
Abstract
The aim of this study was to analyse the relationships between polymorphisms in four candidate genes (AR, DAT1, DRD2 and COMT) and aggression in men from a traditional society of East African pastoralists, the Datoga. Buss and Perry's Aggression Questionnaire was used to measure aggression. The number of CAG repeats in the AR gene was negatively correlated with physical aggression, anger and hostility. Among the genes of the dopaminergic system, a significant single-gene effect was detected only for DRD2 with regard to anger. At the level of a two-gene model, a significant effect for DRD2 and a tendency for DAT1 were observed for the DAT1-DRD2 gene pair regarding hostility and two tendencies were observed for the interaction effect of the DAT1-COMT pair regarding anger and hostility. These data suggest a probable link between physical aggression and direct fitness caused by strong sexual selection in Datoga men.
Similar content being viewed by others


Introduction
Aggressive behaviour in humans is highly sexually dimorphic, with men exhibiting more violent (physical) same-sex aggression than women in almost all cultures1,2,3,4, which is most likely attributable to the greater impulsiveness of men and a stronger fear of physical danger in women5,6. A meta-analysis of 24 genetic studies on aggression demonstrated that heritability accounts for approximately 50% of the variance in aggression7,8, with the heritability of aggressive behaviour being higher in men than in women9. Aggressive behaviour (physical and verbal) and its negative emotional bases (anger and hostility) are the products of complex interactions between many genes associated with the secretion of steroid hormones and the sensitivity of tissues to these hormones. These genes include those associated with the dopaminergic and serotonergic systems (both receptors and transmitters) and likely many others10,11,12.
The results of some studies suggest a relationship between exposure to testosterone in utero and subsequent aggression in adulthood as a result of masculinisation of the brain13,14. Individual behavioural reactions to testosterone have also shown a close dependence on the sensitivity of brain tissue to this steroid. Androgen receptor genes are located on the X chromosome and are therefore only inherited by men from their mothers. Earlier studies, predominantly conducted among representatives of industrial populations, appear to support the hypothesis of a negative correlation between an increasing the number of CAG repeats in the androgen receptor gene (AR) and physical aggression and/or the 2D:4D ratio (the ratio between the lengths of the second and fourth digits, an indicator of the masculinisation of the foetus in utero)15. Another study reported that fewer CAG repeats in AR were more frequently found in male rapists and murderers than in control subjects16. Indeed, an increased number of these repeats in AR results in receptors with lower androgen sensitivity17,18. Manning and colleagues demonstrated in a male sample from an industrial population that masculine finger ratios are associated with androgen receptor alleles with fewer CAG microsatellite repeats in the terminal domain15. In contrast, a recent study performed by our group in traditional hunter–gatherers, the Hadza of Tanzania, revealed no correlation between the number of CAG repeats and aggression or the right-hand 2D:4D ratio in Hadza men19. Thus, the recent data on the associations between AR polymorphisms and aggression are somewhat contradictory and demand a more thorough investigation in other small-scale societies.
Many recent molecular genetic studies on aggression have focused on the genes involved in dopamine neurotransmission. The distributions of polymorphic loci in the dopamine receptor genes DRD4 and DRD2 and the dopamine transporter gene DAT1 were analysed to identify an association between specific alleles and aggressive behaviour (particularly obsessive–compulsive disorder, hostility and physical aggression), some mental disorders and alcohol addiction20,21,22,23,24,25,26,27. Another gene associated with aggression is the gene encoding catechol-O-methyltransferase (COMT), which plays a key role in dopamine catabolism in the prefrontal cortex: the V158M polymorphism in the fourth exon of the COMT gene leads to a 40% reduction of enzymatic activity in carriers of the M allele28.
Dopamine plays a critical role in reward processing and is a potent neuromodulator of ventral striatum (VS) reactivity, which is known to be highly involved in reward processing. VS reward reactivity may be a key neurobiological pathway through which DAT1 and DRD2 polymorphisms contribute to the variability of behavioural impulsivity29.
