
Abstract
Wiwaxia is a bizarre metazoan that has been interpreted as a primitive mollusc and as a polychaete annelid worm. Extensive material from the Burgess Shale provides a detailed picture of its morphology and ontogeny, but the fossil record outside this lagerstätte is scarce and complete wiwaxiids are particularly rare. Here we report small articulated specimens of Wiwaxiafoliosa sp. nov. from the Xiaoshiba fauna (Cambrian Stage 3, Hongjingshao Formation, Kunming, south China). Although spines are absent, the fossils' sclerites – like those of W. corrugata – are symmetrically arranged in five distinct zones. They form rows across the body and were individually added and shed throughout growth to retain an approximately symmetrical body shape. Their development pattern suggests a molluscan affinity. The basic body plan of wiwaxiids is fundamentally conserved across two continents through Cambrian Stages 3–5 – revealing morphological stasis in the wake of the Cambrian explosion.
Similar content being viewed by others


Introduction
The cataphract lophotrochozoan Wiwaxia was first described from isolated sclerites1. These distinctive and widespread scales have been recognized from a range of palaeoenvironmental settings from the early to middle Cambrian (Stage 3–Guzhangian) of western Canada, Utah, Russia, China, eastern Europe and Australia, establishing wiwaxiids as a key component of Cambrian ecosystems2,3,4,5,6,7,8,9,10,11.
Despite this broad geographical occurrence, articulated Wiwaxia specimens are only known from the Burgess Shale12 and two partial specimens from Kaili10,13. The hundreds of complete Burgess Shale specimens denote a scale-covered organism with a creeping foot, distinctive mouthparts and two series of long dorsal spines12,14,15. Sclerites are arranged in four distinct zones on each side of the body, with an additional anterior sclerite zone. The sclerites were initially likened to the fleshy scales (elytra) of certain annelid worms16 – but annelid elytra and Wiwaxia sclerites differ fundamentally in distribution, shape and construction5,12. The flattened chaetae (paleae) of other annelids are more promising homologues5,17 – yet paleae always occur as part of a biramous unit, paired with a ‘normal’ seta. Elements that were putatively interpreted in this role have since been recognized as components of an unrelated organism18; indeed, Wiwaxia sclerites do not form pairs but occur in ‘bundles’15.
On balance, the growth, distribution and microstructure of Wiwaxia sclerites do not bear out an annelid affinity; they instead suggest a relationship with aculiferan molluscs15. Wiwaxia's radula-like mouthparts uphold a molluscan affinity12,14,19; their proposed interpretation as an annelid jaw5,17,20 is not consistent with their detailed anatomy14. The presence of a creeping foot in Wiwaxia15 and the likely molluscan affinity of closely-related Odontogriphus14,21,22 provide further grounds to ascribe Wiwaxia to Mollusca, even if the precise nature of this relationship remains ambiguous.
Whereas the disparate array of isolated sclerites suggests that Wiwaxia was highly diverse3,7, articulated specimens represent just two species, W. taijiangensis and W. corrugata, distinguished based on their sclerites' dimensions and ornament10,15. Here we report a third species of Wiwaxia, represented by articulated specimens from thinly bedded mudstones near the base of the Hongjingshao Formation (Cambrian Series 2, Stage 3), Xiaoshiba, Kunming.
Results
Geological setting
The recently discovered Xiaoshiba lagerstätte within the lower part of the Hongjingshao Formation is characterized by a series of yellow-green or purple-red sandstones, siltstones and shales, occasionally interbedded with yellow, thin-bedded mudstones. It overlies the Yu'anshan Formation (which hosts the Chengjiang lagerstätte) and contains three trilobite genera that also occur in the Chengjiang fauna (Kuanyangia, Yunnanocephalus and Chengjiangaspis) and one (Hongshiyanaspis) that does not. These taxa are found in the upper Qiongzhusian Eoredlichia–Wutingaspis Zone24, although their extensive and poorly-constrained stratigraphic ranges limit their practical utility. Additionally, the Canglangpuian index trilobite Zhangshania has been found about eight metres above the horizon yielding the Xiaoshiba lagerstätte – which is therefore of late Qiongzhusian (Cambrian Series 2, Stage 3) age, rather than early Canglangpuian as previously ascribed25. Biostratigraphically, the Hongjingshao Formation is diachronous in the Kunming area.
