
Abstract
Earthquakes are among the most destructive natural events. The 6 April 2009, 6.3-Mw earthquake in L'Aquila (Italy) markedly altered the karstic Gran Sasso Aquifer (GSA) hydrogeology and geochemistry. The GSA groundwater invertebrate community is mainly comprised of small-bodied, colourless, blind microcrustaceans. We compared abiotic and biotic data from two pre-earthquake and one post-earthquake complete but non-contiguous hydrological years to investigate the effects of the 2009 earthquake on the dominant copepod component of the obligate groundwater fauna. Our results suggest that the massive earthquake-induced aquifer strain biotriggered a flushing of groundwater fauna, with a dramatic decrease in subterranean species abundance. Population turnover rates appeared to have crashed, no longer replenishing the long-standing communities from aquifer fractures and the aquifer became almost totally deprived of animal life. Groundwater communities are notorious for their low resilience. Therefore, any major disturbance that negatively impacts survival or reproduction may lead to local extinction of species, most of them being the only survivors of phylogenetic lineages extinct at the Earth surface. Given the ecological key role played by the subterranean fauna as decomposers of organic matter and “ecosystem engineers”, we urge more detailed, long-term studies on the effect of major disturbances to groundwater ecosystems.
Similar content being viewed by others

Introduction
Though groundwater is predominantly viewed as a life-devoid water reservoir, there is a unique, stable animal community which lives in subterranean waters1,2,3. Life at extremes? Not for groundwater animals, which have developed adaptive traits as long-term groundwater dwellers3. They are small-sized, colourless, blind, with low metabolic rate and fecundity, features which allow survival in total darkness, lack of photosynthesis, low food availability and strict dependence on the organic matter entering groundwater from the surface4,5. Crustaceans dominate in groundwater communities2 and most of them are minute copepods6 (Fig. 1a), followed by ostracods7 and larger-bodied amphipods6.

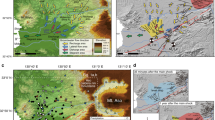
Diagram of the response of groundwater biodiversity to changes of the Gran Sasso aquifer (central Italy) hydrodynamics before and after the earthquake of 6 April 2009.
SEM micrographs of representative groundwater crustacean copepods (white bar = 200 μm) (a), schematic representation of pre-seismic (b, e), co-seismic (c, f) and post-seismic (d, g) groundwater flow from surface recharge areas through the aquifer (main conduit, lateral chambers and fracture network) to the spring discharge area, with obligate groundwater microcrustacean responses. Bottom panels report the discharge of the Tirino Springs (TS) during the March–June periods for each sampling year; mean annual discharges ± standard deviations also are reported.
More than 90% of the groundwater fauna is composed of local endemics, groundwater-exclusive taxa (from species through class) that significantly expand the phylogenetic global animal biodiversity5,6,8,9. Despite its uniqueness and a timid increase in presence within the scientific literature on inland waters, the groundwater fauna remains poorly investigated and the least known component in global biodiversity assessments8,10 being hidden, invisible and often unapproachable. Several groundwater taxa are living fossils, the only remnants of very ancient evolutionary lineages disappeared from the Earth surface, but preserved underground and at high risk of extinction5,8,11. Groundwater fauna importance in the overall freshwater biodiversity goes beyond its uniqueness. Obligate-groundwater species (i.e. stygobites, which complete their life cycle in groundwater, being unable to reproduce and survive in surface-water habitats) play key roles in organic matter recycling12,13 and as “ecosystem engineers” because the burrowing life style of worm-like elements allows the oxygenation of the minute voids among sediment particles14. Groundwater invertebrates may also serve as early warning systems in monitoring groundwater quality and quantity, being sensitive to groundwater contamination15,16,17 and withdrawal18,19, because they respond to such disturbances as much as surface-water species do. However, a knowledge vacuum still surrounds the response of groundwater fauna to both human-induced16 and natural drivers, earthquakes holding the primacy among the latter.
Earthquakes are among the most destructive natural events20. Their effects on underground hydrology are well known and are associated with the potential loss of groundwater supply for human use21. Hydrological responses depend on several factors, such as earthquake magnitude, distance from the epicentre and local geological conditions21,22,23,24. The mainshock of the L'Aquila earthquake on 6 April 2009 (Mw 6.3) dramatically changed the groundwater flow of the Gran Sasso Aquifer (GSA)25,26,27,28,29 and consequently of the Tirino River valley, where most of total aquifer discharge is located (Fig. 2). A strong base-flow component and the presence of no-flow limits defined by aquitards (constant head conditions) determine a steady water table at the Tirino Springs (TS), historically characterised by limited seasonal variations in springtime recharge, summer peak and autumn/winter shortage25,26,27. Short-term earthquake effects on GSA hydrology were determined by a pore pressure increase due to aquifer deformation25. This short-term process triggered a medium-term response in the fractured aquifer: an earthquake-induced increase in bulk hydraulic conductivity in the recharge area, near the earthquake-ruptured fault zone, attributable to fracture clearing and/or microcrack formations25, contributed to the anomalous rising of the water table and flow rate in discharge zones (Fig. 2). The otherwise stable water table at TS had progressively risen since the post-mainshock week to up to one metre (equivalent to 10× the historical data) in a month. Such an increase in water table levels could not be attributed to rainfall or snowmelt25,27,29. The water table in December 2012 was still at least 1 m higher than in the two years after the mainshock. A partial return toward pre-earthquake discharge levels started only in 2013 (Supplementary Fig. 1). The increased post-seismic discharge at TS was evident already in May 2009, reaching out-of-range values (≥30% of the previous 15 years) until the end of 2012.

