
Abstract
Coral populations are in decline due to environmental changes and biological attacks by predators and infectious diseases. Here, we report a localized bloom of the benthic filamentous cyanobacterium Moorea bouillonii (formerly Lyngbya bouillonii) observed exclusively on the gorgonian (sea fan) coral Annella reticulata at around 20 m depth in Japan. The degree of infection has reached 26% among different sizes of Annella colonies. Thick and continuous growth of Moorea may be sustained partly by symbiotic alpheid shrimp, which affix Moorea filaments to gorgonian corals for use as food and shelter. Most filaments get entangled on the coral colony, some penetrate into the stem of the coral with a swollen end like a root hair, which appears to function as an anchor in Annella. In addition to the cyanobacterium–shrimp interaction, the new trait of anchoring by the cyanobacterium into gorgonian coral may contribute to persistence of this bloom.
Similar content being viewed by others

Introduction
Populations of reef-building coral have been in decline due to biological and physiological changes, such as predation by crown-of-thorns starfish and other species, coral bleaching driven by increasing water temperature, a variety of diseases and ocean acidification1,2,3,4,5. It has been predicted that, in regions with coral reefs, a phase shift from coral to algae will occur due to climate change and anthropogenic activities1,3. Algal (including cyanobacteria) blooms, which are usually caused by excess nutrient input into reefs from terrestrial systems4, suppress both coral growth and coral recruitment5. Algae–coral interactions have been of increasing interest because algae can produce poisonous chemicals and vector coral diseases6,7,8.
Harmful cyanobacterial blooms have received increasing attention because they pose a serious threat to the use and sustainability of freshwater and marine resources9,10. Most cyanobacterial blooms in coral reefs are caused by inputs of nutrients; examples include guano containing phosphorus at the Great Barrier Reef, Australia11 and nitrogen inputs into watersheds of Florida, USA4,12,13. In addition to nutrient-induced cyanobacterial blooms, some cyanobacteria cause coral diseases. Black band disease (BBD) is a well-known coral disease, caused by cyanobacteria and a complex consortium of other bacteria14. Soft corals (octocorals), as well as hard corals, have experienced higher rates of infectious diseases in recent years. The fungus Aspergillus sydowii causes the disease aspergillosis and exclusively kills the common Caribbean octocoral Gorgonia ventalina15,16,17.
Moorea bouillonii is a common benthic filamentous cyanobacterium distributed widely such as Papua New Guinea, Guam, Palau, Palmyra atoll etc.18, French Polynesia19 and Japan20. We report on a localized cyanobacterial bloom in a colony of sea fan coral, Annella reticulata (Anthozoa, Alcyonacea, Gorgoniidae), caused by a common benthic cyanobacterium, M. bouillonii, which proliferates in the water near Okinawa, Japan.
Results
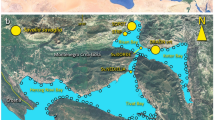
This study was conducted at Sakubaru-reef, Aka-jima, Okinawa, Japan (26°10′37″ N, 127°16′ 27″E), which is approximately 1.5 km from the residential area of Aka-jima (Fig. 1). This is a deep (>20 m), clear-water environment where gorgonian corals (sea fans) are dominant and scleractinian corals also occur along the reef slope. The gorgonian coral Annella reticulata (Fig. 2A) forms a dense population approximately 50 m wide by 100 m long, at 10–25 m depth (A. Kishi, pers. com.). Attachment of filamentous cyanobacteria to gorgonian coral was first observed about 10 years ago at 20 m depth and has gradually increased in both number and in extent since then (H. Matayoshi, personal observation). Cyanobacterial overgrowth was observed exclusively on the colony surface of the gorgonian coral and was not found on other substratum or sessile organisms including scleractinian corals (Fig. 2B,D).

Map showing the study site.
Bloom of the cyanobacterium Moorea bouillonii on the gorgonian coral Annella reticulata was observed at Sakubaru reef (arrow) in Akajima Island (A). Maps were downloaded from The Geospatial Information Authority of Japan.

