
Abstract
The antidiabetic drug metformin efficiently circumvents the dilemma that in reducing the tumourigenicity of stem cells, their essence, specifically their pluripotency, must also be sacrificed. Metformin prevents the occurrence or drastically reduces the size and weight of teratoma-like masses after the transplantation of induced pluripotent stem (iPS) cells into immunodeficient mice. Yet, iPS cells implanted into metformin-treated mice retain full pluripotency, as they produce the same number of distinct tissue types derived from the three embryonic germ layers that is observed in untreated mice. Mechanistically, metformin appears to suppress the Oct4-driven compartment of malignant stem cells responsible for teratocarcinoma growth while safeguarding an intact, Oct4-independent competency to generate terminally differentiated tissues. Metformin's ability to efficiently and specifically control the tumourigenic fate of teratoma-initiating iPS cells without interfering with their pluripotency not only has implications for the clinical use of iPS cells but also in stem cell biology, cancer and ageing.
Similar content being viewed by others


Introduction
A fundamental principle of cell biology is that the greater the self-renewal and pluripotency that stem cells possess, the higher the probability that they will cause tumours1,2. In this regard, the most attractive pluripotent stem cells in cell biology research and regenerative medicine, induced pluripotent stem (iPS) cells, are not only characterised by their developmental potential but also by their inherent tumourigenic properties1,2,3,4. Sharing the basic properties of self-renewal and pluripotency with embryonic stem (ES) cells, iPS cells also share the tumourigenic traits of ES cells. iPS cells are indeed more tumourigenic than ES cells and harbour a risk for the development of benign teratomas or more aggressive teratocarcinomas4. Unlike ES cells, iPS cells also harbour a risk for the formation of somatic tumours due to genetic and epigenetic causes that can be acquired during reprogramming or from the somatic tissue of origin. In this scenario, in which pluripotency and tumourigenicity appear to be inevitably bound, there is an urgent need for strategies that mitigate the tumourigenic traits of iPS cells while preserving their pluripotency and multilineage differentiation potential.
We are accumulating strong evidence that while the AMPK (AMP-activated protein kinase) agonist metformin (N′,N′-dimethylbiguanide), the most widely used drug for type 2 diabetes, specifically regulates the expression of cancer stem cell (CSC)-specific genes to efficiently disrupt the CSC compartment in multiple cancers5,6,7,8, metformin-like drugs may also control the balance of self-renewal and differentiation of embryonic and adult stem cells9. We recently hypothesised that, by interfering with mechanisms that are important for tumourigenesis but are dispensable for adult stem cell development in mature tissues, the pharmacological activation of the metabolic tumour suppressor AMPK10,11 might decrease the teratocarcinogenic potential of iPS cells without compromising their ability to generate cells from all three germ layers. Therefore, we decided to explore how the systemic delivery of metformin could differentially impact the permissiveness of fully undifferentiated iPS cells with respect to tumourigenesis and pluripotency in animal models.
Results
We employed iPS cells derived from mouse embryonic fibroblasts (MEFs) to investigate the effect of the AMPK activator metformin on the tumourigenic potential of pluripotent stem cells. Individual iPS clones were selected by morphological criteria, i.e., flat colonies that were composed of small cells with a high nucleus to cytoplasm ratio and prominent nucleoli and characterised by immunostaining for the pluripotency markers alkaline phosphatase (AP), Ssea-1, Oct4 and Nanog ( Fig. 1 ). Then, we monitored the effects of systemic metformin on the dynamics of teratoma-like mass formation after the injection of iPS cells into immunodeficient mice. Nude mice were injected subcutaneously (s.c.) with 1 × 106 undifferentiated iPS cells; this “saturating” concentration of cells ensured the development of palpable tumour masses within a few weeks ( Fig. 1 ). One week later, the mice were randomised to receive daily intraperitoneal (i.p.) injections of vehicle (PBS) or metformin. The metformin dose used for the mice in our study was 200 mg/kg/day, which is equivalent to 960 mg/day for a 60 kg person12; therefore, this dose was less than half the recommended safe dose for humans (i.e., 2,550 mg/day for an average sized person of 60 kg) and significantly lower than that commonly employed in diabetic patients. We then performed side-by-side comparisons of the tumour growth dynamics and histological tumour composition of the iPS-derived teratomas in the absence or presence of systemic metformin.

