
Abstract
Atmospheric CO2 concentrations appear to have been considerably higher than modern levels during much of the Phanerozoic and it has hence been proposed that surface temperatures were also higher. Some studies have, however, suggested that Earth's temperature (estimated from the isotopic composition of fossil shells) may have been independent of variations in atmospheric CO2 (e.g. in the Jurassic and Cretaceous). If large changes in atmospheric CO2 did not produce the expected climate responses in the past, predictions of future climate and the case for reducing current fossil-fuel emissions are potentially undermined. Here we evaluate the dataset upon which the Jurassic and Cretaceous assertions are based and present new temperature data, derived from the isotopic composition of fossil brachiopods. Our results are consistent with a warm climate mode for the Jurassic and Cretaceous and hence support the view that changes in atmospheric CO2 concentrations are linked with changes in global temperatures.
Similar content being viewed by others

Introduction
During the Phanerozoic, climate has alternated between states characterised by widespread glaciation (icehouses) and largely ice-free hothouse (or warmhouse) conditions1,2,3,4,5,6. Quasi-periodic icehouse-hothouse cycles of varying length have been proposed7,8,9 and many2,8,10 have suggested that that these major changes in Phanerozoic climate are linked to variations in atmospheric CO2. A number of studies have, however, cast doubt on the CO2-climate link4 and suggest instead that the galactic cosmic ray flux is the main driver of Phanerozoic climate change5,11. Although not fully accepted by many3,12,13, if correct, then this view has significant implications for our predictions of future climate change and the case for reducing anthropogenic carbon emissions6.
The basis for this assertion is a putative temperature record5,14,15 based upon the oxygen isotopic composition of well-preserved marine fossils (principally brachiopods, belemnites and planktonic foraminifera). This temperature record5 shows two mismatches between temperature and atmospheric CO2: during the Late Ordovician glaciation (~445 million years ago) and the Early Jurassic to Early Cretaceous interval (~180–100 million years ago), both of which coincided with model2,16 and proxy indications10,18 of high levels of atmospheric CO2. The Late Ordovician paradox has been particularly well studied17,18,19,20. A lower solar luminosity at this time suggests that the CO2 threshold for initiating a glaciation may have been higher18. Moreover, if the cooling pulse that led to ice volumes to expand to a similar size as those of the Pleistocene was relatively short-lived19, atmospheric CO2 proxies may not have recorded this brief reduction in CO2 levels. Furthermore, it has also been suggested that tropical ocean temperatures in excess of 30°C during the Late Ordovician coexisted with substantial south polar ice sheets19. Significant differences in the geosphere, e.g. the distribution of the continents and biosphere, e.g. the absence of significant terrestrial vegetation and calcareous plankton, during the Late Ordovician suggest that the links between CO2 and climate may well have operated differently to more recent times.
The Early Jurassic to Early Cretaceous anomaly presents a more critical test of the relationship between CO2 and temperature change, given that Earth’s palaeogeography and biota were more similar to those of the present-day. Although the Mesozoic is not a direct analogue for future greenhouse warming, such warm intervals in Earth history provide important insights into processes operating in the climate system21. Isotope derived temperature curves for this interval4 suggest significant cooling at this time, whereas CO2 proxy and model data typically indicate that atmospheric CO2 reached peaks not recorded since the Devonian2. Here we present extensive temperature data, derived from the isotopic composition of fossil brachiopods in order to evaluate the hypothesized Jurassic-Cretaceous cooling trends and re-assess Phanerozoic trends.
Results
Over 170 new oxygen isotope measurements from Mesozoic calcitic brachiopods are presented, combined with a further 300 brachiopod-derived measurements from published data22,23,24,25 (Figs. 1, 2) in order to test the robustness of cooling events suggested in previous studies4,5 and test whether galactic cosmic ray flux is potentially a driver of Phanerozoic climate change. The new dataset (see SI) covers the Jurassic to Cretaceous time interval. Almost all shells are derived from shallow epicontinental seas of the European region representing the tropical to subtropical Tethys. The data are constrained by ammonite biozone resolution (~0.4 to 1 Myr on average) allowing the generation of a reliable temporal series calibrated to numerical ages26. Our cathodoluminescence (CL) and petrographic analyses (SI) of the calcitic shells demonstrate excellent ultrastructure preservation of the analysed material. The trace element concentrations of each of the brachiopod shells corresponds to the results of the petrographic and CL analyses, whereby relatively low concentrations of Mn and Fe also argues for well-preserved fossils3,14,15.