Systematic palaeontology
Family Wiwaxiidae Walcott, 1911 (nom. corr. Howell 196226).
Genus Wiwaxia Walcott, 1911.
Type species. Wiwaxia corrugata (Matthew, 1899), Cambrian Stage 5 Ogygopsis Shale, Stephen Formation, Field, Canada.
Wiwaxia foliosa sp. nov.

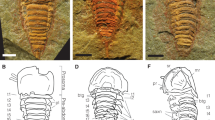
Wiwaxia foliosa sp. nov. from the early Cambrian (Stage 3) of southern China.
(a) YKLP 12121, ventral view of complete specimen. (b) details of the left upper part of (a), showing the symmetrically arranged dorsal sclerites with no more than five ribs, including the marginal ribs. The approximate extent of the transverse sclerite zones is illustrated. Abbreviations: ds, dorsal sclerite; uls, upper lateral sclerite; lls, lower lateral sclerite; vs, ventro-lateral sclerite; L, left; R, right; Z, zone. (c) YKLP 12120 (holotype), ventral view of complete specimen. (d) details of (c), showing incipient ventral sclerites (arrowed) and elongated siculate ventral sclerites, which are not symmetrical about the body's midline. (e) YKLP 12125, lateral view, fully enrolled specimen. (f), YKLP 12124, dorsal surface, showing some sclerites with six ribs (arrowed). (g) YKLP 12122, ventral view of partly-enrolled specimen with anterior portion on higher layer within rock; incipient sclerites arrowed. (h), details of (g), showing sclerite with six ribs (arrowed).
Etymology
foliosus (Latin), many leaves, alluding to the leaf-like arrangement of sclerites.
Holotype
YKLP (Key Laboratory for Paleobiology, Yunnan University) 12120 (Fig. 1c, d), an articulated and almost-complete specimen.
Type locality
Xiaoshiba section, near the bottom of the Hongjingshao Formation, about 8 meters below the Canglangpu Stage Zhangshania Zone (Cambrian Stage 3), near Xiaoshiba village, eastern Kunming, Yunnan.
Material
Six specimens (YKLP 12120–12125, see Supplementary Text online) from the type locality.
Measurements
The smallest specimen (YKLP 12121) is 3.7 mm long and 3.5 mm wide (Fig. 1a); the largest complete specimen (YKLP 12124) is 8.7 mm long, but is not preserved in a position that allows its full width to be accurately measured (Fig. 1f).
Diagnosis
Sclerites unornamented, bearing a single order of ribs. Ribs few in number, typically four per sclerite. Dorsal sclerites symmetrical. Spines absent in specimens under 8 mm in length.
Description
The body is oval in outline (dorsal view) and covered by an imbricate scleritome comprising eight transverse rows (Fig. 2). Prominent dorsal sclerites are aligned along the anterior-posterior axis and form a symmetrical paired series about the sagittal plane. Upper-lateral sclerites are a similar shape; they are arranged in fan-like bundles that radiate from a central attachment point and are slightly erect from the body surface, becoming more so in enrolled specimens. The lower-lateral sclerites are rounded, relatively small and often obscured by other sclerites; their tips are dorsally inclined. Ventro-lateral sclerites are elongate and sickle-shaped; their tips point posteriad. The anterior zone contains symmetrical and sickle-shaped sclerites. Each sclerite bears four to six equally-pronounced ribs. In each sclerite zone, most sclerites are a similar size, but occasional sclerites are smaller and apparently incomplete (Fig. 1d). Sclerite length increases at the same rate as body length, whereas sclerite width increases slightly more slowly (see Supplementary Table S1 online).

Three-dimensional reconstruction of Wiwaxia foliosa sp. nov.