DAT1 and DRD2 polymorphisms were examined in adolescents with pathological aggression and it was found that carriers of the DAT1 10/10 genotype were overrepresented in a sample of aggressors compared to the control group and that DRD2 A1/A1 and A1/A2 were also overrepresented in aggressors30. Vaughn and colleagues31 suggested DAT1 as one of the main genetic markers of antisocial behaviour in adolescent criminals. According to another study, the DAT1 10/10 and 10/9 genotypes are present in violently delinquent young adults twice as often as in controls26,32,33,34. Guo and co-authors26 demonstrated that heterozygotes for the DRD2 A1/A2 variant presented higher aggression scores than A1/A1 or A2/A2 homozygotes; these authors further suggested that there is no correlation or epistasis between DRD2 and DAT1.
Remarkably, the data from a nationally representative sample of American youth revealed that genetic risk factors measured based on two dopamine polymorphisms (DAT1 and DRD2) confound the association between paternal incarceration and a child's arrest35.
A serious problem with the current studies on the molecular genetics of aggression and antisocial behaviour is that such research frequently cannot be replicated36. Moreover, a recently conducted meta-analysis of genetic association studies on violence and aggression provided a rather pessimistic outlook, indicating that the candidate gene approach has not succeeded in identifying genes associated with these outcomes37. One of the general explanations for this difficulty is that “there are few, if any, loci with large effect size and it is becoming increasingly obvious that it will be necessary to consider the impact of genes, not in isolation, but as part of a multifactorial miasma including both other genetic factors and the environment”9. Because most studies are based on clinical samples or samples that are not nationally representative and each sample is likely differentially exposed to particular environmental conditions, certain genes may only have effects when they are paired with certain environments38. Another possible reason for the lack of reproducibility in this arena is population stratification. Indeed, the allelic frequencies of some genes vary significantly across different racial/ethnic categories39 and genetic effects may vary across racial/ethnic groups because of (a) differences in allelic distributions and (b) differences in exposure to environmental risk factors38.
Thus, there is an urgent need to examine the associations between certain genes and aggressive behaviour in samples of various racial/ethnic groups. In this study, we examined the single-gene effects of the AR gene and three dopaminergic genes (DAT1, DRD2 and COMT) as well as the two-gene associations of components of the dopaminergic system with aggression in a sample of Datoga, who are traditional East African pastoralists.
The following hypotheses were tested:
-
1
Men showing lower AR CAG repeat numbers exhibit higher aggression scores, particularly for physical aggression and negative emotions.
-
2
Male carriers of the +10 type of DAT1 (homozygous or heterozygous) present higher aggression scores.
-
3
Men heterozygous for DRD2 display higher aggression scores compared to both types of homozygotes.
-
4
Men homozygous for the high-activity Val/Val genotype of COMT show higher aggression scores.
-
5
Two-gene interaction effects between the three genes in the dopaminergic system (DAT1, DRD2 and COMT) provide important information on the associations of these genes with various types of aggression.
Results
The means and standard deviations for Buss and Perry's Aggression Questionnaire subscales were computed as well as minimum and maximum scores for Datoga males (Table 1).
We performed a simple regression analysis to test the single-gene effect of the AR gene on aggression and detected significant effects of the AR CAG polymorphism on physical aggression and hostility, with carriers of smaller numbers of CAG repeats showing a higher rating (Tables 2 and Figs. 1a,b,c). The results for AR and the self-rated scores for anger exhibited a trend in the same direction (Table 2). Therefore, our data show that men with fewer CAG repeats behave more aggressively than their fellow males with a greater number of AR CAG repeats.

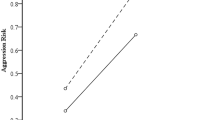
The distributions of the physical aggression (A), anger (B) and hostility (C) subscales as correlated with the number of CAG repeats in the AR gene.
The ordinate numbers designate the self-rated scores according to the subscales of Buss and Perry's Aggression Questionnaire.
We split our sample according to DAT1 genotype, as suggested previously by a number of authors26,30,32. The first group included men with the 9/9 genotype of the DAT1 (−10) type and the second group included individuals with the 9/10 or 10/10 genotype of the DAT1 (+10) type. One-way ANOVA with DAT1 type as the independent variable and aggression traits as the dependent variables revealed no significant association between the +10 type and all AQ scales (Table 3).