(A), The colonies of gorgonian coral Annella reticulata. (B), Overgrown by the cyanobacterium Moorea bouillonii. (C), Enlarged view of M. bouillonii. (D), Close-up of the coral branches entangled by Moorea. (E), After removal of M. bouillonii filaments, dead branch (white) part and collapsed branch leaving only a central axis (arrow) are shown.
The cyanobacterial alga was identified as Moorea bouillonii (Basionym: Lyngbya bouillonii Hoffmann & Demoulin, 1991)18,22,23 by its color, cell size (Fig. 2C) and 16S ribosomal RNA sequence. Partial 16S ribosomal RNA sequence of it (about 680 bp, Accession No. AB922817) supported that the cyanobacteria belonged to the genus Moorea. The gorgonian coral was identified by its colony structure and sclerites as Annella reticulata (Ellis and Solandar, 1786).
Percentage of infected colonies of M. bouillonii on A. reticulata reached 26% and some algal cover was found on every size class of coral colony (Fig. 3). A small amount of other algal species (filamentous green or attached diatoms) was observed, but M.bouillonii was the most abundant and entangled on branches of Annella. The sewing (tube-forming) shrimp, Alpheus frontalis H. Milne Edwards 1958, was identified by the presence of tubular cyanobacteria, but quantitative measurement was not performed because most shrimp escaped from the cyanobacterial tubes during collection. Other small organisms found in the cyanobacterial mat, which were considered to be secondarily attached, included foraminiferas, nematodes, copepods, gastropods and tunicates etc. Nutrient concentrations measured from the sea surface to 25 m in depth were 0.17 µmol for NH4, 0.04 µmol for NO2, 0.99 µmol for NO3 and 0.09 µmol for PO4.

Percentage of infection of the cyanobacterium Moorea bouillonii on the gorgonian coral Annella reticulata at a depth of 18 ± 1 m (n = 91).
Coral branches overgrown by Moorea mats ultimately die, which results in collapse of the branch (Fig. 2E) followed by gradual detachment of outer sclerites and then the loss of successively longer sclerites that form the inner axis. Most Moorea bouillonii filaments were entangled on coral colonies, although some were loosely attached and lying on the branch surface. Some filaments penetrated directly into the coral branch and reached the outermost region of the central axis (Fig. 4A). The terminal end of the boring filament was swollen like a hair root and consisted of a multilayered sheath (Fig. 4B) which functions as an anchor.

(A), Penetration by the cyanobacterium Moorea bouillonii into host coral Annella reticulata. (B), Terminal end of M. bouillonii showing swallen structure composed of multi-layer of sheath.
Discussion
Seaweeds negatively impact corals via multiple mechanisms such as shading, abrasion, vectoring of coral diseases and release of metabolites8,24. Cyanobacterial blooms in coral reefs are due to excessive anthropogenic nutrient loading and have been reported from Florida, USA4,13, Guam25 and Queensland, Australia11,26. The ambient nutrient concentrations measured at this study site (0.99 µmol for NO3 and 0.09 µmol for PO4) were slightly below the levels previously reported for sustaining macroalgal blooms (1.0 µmol for NO3 and 0.1 µmol for PO427 or cyanobacterial growth in enrichment experiments26,28,29. On the other hand, it is widely accepted that reefs are not limited to low-nutrient areas1,27,30. Thus, to accurately address algal growth, we must consider nutrients, producers (algae) and consumers (herbivores, predators).
Herbivores consume algae, affecting algae–coral interactions. In a marine protected area in Fiji, the amount of macroalgae is better controlled than in an adjacent fished reef7. In general, however, filamentous cyanobacteria and gorgonian corals are generally not preferred food for predators such as benthivorous fish31,32,33. Furthermore, L. majuscula produces feeding deterrents, such as ypaoamide34 and lyngbyatoxin35. Cytotoxic macrolides and peptides have been identified from samples of Moorea bouillonii associated with Alpheus frontalis shrimp in Guam36. Similarly, gorgonian corals are not suitable food because they possess chemical metabolites and mechanical sclerites as defenses against fish37,38, as well as antifungal secondary compounds39. Furthermore, among cnidarian animals, sea fan corals develop cell-based immune defenses (amoebocytes)40. Nevertheless, gorgonian corals, as well as hard corals, are facing a crisis of fungal infestations (e.g., aspergillosis disease in the Caribbean Sea17,41) and algal blooms due to eutrophication.