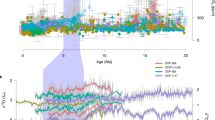
Evolution of Mesozoic-Cenozoic δ18O values.
Smoothed curves (red curve12,13 and brachiopod-foraminifera only isotope data, blue curve) are generated using Kernel regressions with a bandwidth (h) of 10 Myr. Abbreviation: Trias = Triassic. Grey dots = brachiopod data, red circles = belemnite data and green circles = planktonic foraminifera12,13 and black circles = brachiopod data compiled in this study.

Evolution of Phanerozoic δ18O values.
(A): Smoothed brachiopod-foraminifera only curve generated using Kernel regressions with a bandwidth (h) of 10 Myr. Abbreviations: Camb = Cambrian; Ord = Ordovician; Sil = Silurian; Carbon = Carboniferous; Perm = Permian; Trias = Triassic. Green shaded boxes represent glaciations and icehouses12. (B): SiZer map of the isotope data derived from brachiopods and foraminifera only. Blue shading indicates a statistically significant increase in isotopic values (i.e. cooling) and the confidence interval is completely above a slope of 0. Red shading indicates that the confidence interval is completely below a slope of 0 and there is a significant decrease (i.e. warming). Purple shading indicates where the confidence interval for the derivative contains 0. White fill indicates where the data are too sparse. The lower panel C shows the atmospheric CO2 concentration record as determined from multiple proxy reconstructions16 (green curve) and CO2 estimates (yellow curve) from GEOCARB III2.
Discussion
Our new brachiopod-derived δ18O isotope data from the Jurassic and Early Cretaceous are consistently more negative than coeval published data14,15 (Fig. 1). The most likely explanation is that the published data derive from different fossil taxa; the Jurassic and Early Cretaceous portion of the published database is dominated by data derived from belemnites (Cephalopoda, Coleoidea). Studies24,25,27 have also shown that belemnites typically display relatively more positive δ18O values than brachiopods; a difference possibly related to a combination of factors. Firstly, belemnites are interpreted as migratory nektobenthic organisms that were hence able to tolerate deeper and cooler waters, just as extant coleoids do28. Secondly, they may not have precipitated their calcite in isotopic equilibrium with seawater25. Furthermore, belemnites within the database are derived from a range of low, mid and high palaeolatitudes15 whereas the brachiopod data are derived predominantly from low (tropical and subtropical) palaeolatitudes14,15.
In contrast, the Late Cretaceous-Cenozoic proportion of the published database14,15 consists of isotope measurements derived largely from planktonic foraminifera. Where our new data and published brachiopod data24 are coeval with these planktonic data they show a good correspondence. The shells of planktonic foraminifera and brachiopods of similar age record comparable δ18O signatures, whereas coeval belemnites do not. Has, therefore, the inclusion of belemnite data in previous datasets14 resulted in erroneous temperature estimates for the Early Jurassic-Early Cretaceous interval?
As we have excluded a diagenetic influence (and assuming shell calcite is precipitated in isotopic equilibrium with seawater22,23,24,25), the δ18O composition of the brachiopods and planktonic foraminifera must reflect both the isotopic composition of the seawater in which the organisms lived and the water temperature at the time of shell growth. The isotopic composition of the seawater reflects changes in the size of ice-sheets, (ice-sheet growth preferentially removes 16O from seawater), the pH of seawater and possibly a long term change in seawater caused by tectonic processes29. Recent research using the ‘carbonate clumped isotope’ method3,19 is, however, inconsistent with such a secular variation in the δ18O of sea water. For this reason we have not adjusted the isotope data to accommodate for this (Figs. 1, 2) and the curve is interpreted in terms of temperature change. Given the potential problems with including belemnite data, we have combined our new data with the brachiopod- and foraminifera-derived data14,15 only, without including any belemnite data, to produce a novel, high resolution Mesozoic temperature curve.
When the critical Early Jurassic to Early Cretaceous interval is examined, it is clear that the new curve of mean temperature diverges significantly from the previous one (Fig. 1). The maximum difference in δ18O is 2.5 ‰, equivalent to ca. 8°C mean temperature change. We suggest the inclusion of belemnite data4 skews interpretations by creating the impression that temperatures were some 8°C cooler than the new data suggest.
