
Abstract
Clinical trials have shown that antioxidant supplementation increased the risk of lung and skin cancers, but the underlying molecular mechanism is unknown. Here, we show that epigallocatechin gallate (EGCG) as an exemplary antioxidant induced significant death and DNA damage in human lung and skin normal cells through a reductive mechanism. Our results show direct evidence of reductive DNA damage in the cells. We found that EGCG was much more toxic against normal cells than H2O2 and cisplatin as toxic and cancer-causing agents, while EGCG at low concentrations (≤100 μM) increased slightly the lung cancer cell viability. EGCG induced DNA double-strand breaks and apoptosis in normal cells and enhanced the mutation frequency. These results provide a compelling explanation for the clinical results and unravel a new reductive damaging mechanism in cellular processes. This study therefore provides a fresh understanding of aging and diseases and may lead to effective prevention and therapies.
Similar content being viewed by others

Introduction
Lung cancer and skin cancer are two of the most common cancers in the world. Lung cancer is a leading cause of cancer death in men and women in the United States, while skin cancer has the highest incident rate. It is estimated that 228,190 people in the United States will be newly diagnosed with lung cancer in 2013 (ref. 1). The skin cancer has a lower death rate, but its incident rate continues to rise at an increment 3% per year2. Thus, both lung and skin cancers have drawn significant attention in prevention, diagnosis and therapy.
It is long thought that antioxidants kill reactive oxygen species (ROS) produced in normal cellular processes and may therefore protect cells from oxidative damage. Therefore, there is increasing use of dietary and cosmetic antioxidants in attempts to slow down the aging process and to prevent the development of diseases such as cancer and heart disease. However, a recent systematic review including data from 78 randomized clinical trials with totally 296,707 participants, of which 26 trials included 215,900 healthy participants, indicates that the antioxidants β-carotene, vitamin A, vitamin C, vitamin E and selenium showed either no beneficial health effects, or a small increase in human mortality3,4. In particular, a cancer prevention study entitled the Alpha-Tocopherol (vitamin E)/Beta-Carotene Cancer Prevention Study (ATBC) demonstrated that lung cancer rates of male smokers increased significantly with β-carotene5,6. Another β-Carotene and Retinol Efficacy Trial (CARET) study also demonstrated a significant increase in lung cancer associated with antioxidants7,8. Subsequent studies confirmed these adverse effects9, which were seen in non-smokers as well4. A recent clinical trial also showed that antioxidant supplementation increased the risk of skin cancer10. Overall, these clinical trials have shown significant evidence that antioxidants such as β-carotene increased the incidences of lung and skin cancers. However, the underlying molecular mechanism is essentially unknown. This study aims to unravel the molecular mechanism.
Antioxidants may also cause direct damage to DNA and the cell, as they are rich in weakly-bound electrons. The latter are well-known to initiate molecular reactions in many physical, chemical and biological systems11,12. Using the innovative femtomedicine concept12, our group has demonstrated that dissociative electron transfer (DET) reactions with weakly-bound epre− play key roles in many biological processes, ranging from DNA strand breaks13,14 to the activation of anticancer drugs15,16,17. In particular, we found that the DET of weakly-bound epre− to the guanine (G) base is highly effective in inducing chemical bond breaks13 and then single-strand and double-strand breaks (SSBs and DSBs) of the DNA in aqueous solutions14. DSBs of DNA in the cell are difficult to repair and directly relate to genetic mutation, apoptosis or cancer initiation18. Thus, the reductive damage represents a previously unrecognized mechanism of cellular processes, which may have far-reaching significance to the understanding of the aging process and to the prevention and cure of challenging human diseases.
Emerging evidence also supports the positive physiological role of oxidizing ROS such as H2O2 as a necessary ‘evil’ for cell signaling19. Recent research also showed evidence in mice that oncogenes actively promoted a ROS detoxification program, that is, enhanced intracellular antioxidant and ROS detoxication may in fact be pro-tumorigenic20,21. In view of recent data strongly implying that much of late-stage cancer's incurability may be due to its possession of too many antioxidants, Watson22 recently wrote “the time has come to seriously ask whether antioxidant use much more likely causes than prevents cancer”.
Green tea (GT) has been consumed in the world wide and the GT extracts have widely been used in various beverages, health foods, dietary supplements and cosmetic items23. Particularly, epigallocatechin gallate (EGCG), the most abundant catechin (flavonoid) in green tea, is often used as an icon of antioxidants. EGCG and other flavonoids were reported to be beneficial in treating prostate, cervical and bladder cancers24,25,26. But contradictory explanations of the results exist in the literature: some researchers suggested that the cytotoxic effect is associated with the formation of intracellular H2O2 by EGCG27,28,29, where others proposed that EGCG has a protection role against H2O2-induced cell death30,31,32. An interesting study by Hsu and co-workers33 using exogenous catalase and H2O2 concluded that the EGCG-induced cytotoxic effects on tumor cells result mainly from sources other than H2O2. There are also potential detrimental health effects. High intake of flavonoids (tea or coffee) during pregnancy is suspected to increase the risk of infant leukemia and childhood malignant central nervous system tumours34,35,36. Moreover, researchers also observed that plant extracts such as phytoestrogens (Genistein, coumestrol, quercetin, zearalenone and resveratrol) induced genotoxicity and mutagenesis in mammalian cells, suggesting the possible involvement of mutagenicity in initiating phytoestrogen-induced carcinogenesis37,38,39. In contrast, Fox et al40. reported that resveratrol, genistein and baicalein caused genotoxicity but no mutagenesis and hence proposed that these antioxidants are attractive candidates for improved chemotherapeutic agents. Our group41 did observe that some (but not all) antioxidants at a concentration of ≥100 μM enhanced the cytotoxicity of cisplatin in killing cervical, ovarian and lung cancer cells. These contradicting results and explanations, briefly summarized above, indicate that the key mechanism for (either beneficial or adverse) health effects of antioxidants (in the absence of an exogenous chemotherapeutic agent) is most likely unknown.
Here, we hypothesize that antioxidants are potent to cause reductive damage or death to the cell, which may provide a mechanistic understanding of the clinical trial results on lung and skin cancers. We used EGCG as an exemplary antioxidant for two major reasons. EGCG has a reduction energy (ER) of +0.43 eV, which is comparable to ER = +0.48 eV for α-tocopherol and +0.69 eV for β-carotene at standard conditions (pH 7.0, 20°C)42,43,44. More importantly, unlike other antioxidants such as α-tocopherol and β-Carotene insoluble in water, EGCG has excellent stability and water solubility and can therefore be readily tested in cell-culture experiments. 1. We prepared a GT stock solution from a commercial GT and measured time-series EGCG concentrations released in the GT solution using a UV/Vis spectrophotometer. 2. We measured UV absorption spectra to show the reductive reaction of nucleotide dGMP with EGCG/GT and the oxidative reaction of dGMP with hydrogen peroxide (H2O2) that is a major ROS in oxidative stress. We also prepared independently the samples of EGCG+ by reaction of EGCG with HCl and of (G-H)−/G− by reaction of dGMP with epre− uniquely produced by femtosecond (fs) laser pulses13,14 and measured their respective UV absorption spectra. 3. Cell cultures from two human lung diploid fibroblasts (WI-38 and MRC-5 cell lines), a human skin diploid fibroblast (GM05757) and a human lung cancer cell line (A549) were used to test the biological effects of EGCG/GT. WI-38, MRC-5 and GM05757 are often used as human normal cells in cancer research. For comparison, oxidative damage was mimicked by exposing cultured cells to various concentrations of H2O2. And to evaluate the carcinogenic potential of EGCG/GT, we also used cisplatin (CDDP, a highly toxic chemotherapeutic drug and carcinogenic agent) as a second reference. 4. We showed evidence of reductive DNA damage in the cells treated by EGCG and presented a method for quantitative measure of the damage yield. 5. We measured cell viability, DNA DSBs and apoptosis induced by EGCG/GT. 6. We also studied the effects of GT and EGCG as well as their combinations with CDDP on human lung cancer (A549) cells to study the effect of EGCG/GT on the chemotherapy. 7. To test the mutagenicity of EGCG, we conducted the hypoxanthine phosphorybosyl transferase (HPRT) assay using Chinese hamster ovary (CHO) cells, which is widely used as a model to investigate gene mutations in mammalian cell lines45.