Our sample was also split into three groups based on DRD2 polymorphisms, with group 1 including men with the A1/A1 genotype, group 2 consisting of men with the A1/A2 genotype and group 3 comprising individuals with the A2/A2 genotype, following the divisions originally employed in a previous study by Guo and colleagues26. Significant single-gene effects of DRD2 in the Datoga men were found only with regard to anger (Fig. 2). Lastly, we divided our sample into three groups according to COMT polymorphisms: group 1 (V/V), group 2 (V/M) and group 3 (M/M); however, no single-gene effect of COMT was identified in this study (Table 3).

The distribution of the anger subscale as correlated with the DRD2 type (1 – A1/A1 and 2 – A1/A2 and A2/A2).
The designations of the numbers are described in Figure 1.
We subsequently performed a factorial two-way ANOVA to test both the main and interaction effects of the three genes (DAT1, DRD2 and COMT) on three aggression subscales (Tables 4). The two-gene model revealed two significant main effects of the DRD2 gene on anger and hostility, showing a tendency for a main effect of DAT1 on hostility. Tendencies of DAT1 × COMT interaction effects on anger and hostility, but not on physical aggression, were also observed (Fig. 3a,b).

The distribution of the anger (A) and hostility (B) as correlated with the DAT1 × COMT effect.
The designations of the ordinate numbers are described in Figure 1.
Discussion
Overall, we found that the polymorphisms in all four genes tested in this study affected the self-rated aggression scores of the Datoga men, with either single-gene effects or interaction effects between these genes being detected. Our data also confirmed our initial hypothesis regarding the single-gene effect of AR on the self-rated scores for the three AQ scales. Our results regarding the association between aggression and the number of CAG repeats in AR were in the predicted direction: the levels of physical aggression, anger and hostility among the Datoga men were higher in individuals with fewer CAG repeats. Interestingly, we recently demonstrated that individuals with a small number of CAG repeats are overrepresented among world-class judo sportsmen compared to the general Russian population40. At the same time, these data contradict those obtained by Hurd, Vaillancourt and Dinsdale41 in Canadian students, where men with more sensitive androgen receptors tended to score lower for physical aggression and anger. The results also contradict our early findings in Hadza men, for whom no correlations were found between the AQ subscales and the number of AR CAG repeats42. A reasonable explanation for these differences may be the existence of gene–environment effects and may be particularly attributable to differences in cultural attitudes toward aggression among the Hadza and Datoga populations.
The significant single-gene effects of DAT1 polymorphisms on anger and hostility were in the direction predicted in the second hypothesis. Therefore, not only is +10 DAT1 overrepresented in samples of antisocial adolescents in the USA31, but it is possible that these genotypes are positively selected in pastoralist cultures because such personalities are better suited to both cattle raiding and cattle protection. Although significant single-gene effects of the DRD2 polymorphism on anger and hostility were also demonstrated, contrary to our expectations, the carriers of the heterozygous type displayed the same high levels of these characteristics as the homozygous A2/A2 men. We did not find any single-gene effect of COMT in this study; therefore, our fourth hypothesis should be rejected. Lastly, we demonstrated tendencies toward interaction effects between DAT1 and COMT with regard to anger and hostility in Datoga males. According to our findings, these interaction effects are only essential for carriers of the different COMT genotypes and the –10 DAT1 type.
Guo and colleagues26 demonstrated that an interaction between DAT1 and DRD2 produces essentially the same effect on aggression observed in single-gene models, which was confirmed in our analysis of anger and hostility.
However, we did not test putative associations of AR gene polymorphisms with the three genes involved in the dopaminergic system, as there is still no evidence of their functional relationship.
Because these data were collected in a single population and due to the highly similar cultural and social environments of the respondents, we believe that we were able to minimise the inter-individual variation in the environmental influence. Moreover, only males were included in our analysis, as there is convincing evidence to support significant sex-based differences in gene–environment interactions43. The Datoga are polygynous pastoralists and the trajectories for achieving higher reproductive success are radically different in men and women. In contrast to the egalitarian and relatively peaceful Hadza hunter–gatherers, the men of the Datoga, another indigenous culture in the same region44,45, have been selected for their aggression, particularly their physical aggression, as have other African pastoralists46,47. Indeed, men with fierce characters and warrior skills may be more successful in their mating and parental efforts because they can acquire and protect cattle, consequently showing higher reproductive success (more wives and children)48. Furthermore, our data suggest a possible link between physical aggression and direct fitness caused by strong sexual selection in Datoga men, while such a relationship most likely no longer exists in Westernised societies.