Uncoupling tumourigenicity from pluripotency in stem cells using the antidiabetic biguanide metformin.
The cartoon summarizes the experimental design. Initially, we reprogrammed passage-one C57BL/6 mouse embryonic fibroblasts (MEFs) with three of the Yamanaka factors (Oct4, Sox2 and Klf4). iPS colonies grown over a MEFs feeder layer were stained either for alkaline phosphatase (AP) activity using the StemTAG™ Alkaline Phosphatase Staining and Activity Assay Kit (Cell Biolabs, Inc. Cat. No. CBA-302) or with antibodies against Ssea-1, Oct4 or Nanog, as indicated in the “Methods” section. Nuclear staining was performed with Hoechst 33258. Images show representative images of iPS colonies that were captured using different channels for Ssea-1 (green), Oct4 (green), Nanog (green) or Hoechst 33258 (blue). iPS cells were maintained in an undifferentiated stage with culturing on Matrigel-coated plates in the presence of LIF and bFGF for at least two passages for depletion of feeder cells before subcutaneous injection (1 × 106 cells per injection point and one subcutaneous injection point per animal) in nude mice.
In long-term teratoma formation assays, in which animals were sacrificed based on stringent predefined health parameters, a substantial reduction in the formation and growth of teratomas in mice treated with metformin was observed compared with untreated control mice. Whereas the rate of teratoma formation was 100% (10/10 mice) when iPS cells were transplanted into untreated control mice, significantly fewer (i.e., 5/10) metformin-treated mice developed any macroscopic teratomas throughout the entire course of the assay and these tumours were observed to be smaller than those in the untreated control mice. Compared to the mean teratoma size in the untreated control group (mean±SD = 9703±1497 mm3), the mean teratoma size in the metformin-treated group was drastically smaller (925±26 mm3) after 40 days of metformin treatment (Fig. 2A). Thus, the initial tumour volume increased less than 20-fold in the metformin-treated group but more than 200-fold in the untreated control group. Moreover, the growth of the formed teratomas was virtually unchanged after approximately 25 days of treatment with metformin; in contrast, teratoma volumes increased very rapidly in untreated mice. Consequently, i.p. administration of metformin significantly reduced teratoma weight in nude mice compared to PBS-treated control animals. The mean weight of the teratomas excised from mice treated with metformin (0.35±0.08 g) was approximately 90% less than that of the teratomas excised from untreated control mice (3.1±0.82 g) (Fig. 2B). Notably, when metformin administration was suspended on day 40 in teratoma-free mice and the mice were followed for four additional weeks, a palpable tumour with histologically confirmed teratoma-like tissues was only detected in one out of five mice on day 68 (data not shown).

Systemic metformin reduces the tumourigenic potential of induced pluripotent stem (iPS) cells.
iPS cells obtained as described in Fig. 1 were injected subcutaneously in nude mice. (A) Teratomas growth rate was calculated by measuring their volume along several days after iPS cell injection. Mean teratomas volume values (symbols) and standard deviations (bars) are shown. Note that teratomas formed in vehicle-treated mice presented a drastically higher growth rate than those derived from metformin-treated mice. (B) Left. 40 days after injection of iPS cells, mice were sacrificed and teratomas were extracted and weighed. Right. Note that teratomas from both the vehicle-treated group (A–C) and the metformin-treated group (a–c) similarly shown mixed tissues apparently derived from the three germ layers (i.e., ectoderm, mesoderm, endoderm). Indicated statistics in (A) and (B) were performed using a Student's t-test. (MET: Metformin).