Results
Electronic absorption spectroscopic measurements
To determine the EGCG concentration in the commercial green tea (GT) stock solution, we measured UV absorption spectra of EGCG and GT solutions, as shown in Figure 1. The results show that EGCG was quickly released in the GT solution made by adding boiling water into to the GT (leaves) at 1.16 g/l, which is about 10 times less than the typical amount (~2.5 g in 250 ml) in a cup of GT in daily drinking. An equivalent EGCG concentration of ~133 μM was produced in the GT solution at ~1.0 hr after boiling water was added and it further increased by about 20% beyond 4.0 hr. This indicates that EGCG is indeed quite rich and quickly released in daily consumed GT. In Figure 1, the absorption spectrum of the GT solution looks very similar to that of EGCG, though there are also some differences. The pure EGCG has an absorption peak at 273.5 nm and little absorption at wavelengths ≥325 nm, while the GT solution has the main peak slightly blue shifting to 271.5 nm, a weaker absorption at wavelengths below 240 nm and a tail extending up to 400 nm. These indicate the presence of other catechins or components in the GT solution. In all our experiments with GT, we simply calibrated the absorbance at 273.5 nm for estimating the EGCG concentration, which is surely the upper limit of the true EGCG concentration in the GT solution.

UV absorption spectra of 133 μM EGCG in water and the green tea (GT) solution.
The GT solution was prepared with various dissolving times after boiling water was added to the GT (leaves) at 1.16 g/l and followed by naturally cooling at room temperature. The results show that EGCG is quite abundant and quickly released in green tea. The absorption spectrum of the GT solution looks very similar to that of EGCG, while there are also some differences, indicating that there were impurities (other compounds or chemical states) in the GT solution.
We then measured UV (electronic) absorption spectra of 50 μM dGMP, 50 μM EGCG/GT and their mixtures at zero and 24 hr, as well as their difference spectra, as shown in Figures 2A and B. To identify the reaction products clearly, we also prepared the samples of EGCG+ and (G-H)−/G− independently and their absorption spectra are also shown in Figure 2A (See also Supplementary Information and Figures S1 and S2). It is well-known that the UV absorption spectra of nucleotides or DNA arise solely from the electronic excitation in the rings of the bases made of alternating single and double bonds. For dGMP, there are a main electronic absorption peak at 253 nm and a shoulder peak around 272 nm. Interestingly, the obtained difference spectra of the dGMP-EGCG/GT mixtures between zero and 24 hr durations showed a pronounced positive peak at ~251 nm, a minimum at ~272 nm, a second peak around 288 nm and a broad absorption band at wavelengths above 300 nm. Compared with the absorption spectrum of EGCG+ (Figure 2A), the peak at 251 nm for the difference spectra can readily be attributed to the EGCG+ arising from the reaction of dGMP with EGCG/GT. Our first observation of the absorption spectrum of the stable (G-H)−/G− anion uniquely produced by the reaction of dGMP with prehydrated electrons generated by fs laser pulses13,14 shows that there are a main peak at ~266 nm and a broad absorption band above 300 nm (extending to the visible range at 300–600 nm). Interestingly, the DFT calculations by Naumov and von Sonntag46 of the absorption spectrum of the (G-H)•− anion radical predicted a strong band at 266 nm, two very strong bands at 311 and 375 nm and a weaker band at 531 nm. The latter three absorption bands were comparable with those obtained experimentally by Faraggi and Klapper47 for the anion radical with absorption bands at 340, 400 and 510 nm and intensities lower with higher wavelengths. Thus, the absorption band above 300 nm observed for the present dGMP-EGCG/GT mixtures can reasonably be attributed to the formed (G-H)−/G−. Note that the observed main peak at 266 nm for (G-H)−/G− did not appear in the present spectra for the mixtures, but this can be well explained by its overlap with the negative absorption band centered around 275 nm of EGCG+ (due to the depletion of neutral EGCG). The sum of the absorption bands of EGCG+ and (G-H)−/G− gave rise to the observed peak at ~288 nm for dGMP-EGCG/GT mixtures. These spectral features give direct evidence of the electron transfer from EGCG/GT to the G base in dGMP. The results also show that the electron transfer from the GT to dGMP is less significant than that from EGCG. This is reasonable, as the indicated EGCG concentration in the GT was somewhat below the true EGCG concentration (Figure 1).

Spectroscopic observations of the reductive reaction of the nucleotide dGMP by EGCG/GT and of the oxidative reaction of dGMP by H2O2 in air-saturated water.
A–B: UV (electronic) absorption spectra of the mixtures of 50 μM EGCG/GT and 50 μM dGMP at zero and 24 hr at 37°C, as well as their difference spectra; the absorption spectra of the EGCG+ cation and the (G-H)−/G− anion are also shown in A (see text and Supplemental Information). These results give direct evidence of the electron transfer from EGCG/GT to the G base in dGMP. C: The UV spectrum of the dGMP-H2O2 mixture showed extremely small changes within 24 hr even with higher H2O2 concentrations (200–500 μM); the difference spectrum shows clearly three absorption peaks at ~210, 260 and 290 nm, respectively, which are the characteristic absorption of the 8-oxo-dG (the well-known biomarker of oxidative damage). Note that there are contrast differences between difference spectra of EGCG/GT-dGMP and H2O2-dGMP mixtures, especially at wavelength ranges below 250 nm and above 300 nm, reflecting the difference between reductive and oxidative reactions of dGMP.
To compare with the oxidative reaction by H2O2, we also measured time-series UV spectral changes of the dGMP-H2O2 mixture. As shown in Figure 2C, the UV spectrum of the dGMP-H2O2 mixture showed extremely small changes within 24 hr even with higher H2O2 concentrations (200–500 μM). In contrast to the time-series difference spectra of dGMP with EGCG/GT, the difference spectrum of the dGMP-H2O2 mixture showed three characteristic absorption peaks at 210, 260 and 290 nm, respectively, while there was almost zero absorption at wavelengths ≥300 nm. These are the characteristic absorption of the 8-oxo-dG48,49, which is a well-known biomarker of oxidative DNA damage.