Of course, the present work has a number of limitations. First, the sample size was not large. However, because all our respondents were from the same tribal community, practiced traditional rites, participated in the traditional economy and lived in the same area, the inter-individual variability in this respect was somewhat limited. As mentioned above, previous findings regarding the association between genes and particular behavioural traits are highly contradictory36,37. One possible reason for these disparities may be rooted in environmental factors38 and the present data together with our data on the Hadza42 illustrate a situation favouring this suggestion. Additionally, particular polymorphic loci may account for a relatively small proportion of phenotypic variance and these effects may not be significant in relatively small samples of individuals with different socialisation experiences. If more than two genes/polymorphisms and many behavioural traits are selected for analysis, the necessity of making corrections for multiple comparisons is problematic. One possible solution for this situation is to obtain a very large sample; another possibility is the generation of biologically founded multilocus genetic profiles reflecting the cumulative effects of multiple polymorphic loci of known functionality on a specific behaviour49. In accord with this approach, Beaver and Chavian created the Genetic Risk Index for criminal involvement in Hispanics based on three dopaminergic gene polymorphisms and demonstrated its effectiveness38. Other authors proposed the construction of a multilocus genetic profile score based on five functional polymorphic loci for dopamine signalling and reported that it accounted for 10.9% of the inter-individual variability in reward-related VS reactivity50. The dopamine (DA) index was found to be associated with ventral striatum activity in an experimental study and was recently applied in a psychiatric study. However, we decided not to use this index in our work because we do not know how it may be associated with the activity of other brain structures that are activated during aggression and their control in humans, particularly the prefrontal zone51. We refrained from applying such an approach in this study because (i) the initial assumption that particular polymorphisms act additively and not interactively to influence dopamine signalling should be tested in more detail and (ii) more evidence is needed that all of the loci used to generate the multilocus genetic profile score are truly invested equally in behaviour.
Methods
Ethics statement
Institutional approvals, including those from the University (Moscow State University Ethics Committee) and local governmental agencies (including the Tanzanian Commission for Science and Technology), were obtained prior to conducting this study. All subjects gave their informed, verbal consent prior to participation. Verbal consent was deemed appropriate given the low literacy rates among traditional Datoga and was specifically approved by the University EC and Tanzanian agency.
Study sample and data collection
The data were collected during field studies from 2006 to 2012 in the United Republic of Tanzania. The sample analysed here includes 138 adult Datoga men, with a mean age of 34.09 ± 12.12 years (median of 33.5 years; range, 18–70 years).
The participants were personally interviewed to determine their age, sex, ethnicity and personal history, as described previously42. All participants gave their consent prior to participating and the average pay per subject was equal to US$2. All interviews were conducted in a one-to-one dialogue with the respondents. All questions were read aloud and explained if necessary. Many of the Datoga men commented in these interviews that aggression is an appropriate way of ‘saving face’, for both themselves and their relatives and of protecting their property. When asked about family violence, all the men mentioned that they were physically punished during childhood by their parents (father, mother, or both). Self-rated aggression scores were obtained using Buss and Perry's Aggression Questionnaire (AQ)52, which consists of 29 statements, grouped into four subscales: physical aggression (nine items), verbal aggression (five items), anger (seven items) and hostility (eight items). The first two subscales, physical and verbal aggression, represent direct aggression and the other two subscales pertain to the individual's emotional basis for aggressive behaviour. AQ uses a Likert scale, ranging from 1 (extremely uncharacteristic) to 5 (extremely characteristic). This tool was employed because the original English version of the AQ has shown moderate construct validity and high test–retest reliability53 and has been widely applied in aggression studies. In the present work, we used the ki-Swahili version of the AQ, which was initially administered by us and was previously used in our research on the Hadza42. For the details of the data collection procedure, see Butovskaya et al42. Here, we present data for three subscales of the AQ: physical aggression, anger and hostility. Cronbach's Alpha values for these behavioural measures were as follows: physical aggression, 0.64; anger, 0.63; and hostility, 0.66.