The ability to form teratoma-like lesions that recapitulate the three germ layers generated during development, i.e., ectoderm, mesoderm and endoderm, is the gold-standard assay for determining whether potential iPS cell candidates are pluripotent. Because bona fide pluripotent stem cells that give rise to teratomas have the capacity to differentiate into any cell type and should give rise to all of the tissues found in the adult body, it follows that teratomas should be composed of various recognisable tissue elements, such as skin, bone and cartilage. Histological assessment also showed that in the untreated control group, mice developed tumours composed of ectoderm-derived neuroepithelial rosettes recapitulating early mammalian neurulation events, retinal pigment epithelium with glial elements, cystic cavities showing keratinising stratified squamous epithelium, endoderm-derived mucinous and ciliated cysts, mucinous intestinal-type glands and mesoderm-derived cartilage, adipose tissue containing lipoblasts, osteoid (bone-like) tissue and skeletal muscle tissue ( Fig. 3 ). Although their tumours were smaller, metformin-treated mice transplanted with iPS cells were capable of fully generating teratoma-like disorganised structures that harboured all of the differentiated tissues of the three embryonic germ layers observed in untreated control mice ( Fig. 3 ). Therefore, our results indicate that metformin treatment does not interfere with the pluripotency of iPS cells, i.e., iPS cells can still differentiate toward multiple lineages in the presence of systemic metformin in vivo ( Fig. 3 ).

Systemic metformin preserves the pluripotency of induced pluripotent stem (iPS) cells.
Histological analyses of teratomas derived from iPS cells upon subcutaneous transplantation reveal similar tissue composition in vehicle- (control) and metformin-treated mice. Vehicle (A–J)- and metformin-treated (a–j) iPS cells similarly developed into primitive tissues representing all germ layers including neural tissues (ectoderm; (A–C); (a–c)), glandular or respiratory epithelial layers (endoderm; (D–F), (d–f)), or cartilage (mesoderm; (G–J), (g–j)). There were numerous ectodermal neuroepithelial-like tissues including retinal epithelium-like tissues as well as glandular structures. Primitive neural tissues and neural rosettes were somewhat increased in teratomas from metformin-treated mice. In some cases glandular tissues displayed branching from the wall of the cyst into the lumen. Mesodermal-derived tissues likewise filled into the space between ectoderm- and endoderm-derived tissues including cartilage and fibrous connective tissues. Few and small bone spicules can be observed in some samples. (MET: Metformin).
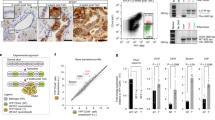
Our findings strongly support a direct role of the AMPK agonist metformin in limiting the teratocarcinoma formation and growth of iPS cells in vivo. We did not observe a significant increase in the apoptotic rate of metformin-treated teratomas (data not shown), suggesting that limited cell proliferation was the major cause for the metformin-induced reduction in teratoma number and size. Because a malignant undifferentiated population of Oct4-positive embryonic carcinoma (EC) cells has been shown to be ultimately responsible for the proliferation of teratomas derived from the injection of mouse iPS cells in immunocompromised mice3,13,14 (and therefore, they should be described as bona fide teratocarcinomas) we explored whether metformin exposure could reduce the EC component of poorly differentiated, primitive-appearing, blast-like teratocarcinoma stem cells in a Oct4-related manner. To directly explore functional effects of metformin on Oct4 expression, iPS cells maintained in an undifferentiated stage on matrigel-coated dishes in the presence of leukaemia inhibitory factor (LIF) were exogenously supplemented with 1 mmol/L metformin for 48 h. Immunofluorescence analysis using confocal microscopy revealed that metformin exposure results in drastically decreased levels of the Oct4 protein compared to vehicle-treated iPS cells ( Fig. 4A , left). The suppression of the nuclear accumulation of Oct4 in the presence of metformin was confirmed by immunoblotting analysis ( Fig. 4B , left). Immunohistochemical analyses revealed also a complete suppression of EC-like cells that were positive for the self-renewal transcription factor Oct4 ( Fig. 4A , right). The metformin's ability to downregulate Oct4 even though the culture media was supplemented with LIF, thus mimicking the gradual decrease of Oct4 protein levels throughout the period of differentiation when culturing iPS cells in medium without LIF, together with the suppression of the Oct4-related proliferative compartment of teratomas, strongly suggests that metformin-regulated Oct4 can underlie both the interference with the tumourigenic traits of iPS cells that lead to teratocarcinoma formation and the safeguarding of an intact iPS-driven pluripotent program. We finally explored how metformin treatment impacted the expression of the early pluripotency marker Ssea-1 as well as the activation status of other crucial members of the network for pluripotency (i.e., Sox2 and Nanog) ( Fig. 4B ). Treatment of LIF-supplemented iPS colonies with 1 mmol/L metformin was sufficient to notably reduce Ssea-1 protein expression (data not shown); however, significantly higher concentrations of metformin (i.e., 10 mmol/L) were necessary to induce major decreases in the expression of Sox2 and Nanog in individual iPS cells ( Fig. 4B ).