The excellent agreement between our new Mesozoic brachiopod data and the published data derived from coeval planktonic foraminifera enables us to confidently link the published Palaeozoic dataset4 (derived almost entirely from brachiopods) with Cenozoic data (derived almost entirely from planktonic foraminifera), providing a new Phanerozoic δ18O (temperature) curve that is free of biases related to the use of belemnite data in the Mesozoic. To test the relationship between temperature and CO2 at various temporal scales through the Phanerozoic, we use the “SiZer” statistical approach (Significance of Zero Crossings of the Derivative30). This method is based on the construction of curves fitting time series using different levels of temporal smoothing (h). The first derivatives of each curve (i.e., the slopes) are computed with their 95% confidence intervals, allowing the signs of derivative estimates to be statistically tested. The results of these multiple tests reported in the form of SiZer maps (Fig. 1) enable significant features to be identified at different time scales. SiZer has been successfully applied to studies of the Quaternary31 and of the Jurassic27, but this is the first Phanerozoic-scale analysis.
The long-term, first-order Phanerozoic trend is one of overall cooling (i.e. increase in δ18O) from the Cambrian/Ordovician boundary to the present day, which matches the first-order decline in CO2 over the same interval (Fig. 2). Our SiZer analysis of the Palaeozoic record indicates two major cooling episodes (through the Ordovician and from the mid-Devonian to end-Carboniferous), both of which culminate with well-known glacial maxima9. Significant repeated shorter-term trends are apparent within the Ordovician and Silurian, a time characterised by warming and cooling/glacial and interglacial cycles19,20. We are therefore confident that our approach will provide a robust means to evaluate Mesozoic-Cenozoic trends.
Our SiZer analysis reveals warming during the Permian corresponding closely to rising CO2 levels, reaching a peak around the Permian/Triassic boundary32. Following the Permian/Triassic temperature maximum, our new Mesozoic brachiopod δ18O record suggests an overall cooling trend through most of the Triassic, mirroring the CO2 estimates from the GEOCARB III model2 and the GEOCLIM32 and COPSE models33. There is less agreement between our data and other proxy CO2 estimates18 in particular those derived from paleosol carbonates10. Through the Jurassic, the long term temperature trend is one of relative stasis. Evidence for long lived and extensive ice sheets during this time is lacking, although short transient events may characterise this time8,34. The SiZer analysis only records one cooling episode in the Jurassic. Indeed, for this interval a wealth of data is available suggesting that high latitude warmth is the norm1,8,18. The apparent long term climatic stasis continues through to the Late Cretaceous (ca. 95 Ma), when a return to increasing δ18O values indicate the onset of another cooling trend, consistent with temperature predictions from the COPSE model33. The transition from equable temperatures to long-term cooling is observed as CO2 levels fell below 1000 ppm. The long–term trend of climatic stability for at least 100 million years (largely encompassing the Jurassic and Early Cretaceous) recorded in our brachiopod δ18O data is also reconcilable with the CO2 predictions for this interval, although there is some disagreement between model and proxy estimates of atmospheric CO2 levels10,16,18 (Fig. 2C).
Our new brachiopod-derived δ18O isotope curve overturns the earlier and contentious suggestions that during the Early Jurassic to Early Cretaceous Earth experienced a cool climate mode. Inclusion of belemnite δ18O data is shown to bias the temperature interpretation towards cooler estimates. Whilst the data are consistent with two icehouse-hothouse intervals lasting tens of millions of years within the Palaeozoic, the Mesozoic-Cenozoic shows a different pattern. The lack of a prolonged cooling episode leading to glacial conditions may be a consequence of elevated atmospheric CO2 coupled with changing continental positions and higher solar luminosity. The Permo-Triassic hence appears as a major turning point in the climatic, as well as biotic35, history of the Earth. Carbon cycle modelling estimates of CO2 changes over the Mesozoic, in particular, correlate well with these changes in temperature. Together our observations support the assertion that atmospheric CO2 is consistent with patterns of past global climate change on a Phanerozoic time scale and serves to invalidate claims against a long-term CO2-climate link.