Although the main conclusions drawn in this work regarding the subjects' aggression profiles were based on self-reported scores, rather than on direct observations, we do not think that this method of data collection significantly distorted the results. A meta-analysis performed earlier by Book and colleagues54 demonstrated that “the measure of aggression (behavioural or self-report) did not have any effect on the relationship between testosterone and aggression,” and there is no reason to assume that this situation differs with regard to genetic data.
Genotyping
Buccal epithelium samples were collected for DNA analysis during the field surveys and DNA was later isolated using the DIAtom™ DNA Prep 200 extraction kit (IsoGene Lab, Moscow, Russia), which is intended for the isolation of DNA from various biological materials, according to the manufacturer's protocol.
AR CAG repeats, DRD2 Taq1A, the DAT1 VNTR 3′-noncoding region and COMT Val-158-Met loci polymorphisms were typed via polymerase chain reaction (PCR), which was performed with the GenePak® PCR MasterMix Core (IsoGene Lab, Moscow, Russia) reagent kit, according to the recommendations of the manufacturer55.
Amplification of the target loci was performed using an MJ Research PTC-100 thermocycler with the following program: 1 cycle of 1 min at 94°C for denaturation, 30 sec at X°C for annealing and 30 sec at 72°C for synthesis. This was followed by 30 cycles of 30 sec at 94°C, 30 sec at X°C and 15 sec at 72°C, with a final elongation at 72°C for 3 min. X denotes the annealing temperatures of individual primer pairs (Table 5).
The reaction products for the AR locus were analysed using an ABI PRISM 3100-Avant automated DNA sequencer (Applied Biosystems, Foster City, USA).
To identify the TaqI polymorphism (T/C), the DRD2 amplification product was subdivided into 10 μl aliquots, one of which was treated with the TaqI restriction endonuclease overnight at 65°C. For the COMT locus, the PCR products were also subdivided into 10 μl aliquots and one of these aliquots was treated with NlaIII overnight at 37°C.
The amplification and restriction products were separated via electrophoresis in a 2% (DRD2 and DAT1) or 3% (COMT) agarose gel and stained with ethidium bromide. The results were photographed and analysed with a BioDocAnalyze device (Biometra, Goettingen, Germany).
Statistics
Statistical treatment of the data was performed with SPSS 13.0 for Windows (IBM, Armonk, NY, USA). The means and standard deviations were computed for the studied traits (Table 1). The Kolmogorov–Smirnov test was employed to test whether our behavioural data were normally distributed. As all of the obtained probability values were greater than 0.1, parametric methods of statistical analysis were suitable for evaluating the dependence of the aggression subscales on the genotypes. The scores at all four aggression subscales were positively correlated with one another (with r values ranging from 0.386 to 0.497); thus, we used the correction for the critical p value for multiple correlated outcomes (http://gump.qimr.edu.au/general/daleN/matSpD/) previously applied by Hurd, Vaillancourt and Dinsdale41. The adjusted critical p value for all of the aggression subscales was 0.01695. Because we tested the association of the AR gene and a set of three dopaminergic genes on aggression scales separately, the following corrections of the critical p-values were suggested. The adjusted critical value for AR was set to p ≤ 0.017/1(gene) = 0.017 (two-tailed), whereas p ≤ 0.034 was regarded as a trend. The significance levels for the three dopaminergic genes were calculated as p ≤ 0.017/3(genes) = 0.0057 (two-tailed); a level of probability ranging from 0.0057 to 0.0113 was considered a tendency.
A linear regression was applied to analyse the associations of the three aggression subscales with the number of CAG repeats in the AR gene. One-way and two-way ANOVAs were conducted to determine the effects of the three candidate dopamine genes on the variations in the examined behavioural traits. We did not test the interaction effects of AR and the three dopamine genes in association with aggression because they represent different functional systems.
A genetic population analysis revealed agreement with Hardy-Weinberg equilibrium for the genotypes of the DAT1 and COMT genes (Hobs = 0.505/Hexp = 0.505, p = 0.835 and Hobs = 0.491/Hexp = 0.443, p = 0.298, respectively), but not for the DRD2 gene (Hobs = 0.692/Hexp = 0.495, p ≪ 0.001, with a corrected level of significance of α/3 – 0.05/3 = 0.017).
References
Archer, J. Sex differences in aggression in real-world settings: a meta-analysis review. Rev Gen Psychol 8, 291–322 (2004).