Study area.
Location of the Gran Sasso Aquifer (GSA), its main recharge and discharge areas, including the Tirino Springs (TS) where sampling was carried out and the epicentre of the earthquake of 6 April 2009 that hit L'Aquila and surroundings. The two red dots represent the main springs (Presciano and Capodacqua) within the TS system (software program CorelDRAW 11 (version 11.633), 2002 Corel Corporation).
A direct response of subterranean biodiversity to earthquakes could be expected, due to the tight relationship between ground shaking, aquifer strain, fracturing and fracture clearing and habitat changes for groundwater animals, but no earthquake-related biological information in groundwater is available so far. What can groundwater fauna witness about the effects of earthquakes? How do they react to catastrophic events that dramatically rearrange the habitat where they spend their entire life cycle? Are they sensitive to earthquake-triggered hydrologic changes in groundwater? Where no other entrance to groundwater is available, answering these questions requires entering into the aquifers through “open windows”30,31, such as springs31 or other natural (caves) or artificial underground openings (boreholes)32. Springs may serve as a window to the underground, as they are considered “readily accessible outcrops of the planet's extensive groundwater resources”31.
We monitored microcrustacean copepod (body size at the μm scale) species richness and abundance along with physicochemical features at the TS, one of the main GSA discharge sites, before and after the earthquake to assess whether groundwater communities responded to hydrological changes triggered by the earthquake. Copepods were used as the target biological variable being by far the most abundant and species-rich groundwater group (~80% by individual-based density). The remainder of the meiofauna33 collected at TS included nematods (~14%), oligochaetes (~3%), ostracods (<1%), halacarid mites (<1%), gastropods (<1%), together with a small fraction of macroinvertebrates, such as amphipods and chironomid larvae (~3%).
Results
Changes in groundwater environmental descriptors
Univariate nonparametric tests returned significant 1997–2005-vs.-2012 differences for most of the environmental parameters analysed. Post-seismic groundwater pH was significantly lower and Ca2+ and dissolved oxygen (DO) concentrations were significantly higher than pre-seismic levels (Fig. 3). Electrical conductivity increased in September 2009 (up to 580 μS cm−1)25, but returned rapidly to pre-earthquake values (Fig. 3). Very fine sand and particulate organic matter (POM) significantly increased in 2012 at the TS outlets (Fig. 3).

Selected abiotic and biological variables recorded at the Tirino Springs before (1997, 2005) and after the 2009 earthquake (2012).
Boxplot representations of median (thick black line), upper and lower quartiles (box), 1.5× interquartile range (whiskers) and outliers (circles) for selected abiotic (green/blue) and biological (yellow/orange) variables at TS in the three sampling years. Post-seismic values in blue or orange. Different letters within each panel represent significantly different data distributions [Nemenyi comparisons after significant Friedman tests (Q, P; df = 2 for each)].
Water temperature at the TS sampling sites remained roughly constant throughout the sampling period, with median values at around 11°C (Supplementary Fig. 2). Post-seismic nitrate concentrations were significantly lower than pre-seismic values (Supplementary Fig. 2). Sulphate concentrations in 2012 returned to 1997 values after a transient peak in 2005 (Supplementary Fig. 2). Except for very fine sand (Fig. 3), none of the grain size classes sampled at TS changed significantly across sampling years, though the finest classes (medium and fine sand and silt and clay) tended to increase after the earthquake, albeit qualitatively (Supplementary Fig. 2). The raw abiotic data set is in Supplementary Table 1.
Effects on groundwater biodiversity
Total groundwater copepod abundance declined significantly after the earthquake (Fig. 3). Though statistical separation remained incomplete, groundwater species richness also exhibited a post-seismic negative trend (Fig. 3). Community patterns for non-obligate groundwater copepods (i.e., species which live in spring habitats because cold stenothermic or generalists and species originally coming from surface water in recharge areas and “leaked” underground) did not change significantly after the earthquake (Fig. 3). Though both non-obligate and obligate groundwater species were found at 26 out of the 32 sampling sites in 2012, such numbers represented a decrease for the stygofauna and an increase for non-obligate groundwater copepods (Table 1). The response at species level was varied for surface-water taxa, with six (~46%) species decreasing and five (~38%) species increasing in site-specific occurrence in 2012, while seven of the nine obligate groundwater species (including all the three most common pre-earthquake species) had decreased significantly in post-earthquake site-specific occurrence (Table 1).