At our study site, how the cyanobacteria initially settled on the coral is unknown, but the important question is how the bloom is maintained in oligotrophic water. Engene et al.18 showed that Moorea bouillonii lacks heterocysts and genes for nitrogen fixation. Nutrient concentrations at the study site were not high enough from the water surface to 25 m deep to cause algal blooms. Furthermore, cyanobacterial coverage was observed exclusively on Annella. The tube-forming or sewing shrimp Alpheus frontalis H. Milne Edwards have been found in cyanobacterial tubes which they made to live and to eat20,21. Thus, the sewing shrimp Alpheus may play an important role in perpetuating continuous blooms by attaching cyanobacterial filaments to coral branches to form tube-like mats that it uses for food and shelter. NH4 and PO4 excreted from the shrimp were absorbed by Moorea determined in a laboratory experiment (not shown, Yamashiro unpublished data). In addition, this shrimp, like other alpheid shrimp, uses its large claws to snap at other animals and protect its nest made of Moorea42,43. This symbiosis also seems to have a synergistic relationship in respect to nutrition (between photosynthetic cyanobacteria and nitrogen/phosphorus-emitting shrimp).
Concerning the mortality of Annella due to Moorea cover, there are several possible mechanisms. Coral death can be caused by metabolic decline including oxygen depletion or a reduction of food supply to the corals by algal (including cyanobacteria) coverage44,45,46,47. In addition to physical stress, biochemical effects such as allelopathic terpenes secreted by algae have been reported as causing coral death48. Titlyanov49 performed direct contact experiment using cyanobacteria Lyngbya (Moorea) bouillonii on live coral Porites and demonstrated that M. bouillonii acted as a one-sided inhibitor for scleractinian corals inducing bleaching and severe damage of live coral tissue. Similar interaction is often observed in the field of Okinawa Island between M. bouillonii and branching corals such as Montipora nested by sewing shrimp. Coral tissue where filamentous M. bouillonii was tied by the shrimp showed bleached and partial death (not shown). The main cause of octocoral death was not identified in this study, physical effects such as abrasion or oxygen depletion, or biochemical (toxic or allelopathic) effects must be involved.
Some cyanobacteria associated with the coral are able to penetrate into the soft tissue and skeleton50. Our study highlights that Moorea bouillonii is capable of penetrating tissues of gorgonian coral branches by changing its shape at the terminal end. A swollen structure of multiple layers of sheath appears to function as an anchor (Fig. 4B), firmly attaching the cyanobacterium to Annella. Strong persistence of M. bouillonii to the host coral should exist, but this trait has previously been unrecognized. The origin and transmission of the cyanobacterium is still unknown, but the synergy between filamentous Moorea and sewing shrimp and the special trait of penetration found in this M. bouillonii, must allow the persistence of year-round blooms on the sea fan Annella reticulata.
Methods
We first collected filamentous algae entangled on the gorgonian coral colony on 3 March 2009. We measured the concentrations of nutrients (NH4, NO3, NO2 and PO4) using a nutrient autoanalyzer (BL Tech Co.) of triplicate seawater samples collected every 5 m down to 25 m depth on 10 April 2009. We identified the filamentous algae by morphology and molecular information (16S rRNA sequence, see below) and the gorgonian coral by morphological observation of colony structure and sclerites on live or formalin-fixed samples using dissecting or digital microscopes (VHX-1000, Keyence Co.). We also made histological sections to determine the method of attachment of the filamentous algae.
On 17 September 2009, we recorded the height of all gorgonian corals within a 2-m-wide × 20-m-long transect at 18 m depth on a nearly vertical reef with the highest local density of cyanobacteria infection. We recorded the overgrowth (infection) by filamentous cyanobacteria on the coral on all colonies (n = 91) within the transect and classified them into size classes.
The cyanobacterial alga was washed 2–3 times with filtered seawater and stored in CHAOS solution (4 M Guanidine thiocyanate, 0.5% Salkosyl,25 mM Tris-HCl pH8.0,0.1 M 2-Mercaptoethanol after removing extra seawater(partly modified51). Following standard phenol chloroform methods, genomic DNA from the cyanobacterial alga was extracted.