Bjorkqvist, K. Sex differences in physical, verbal and indirect aggression: A review of recent research. Sex Roles 30, 177–188 (1994).
Butovskaya, M. L., Timentschik, V. & Burkova, V. Aggression, conflict resolution, popularity and attitude to school in Russian adolescents. Aggressive Behav 33, 170–183 (2007).
Butovskaya, M. L., Burkova, V. N. & Mabulla, A. Sex differences in 2D:4D ratio, aggression and conflict resolution in African children and adolescents: a cross-cultural study. J Aggress Confl Peace Res 2, 17–31 (2010).
Daly, M. & Wilson, M. Risk-Taking, Intrasexual Competition and Homicide French, J. A., Kamil, A. C. & Leger, D. W. (eds.) Volume 47 of the Nebraska Symposium on Motivation: Evolutionary Psychology and Motivation, 1–36 (Lincoln, NE: University of Nebraska Press, 2001).
Knight, G. P., Guthrie, I. K., Page, M. C. & Fabes, R. A. Emotional arousal and gender differences in aggression: A meta-analysis. Aggressive Behav 28, 366–393 (2002).
Miles, D. R. & Carey, G. Genetic and environmental architecture of human aggression. J Pers Soc Psychol 72, 207–217 (1997).
Rhee, S. H. & Waldman, I. D. Genetic and environmental influences on antisocial behavior: a meta-analysis of twin and adoption studies. Psychol Bull 128, 490–529 (2002).
Craig, I. W. & Halton, K. E. Genetics of human aggressive behaviour. Hum Genet 126, 101–113 (2009).
van Gestel, S. et al. Epistatic effect of genes from the dopamine and serotonin systems on the temperament traits of Novelty Seeking and Harm Avoidance. Mol Psychiatr 7, 448–450 (2002).
Trainor, B. C., Sisk, C. L. & Nelson, R. J. Hormones and the Development and Expression of Aggressive Behavior Pfaff, D. W., Arnold, A. P., Etgen, A. M., Fahrbach, S. E. & Rubin, R. T. (eds.) Volume 1 of the Hormones, Brain and Behavior (2nd ed.), 167–203 (San Diego, USA: Academic Press, 2009).
Vasilyev, V. A. Molecular psychogenetics of deviant aggressive behavior in humans. Russ J Genet 47, 1023–1032 (2011).
Cohen-Bendahan, C. C., van de Beek, C. & Berenbaum, S. A. Prenatal sex hormone effects on child and adult sex-typed behavior: methods and findings. Neurosci Biobehav R 29, 353–384 (2005).
Ryan, B. C. & Vandenbergh, J. G. Intrauterine position effects. Neurosci Biobehav R 26, 665–678 (2002).
Manning, J. T., Henzi, P., Venkatramana, P., Martin, S. & Singh, D. Second to fourth digit ratio: ethnic differences and family size in English, Indian and South African populations. Ann Hum Biol 30, 579–588 (2003).
Rajender, S. et al. Reduced CAG repeats length in androgen receptor gene is associated with violent criminal behavior. Int J Legal Med 122, 367–373 (2008).
Chamberlain, N. L., Driver, E. D. & Miesfeld, R. L. The length and location of CAG trinucleotide repeats in the androgen receptor N-terminal domain affect transactional function. Nucleic Acids Res 22, 3181–3186 (1994).
Kazemi-Esfarjani, P., Trifiro, M. A. & Pinski, L. Evidence for a repressive function of the long polyglutamine tract in the human androgen receptor: possible pathogenic relevance for the (CAG)n-expanded neuropathies. Hum Mol Genet 4, 523–527 (1995).
Butovskaya, M. L. Wife battering and traditional methods of it's control in contemporary Datoga pastoralists of Tanzania. J Aggress Confl Peace Res 4, 28–45 (2012).
Alfimova, M. V. & Trubnikov, V. I. Psychogenetics of aggression. Vop Psikhol 6, 112–122 (2000).
Kim, Y.-K. Sexual Selection and Aggressive Behavior in Drosophila Kim, Y. K. (ed.) Handbook of Behavior Genetics, 317–330 (New York: Springer Verlag, 2009).