Abundances of the 22 copepod species that we found at the 32 sampling sites declined dramatically after the earthquake (Supplementary Table 2). Pre- vs. post-earthquake differences in species abundance were highly significant (Supplementary Table 3; pairwise tests after significant one-way PERMANOVA: 1997 vs. 2012: P = 0.0009; 2005 vs. 2012: P = 0.0015), while differences between the two pre-seismic samplings remained non-significant (1997 vs. 2005: P = 0.5131). Obligate groundwater species mostly accounted for the dramatic post-earthquake decrease in faunal abundance (pairwise tests after significant one-way PERMANOVA: 1997 vs. 2012: P = 0.0007; 2005 vs. 2012: P = 0.0012; 1997 vs. 2005: P = 0.7450), with the contribution from non-obligate groundwater species remaining non-significant (one-way PERMANOVA: P = 0.1379) (Supplementary Table 3).
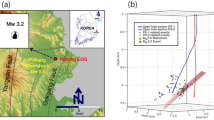
The groundwater species Nitocrella pescei, Diacyclops paolae, Parastenocaris lorenzae and Elaphoidella mabelae cumulatively provided 90% of the discrimination between pre- vs. post-earthquake assemblages (Supplementary Table 4). N. pescei contributed the most (41.7%) to the pre- vs. -post dissimilarity by declining by ~2.5× after the earthquake (Fig. 4, Supplementary Table 4). The other three most common species contributed further to pre- vs. post-seismic dissimilarity to a cumulative ~50%, though D. paolae and P. lorenzae decreased in abundance by only 0.5× and E. mabelae had slightly increased by 2012 (Table 1, Supplementary Table 4, Fig. 4). The vast majority of the remaining groundwater species became rare to very rare in 2012 (Table 1, Supplementary Table 2).

Pre- and post-seismic abundance patterns of dominant groundwater copepods at the Tirino Springs.
Nonmetric multidimensional scaling ordination of pre- and post- earthquake abundance of the obligate groundwater copepod species that accounted for ~90% of abundance changes; 2D stress = 0.16. Pre-earthquake replicate-specific abundance data in light blue and post-earthquake abundances in red; circle size is proportional to abundance).
Discussion
The mainshock of the L'Aquila earthquake of 6 April 2009 markedly changed the Gran Sasso groundwater flow and consequently the discharge at the main spring sites. Medium-term local effects included an anomalous increase in TS discharge through 201225,26,27,28,29 (Supplementary Fig. 1). The TS discharge gradually returned toward pre-seismic values only in summer 2013 (Supplementary Fig. 1), consistent with model predictions27,29.
Pre- vs. post-earthquake changes in groundwater physico-chemistry were observed at the TS main outlets. Post-seismic water pH (Fig. 3) decreased as a reflection of an increase of deeply-derived CO2 along the main faults during the mainshock34. The post-seismic decrease in pH was accompanied by a significant increase in Ca2+ concentration from calcite dissolution25 (Fig. 3). Electrical conductivity increased in September 2009 as a result of the clearing of previously obstructed fractures from which high residence-time ion-enriched water flushed out after crustal stress and ground shaking (Fig. 3). Concentrations of very fine sand and POM increased at the main TS outlets (Fig. 1b–d, Fig. 3) due to the massive dewatering of the conductive systems which emptied the annexed fractures in the aquifer. Dissolved oxygen (DO) concentration increased significantly after the earthquake (Fig. 3), as expected for the increase in fast groundwater flowpaths in the aquifer due to a post-seismic higher hydraulic conductivity.
The decrease in post-seismic nitrate concentrations (Supplementary Fig. 2) was most likely due to a markedly lower human presence in the catchment area following the post-earthquake temporary evacuation of towns and farms. Higher sulphate concentrations in 2005 (Supplementary Fig. 2) were related to the dual groundwater flow system feeding the TS35. The seasonal contribution from local heavy rainfall led to an above-average recharge rate in 2005 and to an enrichment in SO42− due to a more intense interaction with the Triassic evaporite bedrocks outcropping along the local and most superficial groundwater flowpath35. Conversely, the highest post-seismic groundwater contribution was from the deepest, base-flow fractured aquifer, described by large karstic conduits and the annex capacitive subsystem25,26,27,28. Absence of temporal changes for most such variables and the overall low values for nitrates and sulphates (Supplementary Fig. 2) suggest a limited (if any) influence of such variables on the observed changes in aquifer-dwelling biota.
Stygobiotic copepods responded negatively to changes in the groundwater hydrodynamics triggered by the earthquake (Table 1, Figs. 3, 4, Supplementary Table 3). However, the relative contributions of the single-event massive mainshock (from which the stygofauna had yet to recover as of late 2012) or from the post-mainshock diminished drift (due to the overall initial crash in stygobiotic abundance) remain unknown. Interestingly enough, our earthquake-targeting investigation has led to a noteworthy corollary finding: though most species within the harpacticoid genus Elaphoidella are true stygobites36, the slight but significant post-earthquake increase in E. mabelae presence (Table 1, Fig. 4) supports the view of an intermediate stygo- and crenobiotic ecology of this species37.