Metformin suppresses the in vitro and in vivo expression of Oct4.
(A) Left. iPS cells were maintained in an undifferentiated stage on matrigel-coated plates in the presence of LIF. After 48 h of treatment with vehicle (left panels) or 1 mmol/L metformin (right panels), Oct4 (green) levels were analyzed by immunofluorescent confocal microscopy. DNA was counterstained with Hoechst 33258 (blue). Images are representative of five independent experiments testing two individual iPS clones. Right. Two independent teratomas were stained for Oct4 expression by immunohistochemistry from vehicle- (top) and metformin-treated (bottom) mice, as indicated. Black arrowheads mark areas of Oct4 expression. (B) Left. Representative Western blot shows Oct4, Nanog and Sox2 protein expression in MEFs and iPS cells in the absence or presence of graded concentrations of metformin. Note that robust expression of endogenous Oct4 in iPS cells was drastically reduced in the presence of 1 mmol/L metformin, while MEFs do not express Oct4. Right. iPS cells were maintained in an undifferentiated stage on matrigel-coated plates in the presence of LIF. After 48 h of treatment with vehicle or 10 mmol/L metformin, the expression status of Nanog (green) and Sox2 (green) were analyzed by immunofluorescent confocal microscopy. DNA was counterstained with Hoechst 33258 (blue). Images are representative of five independent experiments testing two individual iPS clones. (MET: Metformin).
Discussion
Based on reports demonstrating that as few as twenty undifferentiated pluripotent cells are sufficient to generate a tumour, a considerable amount of effort has been focused on new technologies for the derivation of iPS cells with reduced tumourigenicity and improved safety4,14. However, the dilemma facing investigators is that in reducing the tumourigenicity of stem cells, the very essence of stem cells that makes them useful also has to be reduced, namely their pluripotency. In other words, reducing the tumourigenic nature of stem cells may inevitably reduce their self-renewal and pluripotency competences. Our current approach demonstrates, for the first time, that the AMPK agonist metformin can regulate the tumourigenicity of mouse iPS cells without altering their pluripotent properties. Given that Oct4 is critically involved in the tumourigenic capacity of teratoma-initiating iPS cells3,13,14 but is dispensable for maintaining the pluripotency of somatic stem cells15, this Pou domain-containing protein, which is downstream of the AMPK agonistic effects of metformin11,16, can be considered an excellent candidate for a metformin target that may participate in metformin's ability to control the cell fate of teratoma-initiating iPS cells. Considering that the under-expression and inactivation of AMPK are common phenomena that accompany carcinogenesis5,6, our results now support a model in which stemness and tumourigenicity are highly related processes but separable via evolutionary-conserved metabolic master sensors such as AMPK.