(a) perspective view; checkerboard represents 1 mm; (b) lateral view, indicating partially-erect nature of sclerites; (c) ventral view, indicating broad sweep of dorsal sclerites and elongate nature of ventral sclerites.
Remarks
In terms of shape, aspect ratio and sclerite distribution, the material closely resembles juvenile specimens of W. corrugata from the Burgess Shale15. In the latter case, however, sclerites are generally flush with the body, somewhat obscuring the distinction between separate sclerite zones; here, the distribution of sclerites is more regimented and separate bundles are readily recognized even in fully articulated specimens. The aspect of the upper-lateral sclerites is also distinctive; in the new material, bundles are arranged such that the outermost sclerite is the most lateral, whereas in W. corrugata, the outermost sclerite is closest to the midline. This allows the bundles in W. foliosa to spread more broadly and to overlap the lower-lateral sclerites.
Since the largest articulated specimen is under 9 mm long, it is not possible to determine whether the absence of spines is organic, or ontogenetic (as in W. corrugata).
Discussion
Preservation
Whereas the Burgess Shale fossils are typically preserved in random orientations, the majority of Chengjiang fossils are preserved in a mechanically stable orientation, indicating that they settled under the force of gravity27 rather than in turbulent flow. The same is true of the Xiaoshiba fauna; indeed most of the new Wiwaxia specimens are preserved parallel to the bedding plane in approximately dorso-ventral aspect. The exception is the rolled-up specimen YKLP 12125 (Fig. 1e), indicating that the enrolled organism was stable on its side. The enrolled position cannot have arisen through gravitational settling, but rather implies the presence of an enrolling muscle. Enrolling musculature is present in chitons and juvenile aplacophorans, but not in conchiferan molluscs or other lophotrochozoans28,29.
The absence of larger articulated specimens could result from the Bernoulli effect, where larger objects produce more lift and are thus – if their density is sufficiently low – prone to transport and disarticulation by currents. Such a process seemingly affects other fossils in Xiaoshiba; for instance larger components of the arthropod exoskeleton are routinely transported short distances from the carcass25. This effect may have caused larger Wiwaxia specimens to disarticulate whilst smaller specimens remained intact.
Currents may also have affected the preservation of sclerites in articulated specimens. Because each sclerite is individually attached to the body at the root, some degree of motion – especially in a partially decayed specimen – may have been possible. Furthermore, bundles or ‘fans’ of sclerites may have encountered turbulence and been reoriented as a unit.
Scleritome
In some cases, the corresponding sclerites on opposite sides of the body differ in size. The most extreme case is of ventral sclerites in YKLP 12120 where sclerites are around a third of the typical length. These smaller sclerites correspond in shape and width to the distal part of ventral sclerites and presumably represent sclerites that were incompletely formed when the organism died. Their erratic occurrence indicates that sclerites did not grow synchronously throughout the body; rather, they must have grown to a fixed size, then remained for some time before being individually shed and replaced by a larger sclerite. Because the replacement sclerite, when fully secreted, would be larger than the one it replaced (in order to accommodate the growth of the organism), it would be larger than the opposite sclerite, thus breaking bilateral symmetry (Fig. 1a, c, g). This matches the growth mechanism observed in the spines of W. corrugata15, indicating that the spines grew in the same manner as the body-covering sclerites.
It is also evident that larger individuals bear more sclerites than smaller ones; as with W. corrugata, the dorsal surface grows more rapidly than sclerite width (see Supplementary Table S1 online), meaning that more sclerites are required to cover the expanding area. Accordingly, sclerites are added to each bundle faster than they are shed and the asynchronous addition of sclerites to bundles causes a further deviation from bilateral symmetry.
This uncoordinated growth mechanism has no equivalent in annelid worms, yet bears close relation to the situation in scleritome-bearing molluscs where sclerites grow to a fixed size and are individually replaced30,31,32,33. This periodic deviation from bilateral symmetry adds further support for a molluscan affinity.