Methods
For stable isotope ratios we use the conventional terminology and δ-notation: δ18O = (Rsample/Rstandard-1) * 1000. Rsample and Rstandard are the 18O/16O ratio in the sample and standard, respectively. Positive or negative δ18O values mean that the sample has more or less 18O in the standard in parts per thousand (permil, ‰). The PDB standard, is a Cretaceous marine fossil (Belemnitella americana) and has now been exhausted and replaced by the V-PDB standard. Stable isotopes were determined on a VG Instruments Optima Isotope Ratio Mass Spectrometer (at the University of Plymouth) using 200–300 μg carbonate. Isotopic results were calibrated against NBS-19. Reproducibility for both δ18O and δ13C was better than 0.1‰, based upon multiple sample analysis. Trace element geochemistry in conjunction with optical petrography and cathodoluminescence (CL) was used to determine the state of preservation of each of the brachiopods analysed. Cold cathode CL analysis was undertaken using a CITL CL MK 3A, mounted on a Nikon petrological microscope. Sub-samples for chemical analysis weighing 1–2 mg were dissolved in nitric acid and analysed using a Varian 725-ES ICP-AES. Based upon analysis of duplicate samples, reproducibility was better than +/− 3% of the measured concentration of each element.
References
Huber, B. T. Tropical Paradise at the Cretaceous Poles? Science 282, 2199–2200 (1998).
Berner, R. A. & Kothavala, Z. GEOCARB III: A revised model of atmospheric CO2 over Phanerozoic time. American Journal of Science 301, 182–204 (2001).
Came, R. E. et al. Coupling of surface temperatures and atmospheric CO2 concentrations during the Palaeozoic era. Nature 449, 198–201 (2007).
Veizer, J., Godderis, Y. & François, L. M. Evidence for decoupling of atmospheric CO2 and global climate during the Phanerozoic eon. Nature 408, 698–701 (2000).
Shaviv, N. J. & Veizer, J. Celestial driver of Phanerozoic climate? GSA Today 13, 4–10 (2003).
Kump, L. R. What Drives Climate? Nature 408, 651–652 (2000).
Fischer, A. G. Climatic oscillations in the biosphere, in Biotic Crises in Ecological and Evolutionary Time. Nitecki M. ed., Academic, New York, pp. 103–131 (1981).
Frakes, L. A. Climates throughout geologic time. Elsevier, 310 p. (1979).
Crowell, J. C. Pre-Mesozoic ice ages; their bearing on understanding the climate system. Geol. Soc. Am. Mem. 192, 106 p. (1999).
Breecker, D. O., Sharp, Z. D. & McFadden, L. D. Atmospheric CO2 concentrations during ancient greenhouse climates were similar to those predicted for A. D. 2100. PNAS 107, (2010).
Gies, D. R. & Helsel, J. W. Ice age epochs and the sun's path through the Galaxy. Astrophysical Journal 626, 844–848 (2005).
Royer, D. L., Berner, R. A., Montañez, I. P., Tabor, N. J. & Beerling, D. J. CO2 as a primary driver of Phanerozoic climate. GSA Today 14, 4–10 (2004).
Fletcher, B. J., Brentnall, S. J., Anderson, C. W., Berner, R. A. & Beerling, D. J. Atmospheric carbon dioxide linked with Mesozoic and early Cenozoic climate change. Nature Geoscience 1, 43–48 (2008).
Veizer, J. et al. 87Sr/86Sr, δ13C and δ18O evolution of Phanerozoic seawater. Chemical Geology 161, 59–88 (1999).
Prokoph, A., Shields, G. A. & Veizer, J. Compilation and time-series analysis of a marine carbonate δ18O, δ13C, 87Sr/86Sr and δ34S database through Earth history. Earth-Science Reviews 87, 113–133 (2008).
Berner, R. A. Addendum to “Inclusion of the weathering of volcanic rocks in the GEOCARBSULF model”. American Journal of Science 308, 100–103 (2008).
Poussart, P. F., Weaver, A. J. & Barnes, C. R. Late Ordovician glaciation under high atmospheric CO2: a coupled model analysis. Paleoceanography 14, 542–558 (1999).
Royer, D. L. CO2-forced climate thresholds during the Phanerozoic. Geochimica et Cosmochimica Acta 70, 5665–5675 (2006).
Finnegan, S. et al. The magnitude and duration of Late Ordovician–Early Silurian Glaciation. Science 331, 903–906 (2011).
Turner, B. R., Armstrong, H. A. & Holt, P. Visions of ice sheets in the early Ordovician greenhouse world: Evidence from the Peninsula Formation, Cape Peninsula, South Africa. Sedimentary Geology 236, 226–238 (2011).