Though a few species can cope with the challenging fast-flowing conduit system of karstic aquifers, the dendritic fracture network preferentially allows the establishment of permanent (in ecological times) invertebrate communities38,39. The low flow rate in such fracture “chambers” (Fig. 1b–d) allows the storage of fine sediments, which form the physical substrate for burrowing animals and of POM, which constitutes the bulk of the food. Open-water swimmers also avoid massive drifting (Fig. 1b).
Based on our limited dataset of three non-contiguous sampling years and considering the relatively stable pre-seismic groundwater faunal patterns, we propose that the earthquake flushed out groundwater invertebrates, very fine sand and POM at the aquifer discharge sites (Fig. 1c). The massive earthquake-induced increase in hydraulic conductivity and the consequent aquifer dewatering (Fig. 1e–g) thus acted as what we call a biotrigger. Immediately after the mainshock, groundwater communities were stranded in springbed sediments, where they survived and maintained the pre-earthquake species richness albeit at lower abundances, a condition expected if individuals of fracture-dwelling species were massively flushed out (Table 1). Neither the post-seismic higher DO nor the higher POM availability at the TS outlets favoured the survival and especially the reproduction of groundwater species and the contribution of post-earthquake drift for groundwater species was still negligible as of 2012. Stygobite numerical losses in the aquifer fractures were not consistently or fully counterbalanced by conspecifics coming from their original habitat (i.e. small and large fractures eventually less affected by the aquifer dewatering). Progressively decreasing abundances of obligate groundwater species (i.e., stygobites) were found at the TS outlet, because the aquifer became almost totally deprived of animal life, no longer replenishing groundwater populations normally flushing out at the spring outlets (Fig. 1c, d). Abundance and species richness of non-obligate groundwater species remained unchanged (Fig. 3), suggesting that the earthquake had an impact only on aquifer-dwelling microinvertebrates. We therefore believe that the main driver of the observed crash in post-earthquake biodiversity occurred inside the aquifer.
How much time is needed for the recolonisation of the Gran Sasso aquifer by groundwater species, allowing the reestablishment of viable microcrustacean populations? We think that such a process will take a very long time, probably several decades, much longer than the return to the pre-earthquake aquifer recharge/discharge cycle and geochemistry. Groundwater communities are notorious for their low resilience3. Virtually all species have low reproductive rates, long life spans (up to 3.5 years for groundwater copepods vs. ≤1 year for surface-water relatives) and long post-embryonic development times before reaching sexual maturity4,6. The potential for replacement by other groundwater species is very low, aquifer communities being characterised by low community redundancy3 (although omnivory is claimed as a relatively frequent functional trait in the oligotrophic groundwater40, an ecological statement awaiting confirmation).
Our investigation is based on a limited dataset of three complete but non-contiguous hydrological years, so that a precise quantification of background variability (“noise”) was not possible. Though some abiotic variables were fully comparable in the two pre-earthquake years (e.g., pH, Ca2+, POM and DO in Fig. 3), others were not (electrical conductivity in Fig. 3 and SO42− in Supplementary Fig. 2). Also, some site-specific spatial variability occurred (e.g., electrical conductivity slightly higher at site 8 in Supplementary Table 1), suggesting the existence of some background variability in hydrogeology. However, all statistical analyses pointed to a pre-vs.-post-earthquake variation higher than within pre-earthquake variation (e.g., Friedman tests in Fig. 3 and all multivariate biological tests). Therefore, we believe that our results remain valid and robust as a first-look investigation into the effects of a major single-event disturbance such as an earthquake on groundwater ecosystems, but we urge more detailed, long-term studies on the topic. Despite the limited dataset, our results strongly suggest that a catastrophic natural disturbance such a major earthquake seems to negatively impact the survival or reproduction of aquifer-dwelling stygobites, with a potential detrimental effect on groundwater biodiversity.
Methods
Sampling and sample processing
The Tirino Springs (TS) were selected as the study site because a natural entrance to the aquifer (e.g., caves) is not available in the Gran Sasso karst. In such cases, sampling is carried out at spring outlets31,39,41 or through artificial boreholes32. Most of the Gran Sasso Aquifer (GSA) discharge occurs through springs at the aquifer boundary (Fig. 2) and the TS receive ~65% of the aquifer discharge42. We sampled the TS through bedrock fractures in the springbed, deep into the fractures well below the springbed itself, or in the aquitards where present, at different depths in order to collect obligate groundwater dwellers (i.e., stygobites). The high groundwater discharge from under the springbed maximised the collection of groundwater and stygobites, either drifted from the karstic discontinuities of the carbonate bedrock, or stranded in the alluvial aquitards. Stygobites depend on groundwater to complete their life cycle, as they cannot reproduce in other environments. Species were characterised as stygobites on the basis of well-established morphological, physiological and ecological features2,3,6,36. Stygobites are frequently found in spring habitats where they can temporarily survive after being drifted from their original groundwater habitats38,39,41. Surface-water species are also found along with stygobites, either flushed out from the aquifer where they enter from recharge areas, or because they live in springs, as crenobionts or surface-water habitat generalists43.