The presence of eight transverse rows in the W. foliosa scleritome indicates that this feature is integral to the wiwaxiid body plan, not just a quirk of W. corrugata. This upholds the suggested correspondence between transverse rows in Wiwaxia and aculiferans15. The relationship between Wiwaxia's scleritome and its underlying metameres is ambiguous; the anterior sclerite zone may correspond to a transverse row or may have a separate origin and the metameres may correspond to the rows of sclerites or to the gaps between them (as in aplacophorans34). Depending on the interpretation, Wiwaxia could therefore bear seven to eight (or possibly nine) metameres. This echoes the seven metameres in the aculiferan ground plan (to which polyplacophorans add an eighth late in development29) and the eight iterated units in tryblidid monoplacophorans35. Whether this makes Wiwaxia a stem- or crown-group mollusc depends on relationships within Mollusca, which remain contentious36; either way, the taxon is important for understanding the origin of molluscs' epidermal hard parts19,23. Interpreting the ancestral mollusc as a scleritome-bearing organism would account for the presence of microvillar ‘chaetae’ in early torted conchiferans (Aldanella and Pelagiella)37,38; more speculatively, it would allow the Wiwaxia-like scleritome borne by a hot-vent gastropod39 to represent the reactivation of a dormant genetic machinery15.
Occurrence
The robust nature of Wiwaxia sclerites enhances their preservation potential and accounts for their broad occurrence as carbonaceous fossils, which provides a measure of wiwaxiid distribution through the Cambrian3,4,7,40. This record is complemented by sclerites preserved on bedding planes as the remnants of carbon films2,9,10,11 and more unusually in three dimensions by phosphatization, as in the Cambrian Stage 5 of Australia8 and the Cambrian Stage 3 Qiongzhusian Shuijingtuo Formation in southern China (unpublished material). Despite this wide distribution and good preservation potential, Wiwaxia has not yet been reported from the Chengjiang lagerstätte41, an absence that has been suggested to represent either evolution, biogeography or ecology42. Our new material demonstrates that Wiwaxia had evolved by Chengjiang time and was present in the region. The recalcitrance of Wiwaxia sclerites and the exceptional preservation in Chengjiang makes it difficult to attribute this absence to taphonomic factors. This provides further evidence that the described Chengjiang fauna was ecologically and environmentally distinct from the Burgess Shale43; furthermore, it indicates differentiation between the Chengjiang and Xiaoshiba lagerstätten despite their temporal and geographic proximity.
The new specimens extend the record of articulated Wiwaxia specimens – previously known from the Stage 4 Kaili biota and the Stage 5 Burgess Shale12,13,15 – into Cambrian Stage 3. Differences between the known specimens represent trivial tweaks to the configuration or ornament of sclerites, rather than any meaningful change in body plan – the scleritome exhibits a striking degree of morphological stasis over this 15 million year period. Given this lack of disparity, the Stage 3–5 record of isolated wiwaxiid sclerites can reasonably be reconstructed as components of Wiwaxia-like scleritomes. Wiwaxia therefore joins a growing list of non-mineralised taxa whose morphology changed little from Stage 3 to Stage 5 (e.g. Hallucigenia sparsa44, Nectocaris45), despite the elevated rate of evolution associated with the early Cambrian period46.
Despite their similar body plan, the fossil records of wiwaxiids and halkieriids are more complementary than congruous. Halkieriids have a rich and extensive microfossil record (e.g. ref. 47), but are rarely found articulated17,whereas articulated wiwaxiids are now known from a range of Burgess Shale-type sites, even though their isolated sclerites are rare even in the localities from which they are known7.
Whether these mismatches between the microfossil and macrofossil records reflect biology or taphonomy, the presence of wiwaxiid sclerites (and absence of halkieriid sclerites) in other Burgess Shale-type deposits suggests that halkieriids – like many other constituents of the shelly fauna – were excluded from typical Burgess Shale-type faunas. Similarly, Wiwaxia is yet to be found in the Chengjiang lagerstätte. As such, environmental and ecological factors were clearly intrinsic to the shaping of Cambrian communities.