Crowley, T. J. Are there any satisfactory geologic analogs for a future greenhouse warming? Journal of Climate 3, 1282–1292 (1990).
Carpentier, C., Martin-Garin, B., Lathuiliere, B. & Ferry, S. Correlation of reefal Oxfordian episodes and climatic implications in the eastern Paris Basin (France). Terra Nova 18, 191–20 (2006).
Martin-Garin, B., Lathuilière, E. B., Geister, J. & Ramseyer, K. Oxygen isotopes and climatic control of Oxfordian coral reefs (Jurassic, Tethys). Palaios 25, 721–729 (2010).
Suan, G. et al. Secular environmental precursors to Early Toarcian (Jurassic) extreme climate changes. Earth Planet. Sci. Lett. 290, 448–458 (2010).
Voigt, S. Wilmsen, M. Mortimore, R. N. & Voigt, T. Cenomanian palaeotemperatures derived from the oxygen isotopic composition of brachiopods and belemnites: evaluation of Cretaceous palaeotemperature proxies. Int. Jour. Earth Sciences 92, 285–299 (2003).
Gradstein F. M., Ogg J. G., & Smith A. G. (Eds.), A Geologic Time Scale 2004. Cambridge University Press, Cambridge, 589 p. (2004).
Dera, G. et al. Climatic ups and downs in a disturbed Jurassic world. Geology 39, 215–218 (2011).
Price, G. D., Twitchett, R. J., Smale, C. & Marks, V. Isotopic analysis of the life history of the enigmatic squid Spirula spirula, with implications for studies of fossil cephalopods. Palaios 24, 273–279 (2009).
Jaffrés, J., Shields, G. A. & Wallmann, K. The oxygen isotope evolution of seawater: a critical review of a long-standing controversy and an improved geological water cycle model for the past 3.4 billion years. Earth-Science Reviews 83, 83–122 (2007).
Marron, J. S. & Chaudhuri, P. When is a feature really there? The SiZer approach. in Firooz A. ed., Automatic target recognition VII:. Photo optic and Industrial Engineering Proceedings 3371, 306–312 (1998).
Erästö, P. & Holmström, L. Selection of prior distributions and multiscale analysis in Bayesian temperature reconstructions based on fossil assemblages. Journal of Paleolimnology 36, 69–80 (2006).
Donnadieu, Y. et al. GEOCLIM simulation of climatic and biogeochemical consequences of Pangea breakup. Geochem. Geophys. Geosyst. 7, Q11019, 10.1029 (2006).
Bergman, N. M., Lenton, T. M. & Watson, A. J. COPSE: A new model of biogeochemical cycling over Phanerozoic time. American Journal of Science, 304, 397–437.
Price, G. D. The evidence and implications of polar ice during the Mesozoic. Earth-Science Reviews 48, 183–210 (1999).
Erwin, D. H. The Great Paleozoic Crisis: Life and Death in the Permian. Columbia University Press, 327 p. (1993).
Acknowledgements
This study was funded by an EU FP7 grant (acronym EMBiCC; grant number PIEF-GA-2009-237449) and a NERC Small Grant (NE/F011091/1) to RJT and GDP. We thank Scientific Reports editors and reviewers for constructive comments on an earlier version of the paper.
Author information
Authors and Affiliations
Contributions
G.D.P. and R.J.T. designed the study. J.R.W. and G.B. performed isotope and geochemical analyses. G.D.P. and R.J.T. analysed and interpreted the data. The manuscript incorporates comments on content and structure from all authors.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
SUPPLEMENTARY INFO
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Price, G., Twitchett, R., Wheeley, J. et al. Isotopic evidence for long term warmth in the Mesozoic. Sci Rep 3, 1438 (2013). https://doi.org/10.1038/srep01438
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep01438
This article is cited by
-
The early Cretaceous was cold but punctuated by warm snaps resulting from episodic volcanism
Communications Earth & Environment (2024)
-
Late Campanian-Maastrichtian in Pondicherry Area (Cauvery Basin) Southern India: Bioevents and Palaeoenvironmental Inferences from Planktonic Foraminifera
Journal of the Geological Society of India (2023)
-
The PhanSST global database of Phanerozoic sea surface temperature proxy data
Scientific Data (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