The spectra in Figures 2A–C clearly show that the reaction of dGMP with EGCG/GT was distinctly different from that with H2O2 and the reductive reaction of dGMP by 50 μM EGCG/GT was far more effective than the oxidative reaction by 500 μM H2O2. Even if there was a small amount of H2O2 produced in the air-saturated EGCG/GT + dGMP solution (it is unlikely as EGCG/GT in water under air is well-known to be quite stable), the oxidative reaction of the G base with any formed H2O2 must be negligible. Otherwise, the characteristic absorption peaks of the oxidative-damage biomarker (8-oxo-dG) would be observed in Figures 2A and B. This was not observed. Thus, the reductive reaction of G with EGCG/GT must be a dominant process. In contrast to the characteristic absorption of the oxidative damage biomarker, the distinct feature of reductive DNA damage is obviously the characteristic absorption band at ≥300 nm, which is absent for the dGMP-H2O2 mixture but present for the dGMP-EGCG/GT mixtures. This characteristic absorption band can be considered as a biomarker of reductive DNA damage.
Direct measurements of intracellular reductive DNA damage induced by EGCG
The normal cells (GM05757) were treated by various concentrations (control, 30 and 50 μM) of EGCG added to the culture medium for 24 hr and then washed thoroughly by PBS after removal of the medium. The reductive DNA damage in the treated and washed cells was directly characterized by measuring the absorption spectra at 300–600 nm and its yield was given by the integrated spectral intensity over 300–600 nm (See Methods). The results are shown in Figure 3. It is clearly seen that the treatment of EGCG enhanced the yield of reductive DNA damage in the cells significantly.

Spectral measurements of reductive DNA damage in the cells treated by EGCG.
A. The absorption spectra at 300–600 nm of the normal cells (GM05757) with/without the 24 hr treatment of 30 and 50 μM EGCG; B. The yield of reductive DNA damage given by the integrated spectral intensity over 300–600 nm. The results show significant enhancements in reductive DNA damage in the cells treated by EGCG.
Cell viability assays
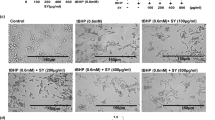
We then investigated the effect of various concentrations of EGCG on the growth and viability of human lung (MRC-5 or WI-38) and skin (GM05757) normal cells seeded in 96-well plates and compared it with the effects of H2O2 and CDDP. As shown in Figures 4A–C, a 24 hr incubation of the normal cells with H2O2 led to increased cell killing with rising H2O2 concentrations up to 100 μM, at which less than 10% of the treated cells survived. And the CDDP-treated cells showed similar viability curves to those for the H2O2-treated cells. These results are not surprising, in view of the well-known oxidative stress induced by H2O2 and the severe toxicity of CDDP as a chemotherapeutic drug and a potent carcinogenic agent. The striking result is, however, that a similar treatment with EGCG decreased the cell survival rate in a dose-dependent manner and the normal cells were killed much more effectively by EGCG than by H2O2 and CDDP (P < 0.007). These results strongly demonstrate that EGCG is highly toxic against human lung and skin normal cells.

MTT cell viability assays of lung and skin normal cells and lung cancer cells with treatments of EGCG/GT/CDDP/H2O2 and the combination treatments of CDDP with EGCG/GT.
The cells were seeded in 96-well plates and after overnight incubation, treated with various agent concentrations for 24 h. The viability values are represented as percents with respective to the untreated cells (regarded as 100% viability). The bars demonstrate the means of triplicate experiments with SD. A, B and C show the striking result that the lung normal cells (WI-38 and MRC-5) and skin normal cells (GM05757) were killed much more effectively by EGCG than by the oxidizing H2O2 and the potent carcinogenic agent CDDP. D, E and F show that though less toxic than EGCG, GT also caused a significant killing of the normal cells, compared with H2O2 and CDDP. In contrast, G shows that at concentrations of ≤100/150 μM, EGCG/GT exhibited an increase by 10–20% in the growth of lung cancer cells, while at very high concentrations of ≥100/150 μM, EGCG/GT showed some killing effect. H and I show that the combination treatment of CDDP with 100 μM EGCG or 150 μM GT showed a small enhancement by about 10% in the survival of lung cancer cells, compared with the treatment of CDDP alone.
We also compared the toxic effects of the pure EGCG and the GT. The results in Figures 4D–F clearly show that significant cell killing was still observed for the normal cells treated with the GT solution, though it was less effective than the pure EGCG. The latter is consistent with the difference in electron transfer effectiveness between EGCG to dGMP and GT to dGMP (See Figure 2). These results indicate that the other chemical components in the GT either contributed to the UV absorption at 273.5 nm (so that the shown EGCG concentrations in the GT solutions were overestimated) or neutralized the toxic effect of EGCG to some extent. Nevertheless, the observation of a significant toxicity of GT is still surprising, compared with those of the oxidizing ROS H2O2 and the potent carcinogenic agent CDDP (see Figures 4A–C). As shown in Table 1, the values of IC50 (the agent concentration required to kill 50% of untreated cells) of EGCG, GT, CDDP and H2O2 were measured to be 17–19, 40–60, 38–85 and 50–75 μM, respectively, depending on the cell lines. These results strongly indicate that EGCG and concentrated GT are indeed highly toxic against human lung and skin normal cells.
Furthermore, to explore the potential beneficial effect of EGCG and GT in treating lung cancer, we also investigated the effects of various concentrations of CDDP, EGCG and GT, as well as combinations of CDDP with 100 μM EGCG or 150 μM GT, on the growth and viability of human lung cancer (A549) cells. The results are shown in Figures 4G–I. First, it can be seen that a significant killing of the cells was observed at low CDDP concentrations, but about 40% of the treated A549 cells still survived even at very high CDDP concentrations of 200–500 μM (Figure 4G). This confirms the strong resistance of A549 cells to CDDP, as observed previously41. Second, in striking contrast to the results for normal cells (Figures 4A–C), EGCG or GT exhibited an increase by 10–20% in the viability of lung cancer cells at lower concentrations of ≤100 or 150 μM, while a killing effect was observed at very high concentrations of 100–400 or 150–500 μM (Figure 4G). Third, the combination of CDDP with 100 μM EGCG or 150 μM GT resulted in an increase by ~10% in the survival of the A549 cells, compared with the treatment of CDDP only (Figures 4H and I). As also shown in Table 1, the IC50 values of EGCG, GT, CDDP, CDDP plus 100 μM EGCG and CDDP plus 150 μM GT on A549 lung cancer cells are 275, ≥500, 300, 400 and ≥400 μM, respectively. These values are approximately one order of magnitude larger than those of EGCG/GT on WI-38, MRC-5 and GM05757 normal cells. Thus, we conclude that EGCG/GT caused a slight increase in the survival of lung cancer cells at concentrations of ≤100/150 μM, while EGCG/GT showed some killing effect at very high concentrations of 100–400 or 150–500 μM. The latter result is generally consistent with those reported in the literature27,28,33. The results also indicate that the combination treatment of CDDP with EGCG/GT showed no positive effects, compared with the treatment of CDDP only.