Conclusions
Wiwaxia's eight sclerite rows are consistently evident in separate species and recall the eight-fold metamerism of the polyplacophorans, aplacophorans and tryblidians – suggesting that this layout was a feature of the molluscan ground plan. Details of the scleritome and the way that it grew suggest that it had much in common with the aculiferan molluscs. Specifically, the piecewise replacement of individual sclerites produced a deviation from bilateral symmetry not seen in annelid worms.
Despite the diverse range of sclerite ornaments and outlines revealed by the small carbonaceous fossil record3,4,41, this new material indicates that the underlying Wiwaxia body plan, once established, remained constant for some 15 million years – seeming to display only species-level variation. Wiwaxia was the most abundant and widespread scleritomorph organism in Burgess Shale-type assemblages, providing a common biological thread between these dispersed communities.
Methods
Fossil material is accessioned in the Key Laboratory for Paleobiology, Yunnan University (YKLP). Specimens were photographed under bright-field illumination using a Leica DFC 500 digital camera mounted to a Stereoscope LEICA M205-C. Statistical data are provided in the Supplementary Table 1.
References
Matthew, G. F. Studies on Cambrian faunas, No. 3. Upper Cambrian fauna, Mount Stephen, British Columbia. The trilobites and worms. Trans. R. Soc. Can. 5, 39–66 (1899).
Fatka, O., Kraft, P. & Szabad, M. Shallow-water occurrence of Wiwaxia in the Middle Cambrian of the Barrandian area (Czech Republic). Acta Palaeontol. Pol. 56, 871–875 (2011).
Butterfield, N. J. Burgess Shale-type fossils from a Lower Cambrian shallow-shelf sequence in northwestern Canada. Nature 369, 477–479 (1994).
Harvey, T. H. P., Ortega-Hernández, J., Lin, J.-P., Zhao, Y. & Butterfield, N. J. Burgess Shale-type microfossils from the middle Cambrian Kaili Formation, Guizhou Province, China. Acta Palaeontol. Pol. 57, 423–436 (2012).
Butterfield, N. J. A reassessment of the enigmatic Burgess Shale fossil Wiwaxiacorrugata (Matthew) and its relationship to the polychaete Canadia spinosa Walcott. Paleobiology 16, 287–303 (1990).
Harvey, T. H. P. & Butterfield, N. J. Macro- and microfossils of the Mount Cap Formation (Early and Middle Cambrian, Northwest Territories). Geosci. Canada 38, 165–173 (2011).
Butterfield, N. J. & Harvey, T. H. P. Small carbonaceous fossils (SCFs): a new measure of early Paleozoic paleobiology. Geology 40, 71–74 (2012).
Porter, S. M. Halkieriids in Middle Cambrian phosphatic limestones from Australia. J. Paleontol. 78, 574–590 (2004).
Ivantsov, A. Y. et al. Palaeoecology of the Early Cambrian Sinsk biota from the Siberian Platform. Palaeogeogr. Palaeoclimatol. Palaeoecol. 220, 69–88 (2005).
Zhao, Y.-L., Qian, Y. & Li, X.-S. Wiwaxia from Early-Middle Cambrian Kaili Formation in Taijiang, Guizhou. Acta Palaeontol. Sin. 33, 359–366 (1994).
Conway Morris, S. & Robison, R. A. More soft-bodied animals and algae from the Middle Cambrian of Utah and British Columbia. Univ. Kansas Paleontol. Contrib. 122, 23–84 (1988).
Conway Morris, S. The Middle Cambrian metazoan Wiwaxia corrugata (Matthew) from the Burgess Shale and Ogygopsis Shale, British Columbia, Canada. Philos. Trans. R. Soc. Lond. B 307, 507–582 (1985).
Sun, H.-J., Zhao, Y.-L., Peng, J. & Yang, Y.-N. New Wiwaxia material from the Tsinghsutung Formation (Cambrian Series 2) of Eastern Guizhou, China. Geol. Mag. 151, 339–348 (2014).