The previously unrecognised stemotoxic nature of metformin may reveal a viable, inexpensive, non-toxic pharmacological strategy to obviate the chances of stem cell-facilitated tumourigenesis and to develop safer iPS-based tissue engineering and cell replacement therapies in a clinical context. Our current findings largely recapitulate a landmark approach to limit the tumourigenicity of iPS cells and increase their safety through increased copy number of tumour suppressors14. Similarly to metformin, an increased dosage of tumour suppressors (i.e., p53 or Ink4a/ARF) in iPS cells prevented the occurrence or drastically reduced the size and weight of teratoma-like masses after the transplantation of iPS cells into immunodeficient mice. In agreement with a milestone study revealing how metformin treatment can promote neurogenesis from endogenous adult stem cells17,18, our findings confirm also that metformin treatment preserves and somewhat increases the occurrence of neuroepithelial-like tissues during iPS differentiation in vivo. The multifaceted metabolic effects of metformin19,20,21 may appear as apparently incompatible with its dual (tumour suppressive & pluripotency preserving)-activity in iPS cells; however, it should be noted that the most widely accepted mechanism of metformin action (i.e., the activation of the metabolic tumour suppressor AMPK) occurs secondarily following the alteration of numerous metabolic pathways. For instance, considering that the demand for metabolic flux via one-carbon metabolism is unusually high in undifferentiated pluripotent stem cells22 and that AMPK activation by intermediates in the purine biosynthetic pathway has been shown to enhance differentiation of ES cells9, the recently unrevealed capacity of metformin to impair the carbon flow through the folate-related one-carbon metabolic pathways23 might explain, at least in part, the metformin's ability to efficiently and specifically control the tumourigenic fate of teratoma-initiating iPS cells without interfering with their pluripotency. The ability of metformin to suppress the expression of Oct4 and survivin11,24 -two pivotal proteins driving the compartment of malignant stem cells responsible for teratocarcinoma growth-13,14 and to circumvent the suppression of AMPK that allows iPS cells to avoid anabolic inhibition similarly to cancer cells25,26, can explain also the metformin's ability to eliminate the compartment of malignant stem cells responsible for teratocarcinoma growth while safeguarding an intact competency to generate terminally differentiated tissues. Indeed, the exact mechanism of activation of AMPK might be relevant in terms of the balance of self-renewal and differentiation. While treatment with metformin significantly reduces pluripotent stem cells markers, Oct4 and Ssea-1, with more limited effects on the expression of Sox2 and Nanog, treatment with the AMPK activator 5-aminoimidazole-4-carboxyamide ribonucleoside (AICAR) also markedly inhibits proliferation of ES cells and enhances erythroid differentiation by significantly reducing Nanog and Ssea-1 but without affecting expression of Oct49.
Although it could be argued that we examined iPS cells in a fully undifferentiated state, which are unlikely to be used in cell transplantation therapy, a metformin-based “tumourigenicity-pluripotency uncoupling strategy” could be of particular interest for iPS cells generated for the treatment of diseases that require genetic correction and carry an intrinsic risk of genomic abnormalities27,28. While it remains to be evaluated whether the administration of metformin allows for the specific elimination of teratoma-initiating pluripotent cells that are intermixed with the desired, differentiated iPS cell derivatives, metformin treatment might still be of interest when undifferentiated progenitor cells must be used for cell therapy. Given that metformin has been shown to increase lifespan in mouse models, with and without cancer prevention29,30, future work is urgently required to confirm whether metformin's ability to efficiently and specifically control the tumourigenic fate of iPS cells without interfering with their pluripotency can be exploited in terms of metformin-like therapeutic molecules that can efficiently rejuvenate the tissue maintenance and repair driven by endogenous stem cells while diminishing tumourigenic predisposition in aging tissues.
Methods
Generation of iPS cells
Mouse primary iPS cells were created by transducing genetically unmodified mouse embryonic fibroblasts (MEFs) with murine retroviruses that individually encoded the transcription factors Oct4, Sox2 and Klf4 following a previously described protocol11. Characterisation of bona fide iPS cell colonies was carried out by analysing pluripotent marker expression by alkaline phosphatase (AP) staining and the expression of Ssea-1, Oct4 and Nanog by immunohistochemistry. For feeder-free culture of iPS cells, the plate was coated with 0.3 mg/mL Matrigel (growth-factor reduced, BD Biosciences) at 4°C overnight. Unbound Matrigel was aspirated and the cells were washed with DMEM/F12. iPS cells were seeded on Matrigel-coated plates in MEF-conditioned ES cell medium supplemented with leukaemia inhibitory factor (LIF) and additional 4 ng/mL bFGF. The medium was changed every day.