Imaging of DNA double-strand breaks (DSBs) and toxicity
Given the cell viability results shown in Figure 4 and Table 1, we investigated the genetic toxic effect of EGCG in the treated human normal cells and compared it with the oxidative damage induced by H2O2. We used the HCS DNA Damage Kit (Invitrogen), which was developed to enable simultaneous quantitation of two cell health parameters, genotoxicity and (cyto-)toxicity. The phosphorylated H2AX (γH2AX) foci formed at the damage site in the nucleus are a biomarker of DNA DSBs, which were measured by specific antibody-based detection. Toxicity was also measured with the Image-iT® DEAD Green™ viability stain. The assay kit also included Hoechst 33342 (a DNA-binding dye emitting blue fluorescence), which shows the nuclear morphology of all normal and damaged cells. Figure 5A and B show the fluorescence images of the treated lung normal (MRC-5) cells and the DNA DSB yield as a function of EGCG/H2O2 concentration, respectively. It can be seen that there was only a slight increase by a factor of ≤2 in DNA DSB yield with increasing H2O2 concentrations up to 30 μM, whereas EGCG at 30 μM caused a much larger enhancement by a factor of 4–5 of DNA DSB yield in the treated cells. Moreover, the cell images shown in Figures S3 and S4 in Supplementary Information also demonstrate that in contrast to H2O2, EGCG led to serious cell injuries, including plasma membrane permeability, in the treated MRC-5 and GM05757 cells. These results are consistent with the cell viability results shown in Figure 4 and Table 1. We therefore conclude that EGCG is highly toxic and effective in inducing DNA DSBs in human lung and skin normal cells.

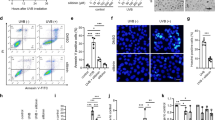
Imaging and analysis of genotoxicity of EGCG and H2O2 in MRC-5 cells using the HCS DNA Damage Kit.
MRC-5 cells were treated with various concentrations of EGCG/H2O2 for 12 hours at 37°C, 5% CO2. Images in (A) show that at the concentration of 30 μM, a large number of γ-H2AX foci were observed in the MRC-5 cells treated with EGCG, but not with H2O2; Hoechst 33342 was used to map nuclei. The graph (B) shows the γ-H2AX intensity in MRC-5 cells versus EGCG/H2O2 concentration, where quantitative analyses of activated γ-H2AX were performed using an Image J software.
Apoptosis and DNA fragmentation assay
It is known that DNA DSBs likely lead to apoptosis. We therefore measured the EGCG-induced apoptosis. A landmark of cellular self-destruction by apoptosis is the activation of nucleases that eventually degrade the nuclear DNA into fragments. Detection of these fragments is relatively straightforward to quantify apoptotic cells and can be done using the APO-BrdU TUNEL assay. As shown in Figure 6 for GM05757 cells grown in T25 flasks, the treatment of EGCG resulted in an enhancement of apoptosis in a dose-dependent manner. The fraction of the BrdU-positive (apoptotic) cells (i.e., cells exhibiting DNA fragmentation) was increased from 2.0% at 0 μM EGCG (the control) to 44.5% at 30 μM, 57.8% at 50 μM and 74.1% at 70 μM EGCG. These data clearly show that EGCG resulted in large enhancements in apoptosis of the normal cells.

APO-BrdU DNA fragmentation assay of GM05757 cells treated with EGCG.
Cells were grown in T25 flasks with the treatment of various EGCG concentrations for 24 h. Cells in the right side of the vertical line in each histogram are BrdU-positive cells (cells exhibiting DNA fragmentation), which represent apoptotic cells.
HPRT gene mutation assay for CHO-K1 cells
The mutagenicity of EGCG was finally tested in CHO-K1 cells using the HPRT assay, which has been used to detect chemicals capable of causing DNA damage that leads to gene mutation. Various types of mutations in the HPRT gene, which is located on the X-chromosome of mammalian cells, lead to cells resistant against lethal 6-TG incorporated into their DNA. Chinese hamster ovary (CHO) cells have become the main cell line for the HPRT assay45,50.
The results for the cytotoxicity and mutagenicity of EGCG on CHO cells treated for 24 hr are shown in Figure 7. Plating efficiencies of the cells decreased from 95% for the control (in the medium with no EGCG) to 46% for the treatment with 100 μM EGCG (Figure 7A); the 7-day clonogenic survival rate of the treated cells decreased with increasing EGCG concentrations to 49% at 100 μM EGCG (Figure 7B). The results for the induction of 6-TG-resistant mutants after treatment with various concentrations of EGCG are shown in Figure 7C. The mutation frequency (MF) showed a clear dose dependence on EGCG. The MFs for the cells treated with various EGCG concentrations (20–100 μM) were significantly higher than the spontaneous MF of the cells in the control. At EGCG concentrations of 50 and 100 μM, the MFs were 78 and 101 mutants/106 clonable cells, respectively, significantly higher than the spontaneous MF (less than 25 mutants/106 clonable cells) for CHO cells. Interestingly, the observed cloning efficiencies for the cells surviving from the 24 hr EGCG treatment and the post-growth of 7 days in the normal culture medium did not decrease with increasing treatment concentrations of EGCG (Figure 7D). This indicates that the induced genetic mutations did not affect the clonogenic ability of the survived cells, which may lead to carcinogenesis.

Plating efficiencies (A), clonogenic survival fraction (B), mutation frequencies at the HPRT gene (C) and cloning efficiencies of the CHO cells in control and treated with various concentrations of EGCG (D).
Bars are the mean +/− SD of three independent assays.
Discussion
We have demonstrated clear evidence of the reductive reaction of nucleotide dGMP with EGCG/GT, which is much more effective than the oxidative reaction of dGMP with H2O2. We have also presented a quantitative measure method and shown clear evidence of reductive DNA damage in the cells treated by EGCG. Correspondingly, we have found that EGCG showed a far more severe toxic effect against human lung and skin normal cells than the oxidizing ROS (H2O2) and the highly toxic and carcinogenetic agent CDDP. Furthermore, we have demonstrated that EGCG not only induced significant DNA double-strand breaks and apoptosis in normal cells but led to a high genetic mutation frequency. Moreover, we have shown that EGCG caused slight increases by ~20% and ~10% in the viability and the survival of lung cancer cells without and with the treatment of cisplatin, respectively. These results are consistent with the results of clinical trials3,4,5,6,7,8,9,10 and with the recent finding that reductive DNA damage induced by weakly-bound electrons is far more potent than oxidative damage induced by OH• radicals produced by ionizing radiation13,14.
The finding of a more lethal reductive pathway appears quite puzzling, as oxidative stress has long been thought to be a major mechanism for causing damage or death to the cell and for aging and many human diseases such as cancer and heart disease. However, this puzzling can be solved if the following mystery about the exact functions of ROS in physiological and pathological processes is revealed. It is true that superoxide anion (O2−), conventionally classified as a ROS, is biologically extremely toxic and may contribute to the pathogenesis of many diseases and perhaps also to aging, though direct observation of its rapid reactions is challenging in techniques51,52. And it is critical for the enzyme superoxide dismutases (SODs) to suppress the activity of O2− by catalyzing the dismutation of O2− into O2 and H2O251. On the other hand, it is also known that when thought as an oxidant, O2− cannot attack polyunsaturated lipids or DNA significantly51. This ‘contradiction’ seems caused by the misconception that O2− would act as an oxidant in these processes. O2− is actually a strong reducing agent in chemistry. O2 has a well-known electron affinity of 0.451 eV only53 and O2− has no positive electron affinity54. In aqueous media, the dominant chemistry of O2− is that of a strong Bronsted base (O2− + HA → ½H2O2 + ½O2 + A−), where O2− donates an electron52,54. This is exactly the case for the well-known Fenton reaction catalyzed by Fe3+/Cu2+, converting O2− into H2O2 or OH•52.