Smith, M. R. Mouthparts of the Burgess Shale fossils Odontogriphus and Wiwaxia: implications for the ancestral molluscan radula. Proc. R. Soc. B 279, 4287–4295 (2012).
Smith, M. R. Ontogeny, morphology and taxonomy of the soft-bodied Cambrian “mollusc” Wiwaxia. Palaeontology 57, 215–229 (2014).
Walcott, C. D. Cambrian Geology and Paleontology II, no. 5. Middle Cambrian annelids. Smithson. Misc. Collect. 57, 109–144 (1911).
Conway Morris, S. & Peel, J. S. Articulated halkieriids from the Lower Cambrian of north Greenland and their role in early protostome evolution. Philos. Trans. R. Soc. Lond. B 347, 305–358 (1995).
Caron, J.-B., Smith, M. R. & Harvey, T. H. P. Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians. Proc. R. Soc. B 280, 20131613 (2013).
Scheltema, A. H., Kerth, K. & Kuzirian, A. M. Original molluscan radula: comparisons among Aplacophora, Polyplacophora, Gastropoda and the Cambrian fossil Wiwaxia corrugata. J. Morphol. 257, 219–245 (2003).
Butterfield, N. J. Hooking some stem-group “worms”: fossil lophotrochozoans in the Burgess Shale. BioEssays 28, 1161–1166 (2006).
Caron, J.-B., Scheltema, A. H., Schander, C. & Rudkin, D. A soft-bodied mollusc with radula from the Middle Cambrian Burgess Shale. Nature 442, 159–163 (2006).
Caron, J., Scheltema, A. H., Schander, C. & Rudkin, D. Reply to Butterfield on stem-group “worms:” fossil lophotrochozoans in the Burgess Shale. BioEssays 29, 200–202 (2007).
Kocot, K. M. et al. Phylogenomics reveals deep molluscan relationships. Nature 477, 452–456 (2011).
Zhang, W.-T. in Biostratigraphy of China (Zhang, W.-T., Chen, P.-J. & Palmer, A. R. eds.) 55–119 (Sci. Press, 2003).
Yang, J., Ortega-Hernández, J., Butterfield, N. J. & Zhang, X.-G. Specialized appendages in fuxianhuiids and the head organization of early euarthropods. Nature 494, 468–71 (2013).
Howell, B. F. in Treatise Invertebr. Paleontol. W (Moore, R. C. ed.) W144–W177 (Geol. Soc. Am., 1962).
Zhang, X.-G. & Hou, X.-G. Gravitational constraints on the burial of Chengjiang fossils. Palaios 22, 448–453 (2007).
Wanninger, A. & Haszprunar, G. Chiton myogenesis: perspectives for the development and evolution of larval and adult muscle systems in molluscs. J. Morphol. 251, 103–113 (2002).
Scherholz, M., Redl, E., Wollesen, T., Todt, C. & Wanninger, A. Aplacophoran mollusks evolved from ancestors with polyplacophoran-like features. Curr. Biol. 23, 2130–2134 (2013).
Blumrich, J. Das integument der chitonen. Zeitschrift für Wissenschaftliche Zool. 52, 404–476, plates 23–30 (1891).
Reinke, J. Beiträge zur bildungsgeschichte der stacheln etc. im mantelrande der chitonen. Zeitschrift für Wissenschaftliche Zool. 18, 305–351, plates 21–22 (1868).
Haas, W. Evolution of calcareous hardparts in primitive molluscs. Malacologia 21, 403–418 (1981).
Fischer, F. P., Maile, W. & Renner, M. Die mantelpapillen und stacheln von Acanthochiton fascicularis L. (Mollusca, Polyplacophora). Zoomorphologie 94, 121–131 (1980).
Scheltema, A. H. & Ivanov, D. L. An aplacophoran postlarva with iterated dorsal groups of spicules and skeletal similarities to Paleozoic fossils. Invertebr. Biol. 121, 1–10 (2002).
Lemche, H. M. & Wingstrand, K. G. The anatomy of Neopilina galatheae Lemche, 1957 (Mollusca, Tryblidiacea). Galathea Rep. 3, 9–73 (1959).