Immunofluorescence staining
High-content confocal imaging was performed in 96-well clear bottom imaging tissue culture plates (Becton Dickinson Biosciences; San Jose, CA) optimized for automated imaging applications. Triton® X-100 permeabilization and blocking, primary antibody staining, secondary antibody staining using Alexa Fluor® 488 goat anti-rabbit/mouse IgGs (Invitrogen, Probes, Eugene, Oregon) and counterstaining (using Hoechst 33258; Invitrogen) were performed by following BD Biosciences protocols. Images were captured in different channels for Alexa Fluor® 488 (pseudocolored green) and Hoechst 33258 (pseudocolored blue) on a BD PathwayTM 855 Bioimager System (Becton Dickinson Biosciences, San Jose, California) with 20× or 40× objectives (NA 075 Olympus). Merging images were obtained according to the Recommended Assay Procedure using the BD AttovisionTM software.
Teratoma assay
To form teratomas, approximately 1 × 106 iPS cells were harvested from Matrigel-coated culture dishes and injected subcutaneously (s.c.) into the dorsal flank of female athymic nude mice (four to five weeks old, 23 to 25 g; Harlan Laboratories, France). After one week, the animals were randomised into two groups, with ten mice in each group: control (vehicle) and metformin-treated. The metformin group received a single intraperitoneal (i.p.) injection of metformin (200 mg/kg) daily. Mice were weighed once per week after dosing, teratoma sizes were measured daily with electronic callipers and tumour volumes were calculated using the following formula: volume (mm3) = length × width2 × 0.5. General health of the mice in response to teratoma development (e.g., subcutaneous teratomas started to cause ulceration on the skin) was daily monitored by a specialized veterinarian. Metformin-treated animals did not show any health problem during the experiments. The experiments were approved by the Institutional Animal Care and Use Committee (IACUC) of the Institut d'Investigació Biomèdica de Bellvitge (IDIBELL; Animal Use Protocol #6302 authorized by the Animal Experimental Commision from the Catalan Goverment, Barcelona, Spain).
Western blot
Cell extracts were prepared using RIPA buffer, resolved on NuPAGE 4–12% gradient Bis-Tris gels, transferred to nitrocellulose and hybridized using an anti-Oct3/4 antibody (Abcam®, cat. No. ab19857), an anti-mouse/human Nanog antibody (Stemgent, Inc., cat. No. 09-0020), an anti-Sox2 antibody (Chemicon® International, Cat. No. AB5603) and an anti-γ-tubulin antibody (Sigma, cat. No.T6557).
Immunohistochemistry
The teratomas were carefully dissected and removed in its entirety, fixed in 10% phosphate buffered formalin (3.6% formaldehyde) for 24 hours and paraffin-embedded. For histopathological analysis of teratomas, four micrometre-thick consecutive sections were cut and stained with haematoxylin and eosin according to standard procedures. For immunohistochemical studies Oct3/4 antibody (1:100) was used following manufacturer's instructions. The solutions obtained from Dako Cytomation were used to perform immunostaining. In brief, tissue sections were deparaffinised, unmasked, blocked with avidin-biotin and incubated with primary antibody overnight. The next day, the reaction was detected using 3,3-diaminobenzidine tetrahydrochloride plus (DAB+) as a chromogen according to the manufacturer's instructions (Dako). The positive cells stained dark brown. The slides were examined under a light microscope and representative pictograms were taken from a minimum of five or six different slides from each group. Immunofluorescence studies with a different Oct3/4 antibody (sc-5279, Santa Cruz Biotechnology) were carried out in parallel teratomas tissues samples on a BD Pathway™ 855 Bioimager by following the Bioimaging Assay Procedure recommended by BD Biosciences.
Statistics
Two-group comparisons were performed by the Student t test for paired and unpaired values.
References
Brickman, J. M. & Burdon, T. G. Pluripotency and tumorigenicity. Nat Genet 32, 557–558 (2002).
Knoepfler, P. S. Deconstructing stem cell tumorigenicity: a roadmap to safe regenerative medicine. Stem Cells 27, 1050–1056 (2009).
Blum, B. & Benvenisty, N. The tumorigenicity of human embryonic stem cells. Adv Cancer Res 100, 133–158 (2008).