Cyanobacteria-specific PCR primers CYA106F (CGGACGGGTGAGTAACGCGTGA) and CYA781R (an equimolar mixture of CYA 781R(a) (GACTACTGGGGTATCTAATCCCATT) and CYA781R(b) (GACTACAGGGGTATCTAATCCCTTT)52 were used to amplify an about 680-bp region of the 16S rRNA gene. Reaction mixture of 25 µl contained 0.6 µM of each primer, 0.2 mM of each dNTP, 1X PCR Reaction Buffer (TaKaRa), 1.5 mM of MgCl2 solution, 0.08% (w/v) bovine serum albumin, 0.2 U of ExTaq DNA Polymerase (TaKaRa) and 20 ng of template DNA. Amplification was performed with initial melting at 94°C for 3 min, followed by 30 cycles of 94°C for 1.5 min, 59°C for 1 min and 72°C for 2 min and a final extension at 72°C for 5 min. After electrophoresis, PCR products were purified with DNA Cleaner (Wako). The purified PCR products were cloned using TOPO cloning kit (Invitrogen). The totals of twenty clones of sequences were carried out on an automated sequencer CEQ8800 (Beckman Coulter).
References
Hughes, T. Catastrophes, phase shifts and large-scale degradation of a Caribbean coral reef. Science 265, 1547–1551 (1994).
Harvell, C. D. et al. Climate warming and disease risks for terrestrial and marine biota. Science 296, 2158–2162 (2002).
Hoegh-Guldberg et al. Coral reefs under rapid climate change and ocean acidification. Science 318, 1737–1742 (2007).
Paul, V. J., Thacker, R. W., Banks, K. & Golubic, S. Benthic cyanobacterial bloom impacts the reefs of South Florida (Broward Country, USA). Coral Reefs 24, 693–697 (2005).
Kuffner, I. B. et al. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 323, 107–117 (2006).
Nugues, M. M. et al. Algal contact as a trigger for coral disease. Ecol. Lett. 7, 919–923 (2004).
Rasher, D. B. & Hay, M. E. Chemically rich seaweeds poison corals when not controlled by herbivores. Proc. Nat. Acad. Sci. USA 107, 9683–9688 (2010a).
Rasher, D. B. & Hay, M. E. Seaweed alleropathy degrades the resilience and function of coral reefs. Comm. Integ. Biol. 3, 564–566 (2010b).
Paerl, H. W., Hall, N. S. & Calandrino, E. S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Env. 409, 1739–1745 (2011).
Paerl, H. W. & Huisman, J. Bloom like it hot. Science 320, 57–58 (2008).
Elmetri, I. & Bell, P. R. F. Effects of phosphorous on the growth and nitrogen fixation rates of Lyngbya majuscula: implication for management in Moreton Bay, Queensland. Mar. Ecol. Prog. Ser. 281, 27–35 (2004).
Gilbert, P. M. et al. Evidence for dissolved organic nitrogen and phosphorus uptake during a cyanobacterial bloom in Florida Bay. Mar. Ecol. Prog. Ser. 280, 78–83 (2004).
Sharp, K. et al. Phylogenetic and chemical diversity of three chemotypes of bloom-forming Lyngbya species (Cyanobacteria: Oscillatoriales) from Reefs of Southeastern Florida. Appl. Envir. Microbiol. 75, 2879–2888 (2009).
Rützler, K. & Santavy, D. The black band disease of Atlantic coral reefs. P.S.Z.N.I: Mar. Ecol. 4, 301–319 (1983).
Smith, G. W., Ives, L. D., Nagelkerken, I. A. & Ritche, K. B. Caribbean sea fan mortalities. Nature 383, 487 (1996).
Nagelkerken, I. et al. Widespread disease in Caribbean sea fans I. Spreading and general characteristics. Proc. 8th Int. Coral Reef Symp. 1, 679–682 (1996).
Geiser, D., Taylor, J., Ritchie, K. & Smith, G. Cause of sea fan death in the West Indies. Nature 394, 137–138 (1998).