Our present results further demonstrate that EGCG was highly effective in injecting a weakly-bound electron into the G base and resulted in severe reductive DNA damage and cell death. Owing to the fact that the reduction energy ER = +0.43 eV of EGCG42,43 is nearly identical to ER = +0.33 eV of O2− (ref. 54), our experiments equivalently demonstrate that O2− can be effective in inducing reductive damage to DNA and the cell. This deduction can reasonably explain not only the observed critical role of the enzyme SODs to suppress the activity of O2− in physiological and pathological processes51, but also the well-known enhancement by a factor of 2.5 ~ 3.0 in response of cultured mammalian cells to ionizing radiation by the presence of oxygen at 10–100 μM (ref. 55).
In this study, we do not rule out the possibility that EGCG present in the cell might enhance the formation of O2− radical, which could cause some reductive damage to the DNA or be converted to less reactive H2O2. However, O2− radical has a short lifetime51, much shorter than the electron in EGCG (EGCG is quite stable in an aqueous solution under air), though both O2•− and EGCG have a similar redox potential. Thus, the toxicity of O2− must be much less than the reductive damage induced directly by EGCG. Moreover, H2O2 (generally believed to be far less toxic than O2−) once formed will rapidly be reduced with the presence of the strong reducing agent EGCG in the cell. Thus, it is reasonable to conclude that O2− radical if formed will have much less toxicity than EGCG in the cell and that the toxicity of H2O2 will be strongly quenched by the presence of EGCG in the cell, as observed previously that EGCG protects the H2O2 –induced cell death30,31,32.
For observed contrast differences in effects of EGCG on normal cells and abnormal (cancer) cells, they can be related to higher-level intrinsic antioxidants in abnormal cells, as recently noted by Watson22. Antioxidants can not only have a promotion effect in tumour initiation21, but reduce the effect of an exogenous reducing agent in killing tumor cells. As illustrated in Figure 8, the present results show that like epre− produced in ionizing radiation or O2− produced in cellular processes, weakly-bound electrons in antioxidants can cause serious reductive DNA damage. The DNA damage, if not repaired properly, leads to apoptosis, genetic mutations and likely diseases notably cancer. Thus, it is not surprising that abnormal (cancer) cells have a higher level of intrinsic antioxidants22. Simultaneously, abnormal (cancer) cells may activate protective mechanisms to balance the detrimental effects of chronically high antioxidant levels and therefore have some resistance to exogenous antioxidants especially at low levels (≤100 μM for EGCG observed in the present study). It is also observed that an exogenous reducing agent (EGCG/GT) at its extremely high concentrations (100–500 μM) can still make DNA damage and destroy the abnormal (tumor) cells via the reductive mechanism. However, it must be kept in mind that at such high concentrations of antioxidants, human normal cells can severely be damaged or killed, as also seen in this study. As a result, there seem no benefits to supplement EGCG to patients receiving chemotherapy.

A fresh understanding of reductive damage and diseases (cancer).
Intrinsic or exogenous reductive sources, such as antioxidants, ionizing radiation and O2− produced in cellular processes, can give rise to weakly-bound electrons, leading to reductive DNA damage, genomic mutations and ultimately diseases (cancer). Thus, abnormal cells such as cancer cells may have a higher level of intrinsic antioxidants. Simultaneously, abnormal (cancer) cells may activate protective mechanisms to balance the detrimental effects of chronically high antioxidant levels and therefore have some resistance to exogenous antioxidants especially at low levels.
Finally, we should note that there are certainly other chemical components and oxidants which may neutralize or lessen the toxicity of EGCG in the human body. However, it should also be noted that the present study did use two references, H2O2 as a major ROS inducing oxidative damage and cisplatin both as a highly toxic chemotherapeutic drug and as a well-known cancer-causing agent. And the results do show that cell killing and DNA DSBs by EGCG were far more severe than those by H2O2 and cisplatin under identical cell culture conditions. Moreover, the present results are in fact consistent with the observations of the clinical trials that the incidences of human lung cancer and skin cancer increased with antioxidant supplementation5,6,7,8,9,10. Thus, the present results are certainly of physiological and pathological significance. Although flavonoids as strong reducing agents in GT may stimulate degradation of high-level fats and proteins in the body and may therefore lead to a lower risk of developing heart disease, their carcinogenic potential cannot be ignored. Note that the deleterious health effects might not be observed immediately, as it is a slow process and may take over years or decades from DNA damage, genetic mutation to disease (cancer) development. In light of the high mean content of EGCG in a cup of GT and severe toxic effects against human normal cells observed in this study, GT drinkers should be very cautious, especially to avoid drinking the GT with GT leaves immerged with hot water for over minutes. The latter was in fact well recorded in the Chádào (the old Chinese art of making tea), but is often neglected by many people including Chinese in modern life.
In conclusion, our results provide a compelling explanation for the well-documented results of clinical trials that increases in lung and skin cancers were associated with antioxidants. We have clearly demonstrated the importance and the genotoxic and mutagenic effects of a previously unrecognized reductive damaging mechanism in cellular processes. Our results strongly suggest that cancer and many other human diseases are more likely caused by reductive damage than oxidative damage to DNA and the cell. This fresh fundamental understanding may be a key step for prevention and cure of challenging human diseases. The present results also show that natural antioxidant supplements can cause detrimental effects on healthy humans and may cause more cancers than they prevent cancers. This study may therefore change the human dietary style and improve the quality of life, health and longevity, as well as provide a possible answer to the question recently asked by James Watson22.
Methods
Chemicals and reagents
Ultrapure water for life science with a resistivity of >18.2 MΩ/cm and TOC<1 ppm obtained freshly from a Barnstead Nanopure water system was used. All of the chemicals and reagents [EGCG, cisplatin, dGMP, 6-Thioguanine(TG), etc.] were obtained from Sigma-Aldrich, while the green tea (GT) was obtained from a commercial source (supermarket). EGCG was prepared in pure water or ethanol (EtOH) at a stock concentration of 10 mM; cisplatin CDDP was stored at a concentration of 3 mM in pure water. The Vybrant 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) Cell Proliferation/viability Assay Kit and the HCS DNA Damage Kit (for DNA DSB measurements) were purchased from Invitrogen.
Preparation of green tea (GT) solution
Weighted 5.8 g of green tea (leaf) and put into a container; added 250 ml of boiling water into the container with a cover. After 20 min, 1, 2, 4, 6 and 8 hr naturally cooling at room temperature, took 4 ml of GT solution and used a syringe with 0.2 μm filter to remove the residues. Took about 20 μl of filtered GT solution and diluted it by 20 times, then used a spectrophotometer to measure the UV absorption spectrum for determination of the concentration of EGCG in the GT stock solution. We observed that after boiling water was added to the GT leaf for ≥4 hr, there was no significant further change in the absorption spectrum, indicating that most of EGCG had been dissolved in the GT solution (see Figure 1). The micro-filtered GT stock solution was then frozen and stored at −20°C for further experiments.