Ben-David, U. & Benvenisty, N. The tumorigenicity of human embryonic and induced pluripotent stem cells. Nat Rev Cancer 11, 268–277 (2011).
Hirsch, H. A., Iliopoulos, D., Tsichlis, P. N. & Struhl, K. Metformin selectively targets cancer stem cells and acts together with chemotherapy to block tumor growth and prolong remission. Cancer Res 69, 7507–7511 (2009).
Vazquez-Martin, A., Oliveras-Ferraros, C., Del Barco, S., Martin-Castillo, B. & Menendez, J. A. The anti-diabetic drug metformin suppresses self-renewal and proliferation of trastuzumab-resistant tumor-initiating breast cancer stem cells. Breast Cancer Res Treat 126, 355–364 (2011).
Song, C. W. et al. Metformin kills and radiosensitizes cancer cells and preferentially kills cancer stem cells. Sci Rep 3, 362 (2012).
Bao, B. et al. Metformin inhibits cell proliferation, migration and invasion by attenuating CSC function mediated by deregulating miRNAs in pancreatic cancer cells. Cancer Prev Res (Phila) 5, 355–364 (2012).
Chae, H. D., Lee, M. R. & Broxmeyer, H. E. 5-Aminoimidazole-4-carboxyamide ribonucleoside induces G(1)/S arrest and Nanog downregulation via p53 and enhances erythroid differentiation. Stem Cells 30, 140–149 (2012).
Hardie, D. G. AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function. Genes & Dev 25, 1895–1908 (2011).
Vazquez-Martin, A. et al. Activation of AMP-activated protein kinase (AMPK) provides a metabolic barrier to reprogramming somatic cells into stem cells. Cell Cycle 11, 974–989 (2012).
Reagan-Shaw, S., Nihal, M. & Ahmad, N. Dose translation from animal to human studies revisited. FASEB J 22, 659–661 (2008).
Blum, B., Bar-Nur, O., Golan-Lev, T. & Benvenisty, N. The anti-apoptotic gene surviving contributes to teratoma formation by human embryonic stem cells. Nat Biotechnol 27, 281–287 (2009).
Menendez, S., Camus, S., Herreria, A., Paramonov, I., Morera, L. B. & Collado, M. et al. Increased dosage of tumor suppressors limits the tumorigenicity of iPS cells without affecting their pluripotency. Aging Cell 11, 41–50 (2012).
Lengner, C. J., Welstead, G. G. & Jaenisch, R. The pluripotency regulator Oct4: a role in somatic stem cells? Cell Cycle 7, 725–728 (2008).
Jung, J. W., Park, S. B., Lee, S. J., Seo, M. S., Trosko, J. E. & Kang, K. S. Metformin represses self-renewal of the human breast carcinoma stem cells via inhibition of estrogen receptor-mediated OCT4 expression. PLoS One 6, e28068 (2011).
Wang, J. et al. Metformin Activates an Atypical PKC-CBP Pathway to Promote Neurogenesis and Enhance Spatial Memory Formation. Cell Stem Cell 11, 23–35 (2012).
Menendez, J. A. & Vazquez-Martin, A. Rejuvenating regeneration: Metformin activates endogenous stem cells. Cell Cycle 11, 3521–2 (2012).
Martin-Castillo, B., Vazquez-Martin, A., Oliveras-Ferraros, C. & Menendez, J. A. Metformin and cancer: doses, mechanisms and the dandelion and hormetic phenomena. Cell Cycle 9, 1057–1064 (2010).
Vazquez-Martin, A., Oliveras-Ferraros, C., Cufí, S., Martin-Castillo, B. & Menendez, J. A. Metformin and energy metabolism in breast cancer: from insulin physiology to Tumour-initiating stem cells. Curr Mol Med 10, 674–691 (2010).
Del Barco, S. et al. Metformin: multi-faceted protection against cancer. Oncotarget 2, 896–917 (2011).
Wang, J., Alexander, P., Wu, L., Hammer, R., Cleaver, O. & McKnight, S. L. Dependence of mouse embryonic stem cells on threonine catabolism. Science 325, 435–439 (2009).
