This study also found that P. tabulaeformis had positive correlations with B. platyphylla and Q. wutaishanica. P. tabulaeformis and Q. wutaishanica had an aggregation habit at small scales, but they could live with each other at scales of 16 m to 34 m. Hence, P. tabulaeformis and Q. wutaishanica showed a possible stable coexistence in the LPC. Bieng et al. investigated the spatial pattern of Sessile oak and Scots pine in mature mixed forests and their conclusion is similar to that of the present study. Their results showed that the productivities of both species are higher in an intimate mixture than in a patchy mixture, thereby indicating that intimate mixtures in mature oak–pine stands are favorable in the optimization of the productivities of tree species21.
Conclusion
B. platyphylla shows a random spatial pattern in the early successional stages. The establishment of Q. wutaishanica and P. tabulaeformis can interfere with the recruitment of B. platyphylla (as evidenced by their local negative spatial relationship) and forest structure can be increasingly mixed. P. davidiana is a transitional species in the LPC3,12,27. Consequently, B. platyphylla can decrease in abundance and be replaced by Q. wutaishanica and P. tabulaeformis. Furthermore, Q. wutaishanica can have a random spatial pattern and be dominant while P. tabulaeformis can live in patches. Although the B. platyphylla forest in the Ziwuling Mountains is advantageous at present, it is expected to become endangered in the long term. Artificial measures must therefore be considered to improve its regeneration.
Methods
Study area and sample selection
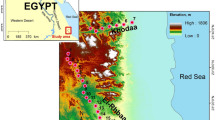
This study was conducted in the Ziwuling Mountains (107°30′ E to 109°40′ E, 33°50′ N to 36°50′ N, China) in the middle area of the LPC (Figure 6 A). The Ziwuling Mountains are the largest in the LPC, covering an area of 2.3 × 105 km2 with an attitude range of 1300 m to 1700 m and a maximum relative elevation of 400 m. The climate in the Ziwuling Mountains belongs to a transition band of temperate semi-arid and temperate sub-humid. The mean annual temperature ranges from 7°C to 8°C, with a maximum of 36°C in summer and a minimum of −27°C in winter. The mean annual relative humidity is 63% to 68% and the mean annual precipitation is 500 mm to 620 mm, with the maximum precipitation occurring during summer months. The vegetation in the area includes forest steppes and semi-arid grasslands. The thickness of loess in this area is more than 50 m.

Location of the study area in the Ziwuling Mountains of China.
Maps generated using ArcGIS 10.0.
The sample plots is located at the northeast side of the Ziwuling Mountains, which is an important habitat of B. platyphylla in the LPC (Figures 6 B and C). According to historical records28, the sample area was a lush region during the periods of the Xia, Shang and Western Zhou dynasties (2070 BC to 771 BC). The destruction of the forest began during the Warring States period (358 BC). In the next 2300 years, people migrated from the Guanzhong Plain to the frontiers (sample area today) for reclamation and frontier defense and the forests were severely damaged. Since 1966, wars and famines have depopulated this region and forests have begun to recover29,30.
Field sampling
After reconnaissance, a large plot (100 m × 100 m) and four small plots (25 m × 25 m) with light disturbance and low landscape fragmentation were selected in the typical B. platyphylla habitat in the Ziwuling Mountains (Figure 6 C). The large plot was used for spatial pattern analysis and the data of the other four small plots were mixed with those of the large plot for species and DBH counting. Based on the data from a large plot of typical forest, our study provided informational details for a better understanding of natural disturbance-restoration process in arid regions. However, it is defective in our field design that the plot selected is single time and lack of ideal replications due to the severe fragmentation and disturbance of stands. We will try our best to make improvements in future research. Within the plots, all trees with DBH (1.3 m above ground level) greater than 2.0 cm or with height taller than 2 m were measured. The names, diameters, heights, clear bole heights, crown diameters and viabilities (levels 1, 2 and 3 are defined as dying, weak and strong, respectively) of the trees were noted and the coordinates of trees were marked in coordinate papers after dividing the large plot into cells (5 m × 5 m).