Gelernter, J., Kranzler, H. & Cubells, J. Serotonin transporter protein (SLC6A4) allele and haplotype frequencies and linkage disequilibria in African– and European–American and Japanese populations and in alcohol dependent subject. Hum Genet 101, 234–246 (1997).
Hu, X.-Zh. et al. Serotonin Transporter promoter gain of function genotypes are linked to Obsessive-Compulsive Disorder. Am J Hum Genet 78, 815–826 (2006).
Kulikova, M. A. et al. Effect of functional catechol-O-methyltransferase Val158Met polymorphism on physical aggression. B Exp Biol Med 145, 62–64 (2008).
Jones, G. et al. Aggressive behaviour in patients with schizophrenia is associated with catechol-O-methyltransferase genotype. Brit J Psychiat 179, 351–355 (2001).
Guo, G., Roettger, M. E. & Shih, J. C. Contribution of the DAT1 and DRD2 genes to serious and violent delinquency among adolescents and young adults. Hum Genet 121, 125–136 (2007).
Sesack, S. R., Aoki, C. & Pickel, V. M. Ultrastructural localization of D2 receptor-like immunoreactivity in midbrain dopamine neurons and their striatal targets. J Neurosci 14, 88–106 (1994).
Chen, J. et al. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein and enzyme activity in postmortem human brain. Am J Hum Genet 75, 807–821 (2004).
Forbes, E. E. et al. Genetic variation in components of dopamine neurotransmission impacts ventral striatal reactivity associated with impulsivity. Mol Psychiatr 14, 60–70 (2009).
Alexander, N. et al. Genetic variants within the dopaminergic system interact to modulate endocrine stress reactivity and recovery. Behav Brain Res 216, 53–58 (2011).
Vaughn, M. G., DeLisi, M., Beaver, K. M. & Wright, J. P. DAT1 and 5HTT are associated with pathological criminal behavior in a nationally representative sample of youth. Crim Justice Behav 36, 1113–1124 (2009).
Beaver, K. M. et al. A gene × gene interaction between DRD2 and DRD4 is associated with conduct disorder and antisocial behavior in males. Behav Brain Funct 3, 1186 (2007).
Burt, S. A. & Mikolajewski, A. J. Preliminary evidence that specific candidate genes are associated with adolescent-onset antisocial behavior. Aggressive Behav 34, 437–445 (2008).
Barnes, J. C. & Jacobs, B. A. Genetic risk for violent behavior and environmental exposure to disadvantage and violent crime: The case for gene-environment interaction. J Interpers Violence 28, 92–120 (2013).
Miller, H. V. & Barnes, J. C. Intergenerational contact with the criminal justice system a consideration of three dopamine polymorphisms. Crim Justice Behav 40, 671–689 (2013).
Shen, H., Liu, Y., Liu, P., Recker, R. R. & Deng, H.-W. Nonreplication in genetic studies of complex diseases—Lessons learned from studies of osteoporosis and tentative remedies. J Bone Miner Res 20, 365–376 (2005).
Vassos, E., Collier, D. A. & Fazel, S. Systematic meta-analyses and field synopsis of genetic association studies of violence and aggression. Mol Psychiatry http://dx.doi.org/10.1038/mp.2013.31 (2013).
Beaver, K. M. & Chaviano, N. The Association Between Genetic Risk and Contact With the Criminal Justice System in a Sample of Hispanics. J Contemp Crim Justice 27, 81–94 (2011).
Freedman, M. L. et al. Assessing the impact of population stratification on genetic association studies. Nat Genet 36, 388–393 (2004).
Butovskaya, P. R. et al. Molecular-genetic polymorphisms of dopamine, serotonin and androgenic systems as molecular markers of success in judo wrestling sportsmen. J Bioanal Biomed S3, http://dx.doi.org/10.4172/1948-593X.S3-005 (2013).
Hurd, P. L., Vaillancourt, K. L. & Dinsdale, N. L. Aggression, digit ratio and variation in androgen receptor and monoamine oxidase A genes in men. Behav Genet 41, 543–556 (2011).
Butovskaya, M. L. et al. Aggression, digit ratio and variation in the Androgen Receptor, Serotonin Transporter and Dopamine D4 Receptor genes in African foragers: the Hadza. Behav Genet 42, 647–662 (2012).