UV (electronic) absorption spectroscopic measurements
The spectra were measured using a spectrophotometer (Beckman Coulter, DU 530). UV absorption spectra of the mixtures of 50 μM dGMP with 50 μM EGCG/GT and 100 μM dGMP with 200–500 μM H2O2 at zero and 24 hr at 37°C, as well as their difference spectra were measured. The sample of the EGCG+ cation was prepared by reaction of 50 μM EGCG with 1 mM HCl for 1 hr; the UV absorption difference spectrum between 0 min and 60 min was obtained to show the absorption feature of the resultant EGCG+ (See Figure S1). The sample of the (G-H)−/G− anion was prepared by the reaction of dGMP with prehydrated electrons generated by femtosecond laser radiolysis of water with the presence of the OH radical scavenger (2 M isopropanol)14 (200 μW, 120 fs, 500 Hz, 5 hr). The resultant G*− anion either rapidly dissociates into a (G-H)•/(G-H)− radical and a H−/H• fragment or forms a low yield of the stabilized G− anion13. Since the neutral (G-H)• radical is well known to have a narrow absorption peak at ~315 nm and have a lifetime less than 1 s (ref. 56), it could not be observed in our present static UV absorption measurements. Thus, only the (G-H)− or G− anion could be detected in our shown UV (electronic) absorption spectra. The UV absorption difference spectrum of dGMP before and after the reaction (irradiation) was obtained to show the characteristic absorption spectrum of (G-H)− or G− anion (See Figure S2).
Cell lines and culture conditions
The two human lung diploid fibroblasts (WI-38 and MRC-5 cell lines), a human skin diploid fibroblast (GM05757 cell line) and a Chinese hamster ovary (CHO-K1: GM15452) cell line were obtained from the Coriell Cell Repository directly, while the lung cancer (A549 cell line), together with RPMI 1640 and F-12K culture media, was obtained from the American Type Culture Collection (ATCC) directly. Fetal bovine serum (FBS) was obtained from Hyclone Laboratories (UT, USA). The human normal cells were cultivated with MEM supplemented with 10–20% fetal bovine serum (FBS), 100 units/mL penicillin G and 100 μg/mL streptomycin (Hyclone). The complete growth medium for A549 cells was the ATCC-formulated F-12K medium with 10% FBS. CHO-K1 cells were grown in Hams F12 medium (from Sigma) containing 5% FBS, 100 units/mL penicillin G and 100 μg/mL streptomycin. The cells were incubated at 37°C in a humidified atmosphere containing 5% CO2.
Direct measurements of intracellular reductive DNA damage induced by EGCG
The normal cells were seeded at a density of 8 × 104 cells/well in 6-well plates and incubated for overnight. The cells were then treated with EGCG at different concentrations (0, 30 and 50 μM) in the culture medium. After the 24 h treatment, detached (dead) cells in each well of the same EGCG concentration were collected into a 50 ml tube while the adherent cells were trypsinized and then added to the 50 ml tube. The cells were then centrifuged for 5 minutes and the supernatant was removed. The EGCG-treated cells were subsequently washed with PBS for three times and then added into the PBS solution at a fixed density of 1.5 × 104 cells/well in a UV transmitted 96-well plate. The absorption spectrum at 300–600 nm of the cells in each well was measured by a MultiScan microplate reader (Thermo Scientific).
Cell viability assay
The effects of EGCG/GT/H2O2/CDDP and the combination effects of CDDP and EGCG/GT on cell viability were determined by the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay, one of the most commonly used cell viability assays. The details have been given previously41. Briefly, cells were plated at a density of 7 × 103 cells/well in 96-well plates. Following overnight incubation, cells were treated with different concentrations of EGCG/H2O2/CDDP or CDDP combined with EGCG for 24 h. After incubation for specified times at 37°C in a humidified incubator, 20 μL of MTT (5 mg/mL in PBS) were added to each well and cells were incubated for a further 4 h. After removal of the medium, 100 μL DMSO was added to each well. The absorbance was recorded on a microplate reader at the wavelength of 540 nm. The effects of the agent(s) on growth inhibition were assessed as percent cell viability where the untreated cells were taken as 100% viability.
Imaging of DNA double-strand breaks (DSBs) and toxicity
The phosphorylated H2AX foci are a biomarker of DNA DSB. The HCS DNA Damage Kit (Invitrogen) was developed to enable simultaneous quantitation of two cell health parameters, genotoxicity and (cyto-)toxicity. DNA DSBs are measured as an indication of genotoxicity and accomplished by specific antibody-based detection of phosphorylated H2AX (γH2AX) in the nucleus. Toxicity is measured with the Image-iT® DEAD Green™ viability stain, which has a high affinity for DNA, is non-fluorescent and impermeant but forms highly fluorescent and stable dye-nucleic acid complexes when bound to DNA. Staining of nuclear DNA cannot occur in live cells due to the impermeability of the plasma membrane for the stain. If test compounds lead to serious cell injuries, including plasma membrane permeability, the Image-iT® DEAD Green™ viability stain is allowed to enter the cells. This property enables the discrimination of dead cells with Image-iT® DEAD Green™ viability stain. In addition, the kit included Hoechst 33342, which is a DNA-binding dye (blue fluorescence) and allows the observation of nuclear morphology of all normal and damaged cells. Briefly, the cells were treated with different concentrations of EGCG/H2O2 for 6 or 12 h under normal cell culture conditions described above. Then following the detailed experimental procedures described in the Protocol provided by the manufacturer, we performed the HCS DNA Damage Assay of the treated cells. The images of cells were acquired with a Nikon Eclipse TS100 fluorescence microscope; quantitative analyses of activated γ-H2AX (DNA DBS yield) in the cells were performed using an Image J software.
DNA fragmentation (apoptosis) measurements by flow cytometry
One late stage marker of apoptosis is the degradation of DNA into small fragments. An enhancement in DNA fragmentation indicates increases in apoptosis. As per manufacturer's protocol, we detected DNA fragmentation using a standard APO-BrdU TUNEL assay (Invitrogen). BrdU-positive cells are apoptotic cells and the fluorescence intensity above the background (at the right side of the vertical line in the histograms) indicates the apoptotic cells with fragmented DNA. Because of a large number of cells required for flow cytometric analysis, our apoptosis measurements started with cells grown in T-25 flasks (unlike the cell viability measurements with cells seeded in 96-well plates) and then treated with EGCG for 24 h. After trypsinized, the cells were washed with PBS, processed for labeling with the deoxythymidine analog 5-bromo-2′-deoxyuridine 5′-triphosphate (BrdU-TP), incubated with an Alexa Fluor 488 dye–labeled anti-BrdU antibody and stained with propidium iodide (PI). The labeled cells were analyzed by flow cytometry. The experimental details have been published previously41.