Abiotic and biological variables were determined at eight sampling sites at the TS adopting a random sampling method with four seasonal replicates per site (Jan–Feb, Mar–Apr, Jul–Aug, Sep–Oct) in three discrete years: 1997 and 2005 (before the earthquake) and 2012 (after the earthquake). Samples were collected with a Bou-Rouch pump44 and mobile pipes hammered at each sampling site. Samples were composite from 0.3, 0.7 and 1.5 m below the springbed. Since both sample size and pumping rate may affect estimates of faunal density for a range of different sediment types32,45,46, we maintained consistent pumping rates and sample volume sizes throughout the investigation. Though 5-L samples would suffice to detect spatial variation in invertebrate composition among sites with different sediment texture, sample size affecting mainly large-sized invertebrates like insect larvae46, a sample size of 20 L was adopted for density estimates, thus including abundance-related rarity47 of groundwater copepods4,6. The copepods were used as the target biological variable being by far the most abundant and species-rich groundwater group (~80% by individual-based density).
Meiofauna was extracted by filtering the 20-L samples through a hand net (mesh size = 60 μm). Faunal samples were preserved in an ethyl alcohol solution at 80%. Meiofaunal individuals were later sorted, counted, identified to species level and assigned to two ecological categories: obligate (stygobites) or non-obligate groundwater species (non-stygobites). Non-stygobites are epigean freshwater species living on the springbed, or as inbenthic in sediment interstices (e.g., to avoid predation) or are habitat generalists. Some of them are defined as crenobionts30, i.e. species which complete their life cycle in the stable and cold thermal regime of surface spring waters.
Temperature, pH, specific conductivity at 25°C and dissolved oxygen (DO) concentration were measured in the field using a multiparametric probe (ECM Multi™; Dr. Lange GmbH, Düsseldorf, Germany); calcium concentration was determined by titration with EDTA; nitrate, sulphate and phosphate concentrations were measured using a HACH DR 2000 spectrophotometer (Hach Co., Loveland, CO, U.S.A.). Particulate organic matter (POM) was quantified on 20-L samples, after removal of all fauna, oven-drying at 105°C for 24 h and weighing in anhydrous conditions. The dry-weighed samples were ashed at 540°C and re-weighed to determine POM concentration as the difference between dry and ash mass. The grain size composition of springbed sediments was measured only once per year at each spring site, as sediment transport was negligible throughout the spring system. A more intensive sampling of sediments also would have altered habitat features and hence species abundance and distribution across sites. The mass of eight granulometric classes was determined at each site on the fractional dry-sieving of desiccated samples through a stack of sieves according to the Wentworth48 scale: medium gravel (4–8 mm), fine gravel (2–4 mm), very coarse sand (1–2 mm), coarse sand (0.5–1 mm), medium sand (0.25–0.5 mm), fine sand (0.125–0.25 mm), very fine sand (0.063–0.125 mm), silt and clay (<0.063 mm).
Statistical analysis
The sampling design was fully balanced, thus maximising statistical power and reliability49,50. Coleman rarefaction curves run for biological data indicated that sampling efforts were adequate for all sampling years (Supplementary Table 5, Supplementary Fig. 3). Coleman rarefaction curves were constructed with EstimateS 951 using resampling with replacement based on 50 iterations.
Between-year comparisons for abiotic and biological variables were tested with nonparametric Friedman tests, a block-based variant of the Kruskal-Wallis test, also based on ranks and used when k > 249,50,51. Sampling sites within sampling years were used as blocks, resulting in triplet-based paired comparisons (simultaneous 1997 vs. 2005 vs. 2012 comparisons). The paired-sample, rank-based approach was chosen also to minimise the impact of the inherent between-site variability, especially for some variables (e.g., grain size composition of sediments). Sample size (n) was 32 for all variables in each year, except for grain size, for which n = 8. Friedman tests were thus fully reliable for all variables, though the outcome remained slightly liberal for the relatively low-replication grain size categories50. However, the Friedman test is considered a robust technique as long as the assumption of homoscedasticity is met49,50,52. The Friedman test was applied only to those variables with homogeneous variances, as tested with a two-tailed Cochran C test53 (P ≥ 0.369 for all abiotic variables and major biological groups; Supplementary Table 6). P values for Friedman tests were determined asymptotically. A Nemenyi paired-sample multiple comparison was performed for statistically significant Friedman tests to identify pairwise year-vs.-year trends. Partly to counterbalance the possible slight liberalism of borderline significant Friedman and/or Nemenyi results, a Bonferroni correction was applied to all tests (whose original significance threshold was set at P < 0.05), leading to a Cochran-Friedman-Nemenyi experiment-wide significance threshold of P < 0.016. Cochran, Friedman and Nemenyi tests were performed using XLSTAT (Addinsoft™ XLSTAT v. 2013.6.01, Paris, France, 2014).