Corominas-Faja, B. et al. Metabolomic fingerprint reveals that metformin impairs one-carbon Metabolism in a manner similar to the antifolate class of Chemotherapy drugs. Aging (Albany NY) 4, 480–498 (2012).
Vazquez-Martin, A., Oliveras-Ferraros, C., del Barco, S., Martin-Castillo, B. & Menendez, J. A. The antidiabetic drug metformin: a pharmaceutical AMPK activator to overcome breast cancer resistance to HER2 inhibitors while decreasing risk of cardiomyopathy. Ann Oncol 20, 592–595 (2009).
Prigione, A. et al. Human induced pluripotent stem cells harbor homoplasmic and heteroplasmic mitochondrial DNA mutations while maintaining human embryonic stem cell-like metabolic reprogramming. Stem Cells 29, 1338–1334 (2011).
Vander Heiden, M. G., Cantley, L. C. & Thompson, C. B. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science 324, 1029–1033 (2009).
Raya, A. et al. Disease-corrected haematopoietic progenitors from Fanconi anaemia induced pluripotent stem cells. Nature 460, 53–59 (2009).
Liu, G. H. et al. Targeted gene correction of laminopathy-associated LMNA mutations in patient-specific iPSCs. Cell Stem Cell 8, 688–694 (2011).
Anisimov, V. N. Metformin for aging and cancer prevention. Aging (Albany NY) 2, 760–774 (2010).
Menendez, J. A., Cufí, S., Oliveras-Ferraros, C., Vellon, L., Joven, J. & Vazquez-Martin, A. Gerosuppressant metformin: less is more. Aging (Albany NY) 3, 348–362 (2011).
Acknowledgements
We are indebted to Prof. Carlos Lopez-Otin and his team (Departamento de Bioquímica y Biología Molecular, Facultad de Medicina, Instituto Universitario de Oncología [IUOPA], Universidad de Oviedo, Oviedo, Spain) for providing most of the reagents and descriptions of the procedures used in this work. This work was financially supported through funding from the Instituto de Salud Carlos III (Ministerio de Sanidad y Consumo, Fondo de Investigación Sanitaria [FIS], Spain, Grants CP05-00090 and PI06-0778 and RD06-0020-0028), the Fundación Científica de la Asociación Española Contra el Cáncer (AECC, Spain) and the Ministerio de Ciencia e Innovación (SAF2009-11579, Plan Nacional de I+D+I, MICINN, Spain). Alejandro Vazquez-Martin received the Sara Borrell post-doctoral contract (CD08/00283, Ministerio de Sanidad y Consumo, Fondo de Investigación Sanitaria [FIS], Spain). Sílvia Cufí received a Research Fellowship (Formación de Personal Investigador [FPI]) from the Ministerio de Ciencia e Innovación (MICINN, Spain).
Author information
Authors and Affiliations
Contributions
J.A.M., A.V.M. and S.C. designed the study. A.V.M., S.C., E.L.B. and C.O.F. performed the experiments. J.A.M., E.L.B. and B.M.C. analysed the data. J.A.M. wrote the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Vazquez-Martin, A., Cufi, S., Lopez-Bonet, E. et al. Metformin limits the tumourigenicity of iPS cells without affecting their pluripotency. Sci Rep 2, 964 (2012). https://doi.org/10.1038/srep00964
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00964
This article is cited by
-
Immunologic targeting of CD30 eliminates tumourigenic human pluripotent stem cells, allowing safer clinical application of hiPSC-based cell therapy
Scientific Reports (2018)
-
Health benefits of late-onset metformin treatment every other week in mice
npj Aging and Mechanisms of Disease (2017)
-
Technical approaches to induce selective cell death of pluripotent stem cells
Cellular and Molecular Life Sciences (2017)
-
Novel Role of Mitochondrial Manganese Superoxide Dismutase in STAT3 Dependent Pluripotency of Mouse Embryonic Stem Cells
Scientific Reports (2015)
-
Immunogenicity and functional evaluation of iPSC-derived organs for transplantation
Cell Discovery (2015)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