Data analyses
On the basis of the age dependence of tree diameter, we used diameter classes to explain the age structures of the main tree species31,32. Each tree species was classified into 17 classes and 3 sizes according to diameter value (Class 1: 2 cm to 3.99 cm; 2 cm were added gradually from Classes 2 to 16; Class 17: >36 cm; sapling: <10 cm; middle-aged trees: 10 ≤ DBH < 20; adult trees: ≥20). Gaussian and logistic models were used for data testing and curve fitting to infer population dynamics. A polynomial fit was used to describe the data that could not be analyzed with Gaussian and logistics models. The spatial distribution patterns, intraspecific spatial relations with different sizes and interspecific correlations of the main tree species were also analyzed at multiple scales by Kriging interpolation and single-variable and double-variable O-ring functions. We used O-ring functions instead of Ripley's K functions to extract information from rings (Ripley's K functions always collect information from circles; thus, the results of Ripley's K functions at large scales repeat the information at small scales) and to correct the effects of scale accumulation from the former10,33. For the single-variable O-ring functions, the heterogeneous Poisson process was used when species distributions were clearly patchy; otherwise, complete spatial randomness was used. For the double-variable O-ring functions, an antecedent condition null model was selected in consideration of the asymmetric competition among different species. The Monte-Carlo simulation was repeated 99 times for a 95% confidence interval (the region between two dotted lines) in each process. Points higher than the confidence interval indicated clumped distributions or positive spatial correlations; points lower than the confidence interval indicated segregated or overdispersed spatial distributions or a negative spatial correlation; points within the confidence interval suggested random distributions (no significant correlation).
Software
Data analysis was completed by Programita2008 (Thorsten Wiegand 2008) and Origin 8.5 (OriginLab Corporation 2011). Figures were plotted by Arcgis10.0 (ESRI Inc. 2014) and Origin 8.5.
References
Peng, K. S. Process of control and research on soil and water loss of loess plateau the loess plateau area water and soil loss characteristics, management stage and thinking research. J. Capital Norm. Univ. (Natural Sci. Ed.). 34, 82–90 (2013).
Shao, S. A. et al. Soil quality degradation processes along a deforestation chronosequence in the Ziwuling area, China. Catena 75, 248–256 (2008).
Qin, J. & Shang, G. Z. P. Physiological-ecological effects of Populus davidiana-Quercus liaotungensis mixed forest in Ziwuling forest area. Chin. J. Appl. Ecol. 17, 927–976 (2006).
Guo, H. & Wang, X. A. Population structure analysis of Pinus tabulaeformis in Malan forest region on Ziwuling Mountain. J. Shaanxi Norm. Univ. (Natural Sci. Ed.). 40, 67–71 (2012).
Li, Y. Y., Zheng, J. Y. & Shao, M. A. Comparison of the Attributes of natural forests and plantations in Ziwuling Mountain. Acta Botanica Boreali-Occidentalia Sinica. 12, 2447–2456 (2005).
Wang, W. T. et al. Study on the characteristic of ecoclimate effect in forest zone of Ziwuling on Loess Plateau. Pratacultural Science. 4, 6–11 (2009).
Zhao, C. X. Land resources of loess plateau area. (Science Press china, Beijing, 1991).
Zou, H. Y., Liu, G. B. & Wang, H. S. The vegetation development in north Ziwuling forest region in last fifty years. Acta Botanica Boreali-Occidentalia Sinica. 1, 1–8 (2002).
Li, S. W. & Liu, L. P. Studies on types and community characters of forest and thicket vegetations in ZiwuMountain. Acta Botanica Boreali-Occidentalia Sinica. 2, 275–280 (2004).
Guo, Y. X. et al. Species composition and point pattern analysis of s tanding trees in secondary Betula albosinensis forest in Xiaolongshan of west Qinling Mountains. Chin. J. Appl. Ecol. 10, 2574–2580 (2011).
Condit, R., Ashton, P. S., Baker, P. et al. Spatial patterns in the distribution of tropical tree species. Science 288, 1414-1418 (2000).
Shao, F. L. et al. Spatial distribution pattern of Populus davidiana and Betula platyphylla populations in a typical natural secondary forest in rocky Mountain area of Northern Hebei. Scientia Silvae Sinicae. 6, 12–17 (2012).
Qin, J., Liu, Y. & Shang, G. Z. P. Photosynthetic physiological and ecological characteristics of Betula platyphylla-Quercus liaotungensis mixed forests in Ziwuling forest area. Acta Botanica Boreali-Occidentalia Sinica. 11, 2331–2337 (2006).
Su, D. K. et al. Quantitative characteristics of a secondary Betula platyphylla forest in Changbai Mountains. J. Northeast Forestry Univ. 10, 1–4 (2009).
Yasutomo, H. et al. Photosynthetic response of early and late leaves of white birch (Betula platyphylla var. japonica) grown under free-air ozone exposure. Environ Pollut. 182, 242–247 (2013).
Keyser, T. L. & Zarnoch, S. J. Stump sprout dynamics in response to reductions in stand density for nine upland hardwood species in the southern Appalachian Mountains. Forest Ecol. Manag. 319, 29–35 (2014).