Data for many individual species of copepods were heteroscedastic (significant two-tailed Cochran test: Supplementary Table 6) and could not be tested with the Friedman/Nemenyi procedure50,52. Year-vs.-year patterns in copepod species distributions were thus analysed with a test of independence on a contingency table based on frequencies of yearly occurrences at the 32 sampling sites (i.e., 32 = the species was observed at all sampling sites in that year: Table 1). Due to the overall low frequencies and patchy species distributions, the original data were coded by a constant ×100 factor52. The test of independence was based on two-tailed Pearson X2 distance (X2P) with df = (number of years − 1) × (number of species − 1) = 42. Significant departures from table-wide independence were followed by an analysis of adjusted Pearson residuals to identify observed frequencies for individual species significantly higher or lower than expected frequencies50. Significance for adjusted Pearson residual analyses was set at the same level as for the preceding Pearson X2 distance, standardised for the non-PERMANOVA investigation-wide statistical elaboration (i.e., P < 0.016). The same procedure was applied for the two major ecological copepod groups (obligate vs. non-obligate groundwater species) in a 3 years × 2 groups contingency table (Table 1). Contingency tables were analysed with Addinsoft™ XLSTAT.
A permutational multivariate analysis of variance (PERMANOVA)54 was used to test for differences in copepod abundances before (1997 and 2005) and after (2012) the earthquake. Post-hoct-tests were used to test for differences between levels within the factor. Significance threshold for PERMANOVA analyses was set at P < 0.05. Following significant PERMANOVA results, patterns of obligate groundwater copepod assemblages before and after the earthquake were examined using nonmetric multidimensional scaling (nMDS) incorporating the Bray-Curtis similarity coefficient. SIMPER analysis55 was then used to identify the obligate groundwater species that mostly accounted for pre-vs.-post-earthquake differences in abundance. In both nMDS and PERMANOVA, data were square-root transformed to weight the contribution of common and rare species and a dummy variable of 1 was added to all samples to allow the inclusion of otherwise empty samples (zero abundance). A Levene test for homogeneity of variances was carried out before PERMANOVA (Supplementary Table 3). All multivariate analyses were done using PRIMER v6 (PRIMER-E, Plymouth, UK, 2006) and R software (R Development Core Team, R Foundation for Statistical Computing, Vienna, Austria, 2008).
References
Holsinger, J. R. Troglobites: the evolution of cave-dwelling organisms. Am. Sci. 76, 147–153 (1988).
Gibert, J. & Deharveng, L. Subterranean ecosystems: a truncated functional biodiversity. Bioscience 52, 473–481 (2002).
Gibert, J., Danielopol, D. L. & Stanford, J. A. Eds. Groundwater Ecology (Academic Press, San Diego, 1994).
Dole-Olivier, M.-J., Galassi, D. M. P., Marmonier, P. & Creuzé des Chatelliers, M. The biology and ecology of lotic microcrustaceans. Freshwater Biol. 44, 63–91 (2000).
Stein, H. et al. Stygoregions – a promising approach to a bioregional classification of groundwater systems. Sci. Rep. 2, http://www.nature.com/srep/2012/120919/srep00673/pdf/srep00673.pdf (2012) Date of access: 01/07/2014.
Galassi, D. M. P., Huys, R. & Reid, J. W. Diversity, ecology and evolution of groundwater copepods. Freshwater Biol. 54, 691–708 (2009).
Smith, A. J. & Delormes, L. D. [Ostracoda] [Thorp J. H. & Covich A. P. (eds.)] [811–849] (Academic Press/Elsevier, Amsterdam, 3rd Ed., 2009).
Gibert, J. et al. Assessing and conserving groundwater biodiversity: synthesis and perspectives. Freshwater Biol. 54, 930–941 (2009).
Purvis, A., Agapow, P.-M., Gittleman, J. L. & Mace, G. M. Nonrandom extinction and the loss of evolutionary history. Science 288, 328–330 (2000).
Zagmajster, M. et al. Geographic variation in range size and beta diversity of groundwater crustaceans: insights from habitats with low thermal seasonality. Global Ecol. Biogeogr. http://onlinelibrary.wiley.com/doi/10.1111/geb.12200/pdf (2014) Date of access: 01/07/2014.
Galassi, D. M. P., Dole-Olivier, M.-J. & De Laurentiis, P. Phylogeny and biogeography of the genus Pseudectinosoma and description of P. janineae sp. n. (Crustacea, Copepoda, Ectinosomatidae). Zool. Scr. 28, 289–303 (1999).
Boulton, A. J., Fenwick, G. D., Hancock, P. J. & Harvey, M. S. Biodiversity, functional roles and ecosystem services of groundwater invertebrates. Invertebr. Syst. 22, 103–116 (2008).
Brunke, M. & Gonser, T. The ecological significance of exchange processes between rivers and groundwater. Freshwater Biol. 37, 1–33 (1997).