Telford, M. J. & Budd, G. E. Invertebrate evolution: bringing order to the molluscan chaos. Curr. Biol. 21, R964–R966 (2011).
Dzik, J. & Mazurek, D. Affinities of the alleged earliest Cambrian gastropod Aldanella. Can. J. Zool. 91, 914–923 (2013).
Thomas, R. D. K., Vinther, J. & Matt, K. Structure and evolutionary implications of finely preserved chaetae associated with Pelagiella, a stem-group gastropod from the Kinzers Formation (Early Cambrian) at Lancaster, Pennsylvania. in Int. Palaeontol. Congr. 3, London, U.K. Program. Abstr. 375 (2010).
Warén, A., Bengtson, S., Goffredi, S. K. & Van Dover, C. L. A hot-vent gastropod with iron sulfide dermal sclerites. Science 302, 1007 (2003).
Harvey, T. H. P., Vélez, M. I. & Butterfield, N. J. Small Carbonaceous Fossils from the Earlie and Deadwood Formations (Middle Cambrian to Lower Ordovician) of southern Saskatchewan. Summ. Investig. 4.1, A–1 (108) (2012).
Zhao, F. et al. Diversity and species abundance patterns of the early Cambrian (Series 2, Stage 3) Chengjiang Biota from China. Paleobiology 40, 50–69 (2014).
Butterfield, N. J. Exceptional fossil preservation and the Cambrian Explosion. Integr. Comp. Biol. 43, 166–177 (2003).
Zhao, F., Zhu, M. & Hu, S. Community structure and composition of the Cambrian Chengjiang biota. Sci. China Earth Sci. 53, 1784–1799 (2010).
Steiner, M., Hu, S.-X., Liu, J. & Keupp, H. A new species of Hallucigenia from the Cambrian Stage 4 Wulongqing Formation of Yunnan (South China) and the structure of sclerites in lobopodians. Bull. Geosci. 87, 107–124 (2012).
Smith, M. R. Nectocaridid ecology, diversity and affinity: early origin of a cephalopod-like body plan. Paleobiology 39, 297–321 (2013).
Lee, M. S. Y., Soubrier, J. & Edgecombe, G. D. Rates of Phenotypic and Genomic Evolution during the Cambrian Explosion. Curr. Biol. 23, 1889–1895 (2013).
Parkhaev, P. Yu. & Demidenko, Yu. E. Zooproblematica and Mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontol. J. 44, 883–1161 (2010).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (41272027) and Ministry of Education of China (20115301110001). M.R.S. is supported by Clare College, Cambridge. We thank H.Q. Zhang for specimen preparation.
Author information
Authors and Affiliations
Contributions
J.Y. arranged fieldwork, J.Y., J.B.H. and T.L. collected material; X.G.Z. and J.Y. conceived the project; X.G.Z. and M.R.S. conducted research, M.R.S. wrote the paper with input from X.G.Z. and T.L.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Info
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported license. The images in this article are included in the article's Creative Commons license, unless indicated otherwise in the image credit; if the image is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the image. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Yang, J., Smith, M., Lan, T. et al. Articulated Wiwaxia from the Cambrian Stage 3 Xiaoshiba Lagerstätte. Sci Rep 4, 4643 (2014). https://doi.org/10.1038/srep04643
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep04643
This article is cited by
-
Sclerite-bearing annelids from the lower Cambrian of South China
Scientific Reports (2019)
-
Formation and structure of paleae and chaetal arrangement in chrysopetalidae (Annelida)
Zoomorphology (2019)
-
Orthrozanclus elongata n. sp. and the significance of sclerite-covered taxa for early trochozoan evolution
Scientific Reports (2017)
-
A predatory bivalved euarthropod from the Cambrian (Stage 3) Xiaoshiba Lagerstätte, South China
Scientific Reports (2016)
-
New reconstruction of the Wiwaxia scleritome, with data from Chengjiang juveniles
Scientific Reports (2015)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