Wang, S. L. The possibility analysis of natural poplar and brich forest recovery to deciduous Korean Pine forest through the method of life table in Xiaoxingan Mountains. Forestry Sci. Technol. 18, 1–4 (1993).
Huang, G. et al. Life table and survival analysis of Betula microphylla in the southern margin of junggar basin. Jiangsu Agric. Sci. 40, 310–313 (2012).
Lu, Y. C., Zhang, W. H. & Lu, Y. C. Effects of different management practices on population structure and dynamics of Quercus liaotungensis. Acta Botanica Boreali-Occidentalia Sinica. 7, 1407–1413 (2006).
Lookingbill, T. R. & Zavala, M. A. Spatial pattern of Quercus ilex and Quercus pubescence recruitment in Pinus halepensis dominated woodlands. J. Veg Sci. 11, 607–612 (2000).
Bieng, M. A. N. et al. Spatial pattern of trees influences species productivity in a mature oak–pine mixed forest. Eur. J. Forest Res. 132, 841–850 (2013).
Zhang, X. J. et al. Spatial pattern of the trees in a natural Pinus tabulaeformis forest in Taiyue Mountain. Acta Ecologica Sinica. 30, 4821–4827 (2010).
Wang, W. et al. Population structure and dynamics of Quercus liaotungensis in two broad-leaved deciduous forests in Dongling Mountain, Northern China. Acta Bot Sin. 41, 425–432 (1999).
Yi, L. T. et al. Spatial distribution patterns of Quercus liaotungensis population in Lingkong Mountains. Acta Ecologica Sinica. 28, 3254–3261 (2008).
Getzin, S. et al. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography 29, 671–682 (2006).
Barot, S., Gignoux, J. & Menaut, J. C. Demography of a savanna palm tree: Predictions from comprehensive spatial pattern analyses. Ecology 80, 1987–2005 (1999).
Jin, H. et al. Point pattern analysis of secondary poplar-birch forest in the mountainous regions of northern Hebei. J. Agric. Univ. Hebei. 3, 63–68 (2012).
Wang, Z. Y. & Lou, S. Y. Fu Xian Zhi. (People's press of Shaanxi, Xi'an, 1994).
Fan, W. Yi., Wang, X. A. & Guo, H. C. Analysis of plant community successional series in the Ziwuling area on the Loess Plateau. Acta Ecologica Sinica. 26, 706–714 (2006).
Li, Y. Y. & Shao, M. A. The change of plant diversity during natural recovery process of vegetation in Ziwuling area. Acta Ecologica Sinica. 24, 252–260 (2004).
Han, L. et al. Size-class structure and distribution pattern of Populus euphratica Oliv. in different habitats. J. Beijing Forestry Univ. 1, 7–12 (2010).
Kang, D. et al. Diameter-class structure and understory diversity of Pinus tabulaeformis plantations in Xiaolong Mountain. J. Northwest Forestry Univ. 27, 54–59 (2012).
You, H. Z. et al. The latest method of pattern analysis-Spatial point pattern analysis. J. Sichuan Forestry Sci. Technol. 6, 106–110 (2009).
Acknowledgements
We are grateful for the financial support from the Special Fund for forest-scientific Research in the Public Interest (201304312) and National Natural Science Foundation of China (41301601).
Author information
Authors and Affiliations
Contributions
D.K. and G.H.Y. conceived and designed the experiments. D.K., C.J.R. and F.Z.Z. carried out the experiments. D.K. and Y.X.G. wrote the paper. Professor Y.Z.F. and X.H.H. worked on text and language correction. All the authors discussed the results and commented on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Kang, D., Guo, Y., Ren, C. et al. Population Structure and Spatial Pattern of Main Tree Species in Secondary Betula platyphylla Forest in Ziwuling Mountains, China. Sci Rep 4, 6873 (2014). https://doi.org/10.1038/srep06873
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06873
This article is cited by
-
Tree mortality and regeneration of Euphrates poplar riparian forests along the Tarim River, Northwest China
Forest Ecosystems (2021)
-
Spatial point-pattern analysis as a powerful tool in identifying pattern-process relationships in plant ecology: an updated review
Ecological Processes (2021)
-
Vitality variation and population structure of a riparian forest in the lower reaches of the Tarim River, NW China
Journal of Forestry Research (2018)
-
Species-specific spatial structure, species coexistence and mortality pattern in natural, uneven-aged Scots pine (Pinus sylvestris L.)-dominated forest
European Journal of Forest Research (2018)
-
Effect of competition on spatial patterns of oak forests on the Chinese Loess Plateau
Journal of Arid Land (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