Dole-Olivier, M.-J. et al. Assessing invertebrate assemblages in the subsurface zone of stream sediments (0–15 cm deep) using a hyporheic sampler. Water Resour. Res. 50, 453–465, 10.1002/2012WR013207 (2014). Date of access: 01/07/2014.
Tomlinson, M., Boulton, A. J., Hancock, P. J. & Cook, P. G. Deliberate omission or unfortunate oversight: should stygofaunal surveys be included in routine groundwater monitoring programs? Hydrogeol. J. 15, 1317–1320 (2007).
Griebler, C., Malard, F. & Lefébure, T. Current developments in groundwater ecology — from biodiversity to ecosystem function and services. Curr. Opin. Biotechnol. 27, 159–167 (2014).
Korbel, K. L. & Hose, G. C. A tiered framework for assessing groundwater ecosystem health. Hydrobiologia 661, 329–349 (2011).
Di Lorenzo, T. & Galassi, D. M. P. Agricultural impact on Mediterranean alluvial aquifers: do groundwater communities respond? Fund. Appl. Limnol. 182, 271–282 (2013).
Stumpp, C. & Hose, G. C. The impact of water table drawdown and drying on subterranean aquatic fauna in in-vitro experiments. PLOS ONE 8, e78502 (2013).
Millennium Ecosystem Assessment, Ecosystems and Human Well-being: Synthesis (Island Press, Washington, DC, 2005).
Montgomery, D. R. & Manga, M. Streamflow and water well responses to earthquakes. Science 300, 2047–2049 (2003).
Chen, J. S. & Wang, C.-Y. Rising springs along the Silk Road. Geology 37, 243–246 (2009).
Wang, C. -Y. & Manga, M. Hydrologic responses to earthquakes - a general metric. Geofluids 10, 206–216 (2010).
Manga, M. & Rowland, J. C. Response of Alum Rock springs to the October 30, 2007 earthquake and implications for the origin of increased discharge after earthquakes. Geofluids 9, 237–250 (2009).
Adinolfi Falcone, R. et al. Changes on groundwater flow and hydrochemistry of the Gran Sasso carbonate aquifer after 2009 L'Aquila earthquake. Ital. J. Geosci. (Boll. Soc. Geol. It.) 131, 459–474 (2012).
Amoruso, A. et al. Impact of the 6 April 2009 L'Aquila earthquake on groundwater flow in the Gran Sasso carbonate aquifer, Central Italy. Hydrol. Process. 25, 1754–1764 (2011).
Amoruso, A., Crescentini, L., Petitta, M. & Tallini, M. Parsimonious recharge/discharge modeling in carbonate fractured aquifers: The groundwater flow in the Gran Sasso aquifer (Central Italy). J. Hydrol. 476, 136–146 (2013).
Amoruso, A., Crescentini, L., Martino, S., Petitta, M. & Tallini, M. Correlation between groundwater flow and deformation in the fractured carbonate Gran Sasso aquifer (INFN underground laboratories, central Italy). Water Resour. Res. 50, http://onlinelibrary.wiley.com/doi/10.1002/2013WR014491 (2014) Date of access: 01/07/2014.
Tallini, M. et al. Isotope hydrology and geochemical modeling: new insights into the recharge processes and water–rock interactions of a fissured carbonate aquifer (Gran Sasso, central Italy). Environ. Earth Sci. http://link.springer.com/article/10.1007%2Fs12665-014-3364-9 (2014) Date of access: 01/07/2014.
Botosaneanu L. Ed. Studies in Crenobiology: the Biology of Springs and Springbrooks. (Backhuys Publishers, Leiden, 1988).
Barquín, J. & Scarsbrook, M. Management and conservation strategies for coldwater springs. Aquatic Conserv: Mar. Freshw. Ecosyst. 18, 580–591 (2008).
Sorensen, J. P. R. et al. Using boreholes as windows into groundwater ecosystems. PLOS ONE 8, e70264 (2013).
Giere, O. Meiobenthology. The microscopic motile fauna of aquatic sediments. (Springer-Verlag Berlin, 2009).
Chiodini, G. et al. Geochemical evidence for and characterization of CO2 rich gas sources in the epicentral area of the Abruzzo 2009 earthquakes. Earth Planet. Sci. Lett. 304, 389–398 (2011).
Petitta, M., Caschetto, M., Galassi, D. M. P. & Aravena, R. Dual-flow in karst aquifers toward a steady discharge spring (Presciano, central Italy): influences on a subsurface groundwater dependent ecosystem and on changes related to post-earthquake hydrodynamics. Environ. Earth Sci. http://link.springer.com/article/10.1007%2Fs12665-014-3440-1 (2014) Date of access: 30/07/2014.
Galassi, D. M. P. Groundwater copepods: diversity patterns over ecological and evolutionary scales. Hydrobiologia 453/454, 227–253 (2001).
Galassi, D. P. & Pesce, G. L. Elaphoidella mabelae, n. sp., a crenobiont harpacticoid from Italy (Copepoda, Canthocamptidae). Crustaceana 60, 1–6 (1991).