Engene, N. et al. Moorea producens gen. nov., sp. nov. and Moorea bouillonii comb. nov., tropical marine cyanobacteria rich in bioactive secondary metabolites. Int. J. Syst. Evol. Microbiol. 62, 1171–1178 (2012).
Poupin, J. Crustacea Decapoda and Stromatopoda of French Polynesia. Atoll Res. Bull. 451, 1–62 (1998).
Miya, Y. The Alpheidae (Crustacea, Decapoda) of Japan and its adjacent waters. Part II. Publ Amakusa Mar Biol Lab., Kyushu University 3, 103–195, pl. 15–31 (1974).
Fishelson, L. Observations of the littoral fauna of Israel, V. On the habitat and behavior of Alpheus frontalis H. Milne Edwards (Decapoda, Alpheidae). Crustaceana 11, 98–104 (1966).
Hoffmann, L. & Demoulin, V. Marine cyanophycea of Papua New Guinea. II. Lyngbya bouillonii sp. nov., a remarkable tropical reef-inhabiting blue-green alga. Berg. J. Bot. 124, 82–88 (1991).
Engene, N., Coates, R. C. & Gerwick, W. H. 16S rRNA gene heterogeneity in the filamentous marine cyanobacterial genus Lyngbya. J. Phycol. 46, 591–601 (2010).
Box, S. J. & Mumby, P. J. Effect of macroalgal competition on growth and survival of juvenile Caribbean corals. Mar. Ecol. Prog. Ser. 342, 139–149 (2007).
Matson, E. A. Nutrient chemistry of the coastal waters of Guam. Micronesica 24, 109–135 (1991).
Albert, S. et al. Blooms of the cyanobacterium Lyngbya majuscula in coastal Queensland, Australia: disparate sites, common factors. Mar. Poll. Bull. 51, 428–437 (2005).
Lapointe, B. E., Littler, M. M. & Littler, D. S. Macroalgal overgrowth of fringing coral reefs at Discovery Bay, Jamaica: bottom-up versus top-down control. Proc. 8th Int. Coral Reef Sym. 1, 927–932 (1997).
Kuffner, I. B. & Paul, V. J. Effects of nitrate, phosphate and iron on the growth of macroalgae and benthic cyanobacteria from Cocos Lagoon, Guam. Mar. Ecol. Prog. Ser. 222, 63–72 (2001).
Thacker, R. W., Ginsburg, D. E. & Paul, V. J. Effects of herbivore exclusion and nutrient enrichment on coral reef macroalgae and cyanobacteria. Coral Reefs 19, 318–329 (2001).
Hatcher, B. G. Coral reef ecosystems: how much greater is the whole than the sum of the parts? Coral Reefs (suppl.) 16, S77–S91 (1997).
Tsuda, R. T. & Bryan, P. G. Food preference of the juvenile Siganus rostratus and S. spinus in Guam. Copeia 3, 604–606 (1973).
Thacker, R. W., Nagle, D. G. & Paul, V. J. Effects of repeated exposures to marine cyanobacterial secondary metabolites on feeding by juvenile rabbitfish and parrotfish. Mar. Ecol. Prog.Ser. 147, 21–29 (1997).
Gilby, B. L., Burfeind, D. D. & Tibbetts, I. R. Lyngbya majuscula blooms and the diet of small subtropical benthivorous fishes. Mar. Biol. 158, 245–255 (2011).
Nagle, D. G. & Paul, V. J. Chemical defense of a marine cyanobacterial bloom. J. Exp. Mar. Biol. Ecol 225, 29–38 (1998).
Capper, A., Cruz-Rivera, E., Paul, V. J. & Tibbetts, I. R. Chemical deterrence of a marine cyanobacterium against sympatric and non-sympatric consumers. Hydrobiologia 553, 319–326 (2006).
Matthew, S. et al. Cytotoxic halogenated macrolides and modified peptides from the apratoxin-producing marine cyanobacterium Lyngbya bouillonii from Guam. J. Nat. Prod. 73, 1544–1552 (2010).