HPRT gene mutation assay for CHO-K1 cells
The hypoxanthine phosphorybosyl transferase (HPRT) gene on the X chromosome of mammalian cells is widely used as a model gene to investigate gene mutations in mammalian cell lines45. The HPRT methodology detects mutations, which destroy the functionality of the HPRT gene and or/protein, by positive selection using a toxic analogue (6-TG) and cells with HPRT − mutants are seen as viable colonies. It detects a broad spectrum of mutagens, since any mutation resulting in the ablation of gene expression/function produces a HPRT − mutant. Chinese hamster ovary (CHO) cells have been the main cell line for HRPT assay; the detailed procedures can be found in the literature45,50. Our design of the HPRT assay tested four EGCG concentrations with duplicate treatments per concentration. For each experiment, CHO-K1 cells were seeded in 4 dishes (100 mm, 5 × 105 cells/dish) and incubated at 37°C in a humidified atmosphere containing 5% CO2 for 24 hrs. The cells in the dishes were then treated with various EGCG concentrations (0, 25, 50, 100 μM) in incubator for 24 hrs, where the dish without EGCG served as the control. The cells were then rinsed, trypsinized and re-seeded in triplicate in 60 mm dishes with 100 cells/dish for cytotoxicity assessment. The remaining cells were cultured in fresh medium in the same 100 mm dishes, including subcultures every 2–4 days, to initiate the post-treatment period for expression of the mutant phenotype. After 7 days, the medium in the 60 mm dishes was removed, the cells were washed with PBS, fixed and stained with a mixture of 6.0% glutaraldehyde and 0.5% crystal violet and then the number of colonies in each dish was counted. The plating efficiencies and clonogenic survival fractions of the cells treated with various EGCG concentrations were calculated. For determination of mutant frequencies, the cells in the 4 dishes (100 mm) after 7 days were reseeded at a density of 2 × 105 cells/dish (5 dishes for each EGCG dose) and cultured for mutant selection in the medium containing 10 μM 6-TG. For determination of cloning efficiencies, the cells were also re-seeded in 60 mm dishes (100 cells/dish) in triplicate and cultured in the medium without 6-TG and incubated under standard conditions. After another 7-day incubation, the numbers of colonies in the 100 mm and 60 mm dishes (averages of 5 or 3 dishes) were counted and the mutant frequencies and the cloning efficiencies were calculated.
Statistical analysis
The data are expressed as mean value ± SD and statistically analyzed with two-tailed and paired t-tests. A P value < 0.05 was considered statistically significant. In each cell viability experiment, the cells were repeatedly measured in 5 wells of the 96-well plate for the treatment of each concentration of EGCG/GT/H2O2/CDDP. All quantitative experiments were conducted with at least two independent experiments.
References
American Cancer Society. Cancer Facts and Figures 2013. (http://www.cancer.org/acs/groups/content/@epidemiologysurveilance/documents/document/acspc-036845.pdf (2013).
Lui, H., Zhao, J., McLean, D. & Zeng, H. Real-time Raman spectroscopy for in vivo skin cancer diagnosis. Cancer Res. 71, 2491–2500 (2012).
Bjelakovic, G., Nikolova, D., Gluud, L., Simonetti, R. & Gluud, C. Mortality in Randomized Trials of Antioxidant Supplements for Primary and Secondary Prevention: Systematic Review and Meta-analysis. JAMA 297, 842–857 (2007).
Bjelakovic, G., Nikolova, D., Gluud, L., Simonetti, R. & Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database of Systematic Reviews 2012, Issue 3. Art. No.: CD007176; 10.1002/14651858.CD007176.pub2 (2012).
The Alpha-Tocopherol, Beta Carotene Cancer Prevention Study Group. The effects of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 330, 1029–1035 (1994).
Albanes, D. et al. Alpha-tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: effects of base-line characteristics and study compliance. J. Natl. Cancer. Inst. 88, 1560–1570 (1996).
Omenn, G. S. et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy Trial. J. Natl. Cancer. Inst. 88, 1550–1559 (1996).
Omenn, G. S. et al. Effects of a combination of beta-carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 334, 1150–1155 (1996).
Albanes, D. Beta-carotene and lung cancer: a case study. Am. J. Clin. Nutr. 69, 1345S–1350S (1999).
Hercberg, S. et al. Antioxidant supplementation increases the risk of skin cancers in women but not in men. J. Nutr. 137, 2098–2105 (2007).
Lu, Q.-B. Cosmic-Ray-Driven Electron-Induced Reactions of Halogenated Molecules Adsorbed on Ice Surfaces: Implications for Atmospheric Ozone Depletion and Global Climate Change. Phys. Rep. 487, 141–167 (2010).
Lu, Q.-B. Effects of Ultrashort-Lived Prehydrated Electrons in Radiation Biology and Their Applications for Radiotherapy of Cancer. Mutat. Res.-Rev. Mutat. Res. 704, 190–199 (2010).
Wang, C.-R., Nguyen, J. & Lu, Q.-B. Bond Breaks of Nucleotides by Dissociative Electron Transfer of Nonequilibrium Prehydrated Electrons: A New Molecular Mechanism for Reductive DNA Damage. J. Am. Chem. Soc. 131, 11320–11322 (2009).
Nguyen, J. et al. Direct Observation of Ultrafast Electron Transfer Reactions Unravels High Effectiveness of Reductive DNA Damage. Proc. Natl. Acad. Sci. USA 108, 11778–11783 (2011).
Lu, Q.-B. Molecular Reaction Mechanisms of Combination Treatments of Low-Dose Cisplatin with Radiotherapy and Photodynamic Therapy. J. Med. Chem. 50, 2601–2604 (2007).
Wang, C. R. & Lu, Q.-B. Real-Time Observation of Molecular Reaction Mechanism of Aqueous 5-halo-2′-deoxyurimidines under UV/Ionizing Radiation. Angew. Chem. Intl. Ed. 46, 6316–6321 (2007).
Wang, C. R. & Lu, Q.-B. Molecular Mechanism of the DNA Sequence Selectivity of 5-Halo-2′-Deoxyuridines as Potential Radiosensitizers. J. Am. Chem. Soc. 132, 14710–14712 (2010).
Bristow, R. G. & Hill, R. P. Hypoxia, DNA Repair and Genetic Instability. Nat. Rev. Cancer 8, 180–192 (2008).
Rhee, S. G. CELL SIGNALING: H2O2, a Necessary Evil for Cell Signaling. Science 312, 1882–1883 (2006).
Perera, R. M. & Bardeesy, N. When antioxidants are bad. Nature 475, 43–44 (2011).
DeNicola, G. M. et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 475, 106–109 (2011).
Watson, J. Oxidants, antioxidants and the current incurability of metastatic cancers. Open Biol. 3, 120144 (2013).
UN Food and Agriculture Organization, Intergovernmental Group on Tea. Current situation and medium-term outlook. 9 (2008).
Qiao, Y., Cao, J., Xie, L. & Shi, X. Cell growth inhibition and gene expression regulation by (−)-epigallocatechin-3-gallate in human cervical cancer cells. Arch. Pharm. Res. 32, 1309–1315 (2009).
Philips, B. J., Coyle, C. H., Morrisroe, S. N., Chancellor, M. B. & Yoshimura, N. Induction of apoptosis in human bladder cancer cells by green tea catechins. Biomed. Res. 30, 207–215 (2009).
Hsieh, T.-C. & Wu, J. M. Targeting CWR22Rv1 Prostate Cancer Cell Proliferation and Gene Expression by Combinations of the Phytochemicals EGCG, Genistein and Quercetin. Anticancer Res. 29, 4025–4032 (2009).
Yang, G.-Y., Liao, J., Kim, K., Yurkow, E. J. & Yang, C. S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 19, 611–616 (1998).
Elbling, L. et al. Hydrogen peroxide mediates EGCG-induced antioxidant protection in human keratinocytes. Free Radical Bio. Med. 49, 1444–1452 (2010).
Nakagawa, H., Hasumi, K., Woo, J.-T., Nagai, K. & Wachi, M. Generation of hydrogen peroxide primarily contributes to the induction of Fe(II)-dependent apoptosis in Jurkat cells by (−)-epigallocatechin gallate. Carcinogenesis 25, 1567–1574 (2004).