Rouch, R., Pitzalis, A. & Descouens, A. Effets d'un pompage à gros débit sur le peuplement des Crustacés d'un aquifère karstique. Ann. Limnol. 29, 15–29 (1993).
Gibert, J., Vervier,. Ph.,. Malard, F., Laurent, R. & Reygrobellet, J.-L. [Dynamics of communities and ecology of karst ecosystems: example of three karsts in Eastern and Southern France] [Gibert J., Danielopol D. L., & Stanford J. A. (eds.)] [425–450] (Academic Press, San Diego, 1994).
Claret, C. et al. A functional classification of interstitial invertebrates: supplementing measures of biodiversity using species traits and habitat affinities. Arch. Hydrobiol. 145, 385–403 (1999).
Fiasca, B. et al. The dark side of springs: what drives small-scale spatial patterns of subsurface meiofaunal assemblages? J. Limnol. 73, 55–64.
Barbieri, M., Boschetti, T., Petitta, M. & Tallini, M. Stable isotopes (2H, 18O and 87Sr/86Sr) and hydrochemistry monitoring for groundwater hydrodynamics analysis in a karst aquifer (Gran Sasso, central Italy). Appl. Geochem. 20, 2063–2081 (2005).
Gerecke, R. et al. [Studies in crenobiology: the biology of springs and springbrooks] [167–182] [Botosaneanu L. (ed.)]. (Backhuys Publishers, LeidenNL, 1988).
Bou, C. & Rouch, R. Un nouveau champ de recherches sur la faune aquatique souterraine. C. R. Acad. Sci. 265, 369–370 (1967).
Hunt, G. W. & Stanley, E. H. An evaluation of alternative procedures using the Bou–Rouch method for sampling hyporheic invertebrates. Can. J. Fish. Aquat. Sci. 57, 1545–1550 (2000).
Boulton, A. J., Dole-Olivier, M.-J. & Marmonier, P. Effects of sample volume and taxonomic resolution on assessment of hyporheic assemblage composition sampled using a Bou-Rouch pump. Arch. Hydrobiol. 159, 327–355 (2004).
Gaston, K. J. Rarity (Springer Netherlands, 1994).
Wentworth, C. K. A scale of grade and class terms for clastic sediments. J. Geol. 30, 377–392 (1922).
Underwood, A. J. Experiments in ecology — Their logical design and interpretation using analysis of variance. (Cambridge Univ. Press, Cambridge, UK, 1997).
Quinn, G. P. & Keough, M. J. Experimental design and data analysis for biologists. (Cambridge Univ. Press, Cambridge, UK, 2002).
Colwell, R. K. EstimateS: statistical estimation of species richness and shared species from samples. Version 9. User's Guide and application published at: http://viceroy.eeb.uconn.edu/estimates (2013) Date of access: 01/07/2014.
Zar, J. H. Biostatistical analysis. 5th ed. (Prentice Hall/Pearson, Upper Saddle River, NJ, 2009).
't Lam, R. U. E. Scrutiny of variance results for outliers: Cochran's test optimized. Anal. Chim. Acta 659, 68–84 (2010).
Anderson, M. J. A new method for non-parametric multivariate analysis of variance. Austral. J. Ecol. 26, 32–46 (2001).
Clarke, K. R. Non-parametric multivariate analysis of changes in community structure. Austral. J. Ecol. 18, 117–143 (1993).
Acknowledgements
We thank the technical staff of the Stygobiology Research Group of the University of L'Aquila (Italy) for assistance in field work. The project was funded by the European Community (LIFE12 BIO/IT/000231 AQUALIFE) and the Gran Sasso-Laga National Park (Italy).
Author information
Authors and Affiliations
Contributions
D.M.P.G. had the idea; B.F. and A.D.C. designed the sampling method and analysed the samples; D.M.P. and B.F. identified the species; P.L., B.F. and T.D.L. performed the statistical analyses; P.L. and P.D.C created the figures; M.P. wrote the hydrogeological setting of the study area. All authors discussed the results and contributed to writing the paper.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Galassi, D., Lombardo, P., Fiasca, B. et al. Earthquakes trigger the loss of groundwater biodiversity. Sci Rep 4, 6273 (2014). https://doi.org/10.1038/srep06273
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06273
This article is cited by
-
Distribution, threats and protection of selected karst groundwater-dependent ecosystems in the Mediterranean region
Hydrogeology Journal (2023)
-
Large extensional earthquakes push-up terrific amount of fluids
Scientific Reports (2022)
-
The impact of nitrate on the groundwater assemblages of European unconsolidated aquifers is likely less severe than expected
Environmental Science and Pollution Research (2021)
-
Shifts in zooplankton communities in high-mountain lakes induced by singular events (fish stocking, earthquakes): evidence from a 20-year survey in Slovenia (Central Europe)
Aquatic Ecology (2021)
-
Spatial distribution of stygobitic crustacean harpacticoids at the boundaries of groundwater habitat types in Europe
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