Puglisi, M. P., Paul, V. J. & Slattery, M. Biogeographic comparisons of chemical and structural defenses of the Pacific gorgonians Annella mollis and A. reticulata. Mar. Ecol. Prog. Ser. 207, 263–272 (2000).
Puglisi, M. P., Paul, V. J., Biggs, J. & Slattery, M. Co-occurrence of chemical and structural defenses in the gorgonian corals of Guam. Mar. Ecol. Prog. Ser. 239, 105–114 (2002).
Kim, K. et al. Fungal disease resistance of Caribbean sea fan corals (Gorgonia spp.). Mar. Biol. 136, 259–267 (2000).
Mydlarz, L. D., Holthouse, S. F., Peters, E. C. & Harvell, C. D. Cellular responses in sea fan corals: granular amoebocytes react to pathogen and climate stressors. PLoS ONE 3, e1811 (2008).
Antonius, A. The ‘band’ diseases in coral reefs. Proc 4th Int Coral Reef Symp 2, 7–14 (1981).
Ritzmann, R. Snapping behavior of the shrimp Alpheus californiensis. Science 181, 459–460 (1973).
Schein, H. The role of snapping in Alpheus heterochaelis Say, 1818, the big-clawed snapping shrimp. Crustaceana 33, 182–188 (1977).
Fabricius, K. E., Genin, A. & Benayahu, Y. Flow-dependent herbivory and growth in zooxanthellae-free soft corals. Limnol. Oceanogr. 40, 1290–1301 (1995).
Birrell, C. L., McCook, L. J., Willis, B. L. & Diaz-Pulido, G. A. Effects of benthic algae on the replenishment of corals and the implications for the resilience of coral reefs. Oceanogr. Mar. Biol. Ann. Rev. 46, 25–63 (2008).
Yamashiro, H., Mikame, Y. & Suzuki, H. Localized outbreak of attached diatoms on the coral Montipora due to low-temperature stress. Sci. Rep. 2, 522 (2012).
Wangpraseurt, D. et al. M.M. In situ oxygen dynamics in coral-algal interactions. PLoS ONE 7, 31192 (2013).
Carpenter, K. E. et al. One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science 321, 560–563 (2008).
Titlyanov, E. A., Yakovleva, I. M. & Titlyanova, T. V. Interaction between benthic algae (Lyngbya bouillonii, Dictyota dichotoma) and scleractinian coral Porites lutea in direct contact. J Exp Mar Biol Ecol. 342, 282–291 (2007).
Miller, A. W., Blackwelder, P., Al-Sayegh, H. & Richardson, L. L. Fine-structural analysis of black band disease-infected coral reveals boring cyanobacteria and novel bacteria. Mar. Ecol. Prog. Ser. 93, 179–190 (2011).
Fukami, H. et al. Conventional taxonomy obscures deep divergence between Pacific and Atlantic corals. Nature 427, 832–835 (2004).
Nübel, U., Garcia-Pichel, F. & Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 63, 3327–3332 (1997).
Acknowledgements
We thank the staff of Zamami Diving Association for informing us of this phenomenon and for their support during sample collection. Thanks to Dr. M. Nonaka of Churaumi Aquarium for identification of the gorgonian coral and to Dr. H. Ohba of Tokyo University Marine Science and Technology for valuable comments on cyanobacteria. A. Kishi and K. Shiroma helped with sample collection and nutrient analysis using the nutrient autoanalyzer. Coral sampling was done with the approval of Okinawa Prefecture, Japan. The underwater photograph (Fig. 2B) was taken by Y. Itayama, from Yomiuri Shimbun. This work was supported by A Grant-in-Aid for Scientific Research (No. 20121002 and No. 25430197) of the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.
Author information
Authors and Affiliations
Contributions
H.Y. designed the entire study, performed most of the experiments and wrote the paper. N.I. performed genetic analysis. K.S. conducted field observation of infected octocorals.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Yamashiro, H., Isomura, N. & Sakai, K. Bloom of the cyanobacterium Moorea bouillonii on the gorgonian coral Annella reticulata in Japan. Sci Rep 4, 6032 (2014). https://doi.org/10.1038/srep06032
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06032
This article is cited by
-
Cyanobacteria-shrimp colonies in the Mariana Islands
Aquatic Ecology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