Tobi, S. E., Gilbert, M., Paul, N. & Mcmillan, T. J. The Green Tea Polyphenol, Epigallocatechin-3-Gallate, Protects against the Oxidative Cellular and Genotoxic Damage of UVA Radiation. Int. J. Cancer 102, 439–444 (2002).
Saffari, Y. & Sadrzadeh, S. M. H. Green tea metabolite EGCG protects membranes against oxidative damage in vitro. Life Sci. 74, 1513–1518 (2004).
Thichanpiang, P., Khanobdee, K., Kitiyanant, Y. & Wongprasert, K. Green Tea epigallocatechin-3-gallate Protects Against Oxidative Stress-Induced Nuclear Translocation of p53 and Apoptosis in Retinal Pigment Epithelial Cells, ARPE-19. J. Agri. Sci. 5, No. 4 (2013).
Yamamoto, T. et al. Roles of Catalase and Hydrogen Peroxide in Green Tea Polyphenol-Induced Chemopreventive Effects. J. Pharmaco. Exp. Therapeutics (JPET) 308, 317–323 (2004).
Strick, R., Strissel, P. L., Borgers, S., Smith, S. L. & Rowley, J. D. Dietary bioflavonoids induce cleavage in the MLL gene and may contribute to infant leukemia. Proc. Natl. Acad. Sci. USA 97, 4790–4795 (2000).
Paolini, M., Sapone, A. & Valgimigli, L. Avoidance of bioflavonoid supplements during pregnancy: a pathway to infant leukemia? Mutat. Res. 527, 99–101 (2003).
Plichart, M. et al. Parental smoking, maternal alcohol, coffee and tea consumption during pregnancy and childhood malignant central nervous system tumours: the ESCALE study (SFCE). Eur. J. Cancer. Prev. 17, 376–383 (2008).
Kulling, S. E. & Metzler, M. Induction of Micronuclei, DNA Strand Breaks and HPRT Mutations in Cultured Chinese Hamster V79 Cells by the Phytoestrogen Coumoestrol. Food Chem. Toxicol. 35, 605–613 (1997).
Stopper, H., Schmitt, E. & Kobras, K. Genotoxicity of phytoestrogens. Mutat. Res. 574, 139–155 (2005).
Gatz, S. A. & Wiesmüller, L. Take a break–resveratrol in action on DNA. Carcinogenesis 29, 321–332 (2008).
Fox, J. T. et al. High-throughput genotoxicity assay identifies antioxidants as inducers of DNA damage response and cell death. Proc. Natl. Acad. Sci. USA 109, 5423–5428 (2012).
Luo, T. et al. Electron transfer-based combination therapy of cisplatin with tetramethyl-p-phenylenediamine for ovarian, cervical and lung cancers. Proc. Natl. Acad. Sci. USA 109, 10175–10180 (2012).
Jovanavic, S. V., Steenken, S. & Simic, M. G. Reduction potentials of flavonoid and model phenoxyl radicals. J. Chem. Soc. Perkins Trans. 2, 2497–503 (1996).
Jovanavic, S. V., Hara, Y., Steenken, S. & Simic, M. G. Antioxidant potential of theaflavins. A pulse radiolysis study. J. Am. Chem. Soc. 119, 5337–5343 (1997).
Kleinová, M. et al. Antioxidant properties of carotenoids: QSAR prediction of their redox potentials. Gen. Physiol. Biophys. 26, 97–103 (2007).
Johnson, G. E. Chapter 4. Mammalian Cell HPRT Gene Mutation Assay: Test Methods. In: Genetic Toxicology: Principles and Methods, Methods in Molecular Biology, vol. 817, [Parry, J. M. & Parry, E. M. (ed)] [55–67] (Springer Science + Business Media, LLC, 2012).
Naumov, S. & von Sonntag, C. Guanine-Derived Radicals: Dielectric Constant-Dependent Stability and UV/Vis Spectral Properties: A DFT Study. Radiat. Res. 169, 364–372 (2008).
Faraggi, M. & Klapper, M. H. One electron oxidation of guanine and 2-deoxyguanosine by the azide radical in alkaline solutions. J. Chim. Phys. 91, 1062–1069 (1994).
Culp, S. J., Cho, B. P., Kadlubar, F. F. & Evans, F. E. Structural and Conformational Analyses of 8-Hydroxy-2′-deoxyguanosine. Chem. Res. Toxicol. 2, 416–422 (1989).
Choi, S. Mechanism of Two-Electron Oxidation of Deoxyguanosine 5-Monophosphate by a Platinum (IV) Complex. J. Am. Chem. Soc. 126, 591–598 (2004).
Slesinski, R. S., Clarke, J. J., San, R. H. & Gudi, R. Lack of mutagenicity of chrominum picolinate in the hypoxanthine phosphoribosyltransferase gene mutation assay in Chinese hamster ovary cells. Mutat. Res. 585, 86–95 (2005).
Fridovich, I. Superoxide Anion Radical (O•−2), Superoxide Dismutases and Related Matters. J. Bio. Chem. 272, 18515–18517 (1997).
Prousek, J. Fenton chemistry in biology and medicine. Pure Appl. Chem. 79, 2325–2338 (2007).
CRC Handbook of Chemistry and Physics, 93rd Ed. [Lide, D. R. (ed)] [10–149] (CRC Press, 2003).
Bielski, B. H. J., Cabelli, D. E., Arudi, R. L. & Ross, A. B. Reactivity of HO2/O2− Radicals in Aqueous Solution. J. Phys. Chem. Ref. Data 14, 1041–1100 (1985).
Lehnert, S. Biomolecular Action of Ionizing Radiation. [406] (Taylor & Francis Ltd., New York, 2007).
Shafirovich, V., Dourandin, A., Huang, W. & Geacintov, N. E. The Carbonate Radical Is a Site-selective Oxidizing Agent of Guanine in Double-stranded Oligonucleotides. J. Bio. Chem. 276, 24621–24626 (2001).
Acknowledgements
This work was supported in part by the Canadian Institutes of Health Research (a New Investigator Award to Q.B.L), the Ontario Ministry of Research and Innovation (an Early Researcher Award to Q.B.L) and Natural Science and Engineering Research Council of Canada (NSERC, a grant to Q.B.L).
Author information
Authors and Affiliations
Contributions
L.Y.L., N.O. and Q.B.L. carried out the experiments; L.Y.L. and Q.B.L. wrote the main manuscript text; and L.Y.L. and N.O. prepared all the figures. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Lu, L., Ou, N. & Lu, QB. Antioxidant Induces DNA Damage, Cell Death and Mutagenicity in Human Lung and Skin Normal Cells. Sci Rep 3, 3169 (2013). https://doi.org/10.1038/srep03169
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03169
This article is cited by
-
New combination chemotherapy of cisplatin with an electron-donating compound for treatment of multiple cancers
Scientific Reports (2021)
-
High doses of synthetic antioxidants induce premature senescence in cultivated mesenchymal stem cells
Scientific Reports (2019)
-
Antioxidant and anticancer activities of Trigonella foenum-graecum, Cassia acutifolia and Rhazya stricta
BMC Complementary and Alternative Medicine (2018)
-
Kynurenic Acid Restores Nrf2 Levels and Prevents Quinolinic Acid-Induced Toxicity in Rat Striatal Slices
Molecular Neurobiology (2018)
-
Antigenotoxic properties of Paliurus spina-christi Mill fruits and their active compounds
BMC Complementary and Alternative Medicine (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