Dmitrieva, J., Chen, Ch., Greenberger, E., Ogunseitan, O. & Ding, Y.-C. h. Gender-specific expression of the DRD4 gene on adolescent delinquency, anger and thrill seeking. SCAN 6, 82–89 (2011).
Butovskaya, M. L., Karelin, D. V. & Burkova, V. N. Datoga of Tanzania today: Ecology and cultural attitudes. Asia and Africa Today 11, 51–55 (2012).
Marlowe, F. The Hadza: Hunter-Gatherers of Tanzania. Berkeley: University of California Press. (2010).
Blystad, A. Fertile Mortal Links: Reconsidering Barabaig Violence Broch-Due, V. (ed.) Violence and Belonging: The Quest for Identity in Post-Colonial Africa 112–130 (London, New York: Routledge, 2005).
Gray, S. et al. Cattle raiding, cultural survival and adaptability of East African pastoralists. Curr Anthropol 44, S3–S30 (2003).
Lane, C. Pastures Lost: Barabaig Economy, Resource Tenure and the Alienation of Their Land in Tanzania (Nairobi: Initiatives Publishers, 1996).
Plomin, R., Haworth, C. M. A. & Davis, O. S. P. Common disorders are quantitative traits. Nat Rev Genet 10, 872–878 (2009).
Nikolova, Y. S., Ferrell, R. E., Manuck, S. B. & Hariri, A. R. Multilocus genetic profile for dopamine signaling predicts ventral striatum reactivity. Neuropsychopharmacol 36, 1940–1947 (2011).
Best, M., Williams, M. & Coccaro, E. F. Evidence for a dysfunctional prefrontal circuit in patients with an impulsive aggressive disorder. Proc Natl Acad Sci U S A 99, 8448–8453 (2002).
Buss, A. H. & Perry, M. The aggression questionnaire. J Pers Soc Psychol 63, 452–459 (1992).
Harris, J. A. A further evaluation of the aggression questionnaire: Issues of validity and reliability. Behav Res Ther 35, 1047–1053 (1997).
Book, A. S., Starzyk, K. B. & Quinsey, V. L. The relationship between testosterone and aggression: a meta-analysis. Aggress Violent Beh 6, 579–599 (2001).
Vasilyev, V. A. et al. Polymorphism of the dopamine D4 receptor (DRD4) and serotonin transporter (5-HTTL) gene promoter regions in African tribes of Hadza and Datoga. Russ J Genet 47, 226–229 (2011).
Acknowledgements
This study was partly supported by RFHR (project no. 12-01-00032), the Federal Innovation Program (no. 16.740.11.0172), the Ministry of Education and Science of the Russian Federation (project no. 8776), the RAS Program “Molecular and Cell Biology”, RFBR (projects no. 12-04-31869 and 13-04-00858) and the RF President's Program (no. 5233.2012.4). This study would have been utterly impossible without the tolerance and constant help of our Datoga friends.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: M.L.B., A.P.R., A.M.K., V.A.V. Performed the experiments: M.L.B., V.A.V., E.M.S., D.V.S. Data collection: M.L.B., D.V.K., V.N.B. Supporting data collection: A.M. Analyzed the data: M.L.B., A.M.K., O.E.L. Wrote the paper: M.L.B.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Participants: Cultural and ecological backgrounds
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Butovskaya, M., Vasilyev, V., Lazebny, O. et al. Aggression and polymorphisms in AR, DAT1, DRD2 and COMT genes in Datoga pastoralists of Tanzania. Sci Rep 3, 3148 (2013). https://doi.org/10.1038/srep03148
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03148
This article is cited by
-
Androgen receptor gene and sociosexuality. Does fighting ability moderate the effect of genetics in reproductive strategies?
Current Psychology (2023)
-
Facial and body sexual dimorphism are not interconnected in the Maasai
Journal of Physiological Anthropology (2022)
-
Oxytocin receptor gene polymorphism (rs53576) and digit ratio associates with aggression: comparison in seven ethnic groups
Journal of Physiological Anthropology (2020)
-
Behavioral Genetics, Population Genetics, and Genetic Essentialism
Science & Education (2020)
-
Serotonergic gene polymorphisms (5-HTTLPR, 5HTR1A, 5HTR2A), and population differences in aggression: traditional (Hadza and Datoga) and industrial (Russians) populations compared
Journal of Physiological Anthropology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
