
Abstract
Observational studies in humans have found associations between overstimulation in infancy via excessive television viewing and subsequent deficits in cognition and attention. We developed and tested a mouse model of overstimulation whereby p10 mice were subjected to audio (70 db) and visual stimulation (flashing lights) for six hours per day for a total of 42 days. 10 days later cognition and behavior were tested using the following tests: Light Dark Latency, Elevated Plus Maze, Novel Object Recognition and Barnes Maze. In all tests, overstimulated mice performed significantly worse compared to controls suggesting increased activity and risk taking, diminished short term memory and decreased cognitive function. These findings suggest that excessive non-normative stimulation during critical periods of brain development can have demonstrable untoward effects on subsequent neurocognitive function.
Similar content being viewed by others

Introduction
First discovered by Hebb in 1947 environmental enrichment is widely known to significantly enhance memory functions1,2,3,4,5. Environmental enhancement results in numerous neurobiological changes including increased dendritic branching in cortical neurons6,7, increased size and complexity of the superior colliculus6 and increased neurogenesis in the hippocampus1,8,9,10,11,12. The key features of enriched environment includes multiple sensory experiences (sensory integration) and the active engagement with the novel environment13,14,15.
Building on the importance of these early environmental experiences in mice, the Institute of Medicine report highlighted how early human environments should likewise be optimized to enhance brain development16. What followed was an explosion of products, many of them video-based, that claim that they stimulated infant brains making them for example “smarter” or more “musical” with no empiric basis of support17. As a result, in the last 30 years, the average age at which children begin to regularly watch TV has moved from 4 years of age to 5 months of age and the typical preschool child spends as much as 30–40% of their waking time in front of a screen18,19,20,21,22.
Central, but implicit in the enriched environment studies is the notion that the stimulation provided was normative, that is consistent with what might be experienced by mice in a naturalistic setting. But in contrast to the pace with which real life unfolds, video programs, particularly those viewed by very young children are extremely rapidly and surreally sequenced23. Observational data in humans have supported the association between excessive viewing of such programs and subsequent attentional problems21,24. Unfortunately experimental data from infants are lacking and difficult to obtain for a variety of practical and ethical reasons although one brief experimental study found viewing a rapidly sequenced show lead to short term attentional problems in preschool children21,24,25.
Here we examined the behavioral consequences of “excessive non-normative stimulation” akin to what baby DVD's provide in a mouse model. Ours is a fundamental departure from the enriched environment approach as we tested the “opposite” hypothesis: excessive, non-normative stimulation during a similar period will diminish performance subsequently.
Results
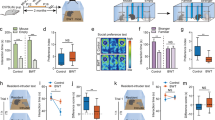
We mounted speakers to standard mouse cages and positioned colored light sources at all four walls. (Figure 1). To mimic television viewing we piped audio from the “cartoon channel” into the mouse cage at 70 decibels, which is well below the levels of 100–115 db that are typically used for acoustic stress models and consistent with National Institutes of Health acceptable noise levels26,27,28,29,30,31. A photorhythmic modulator was used to change colors and intensities in concordance with the audio thereby simulating television that cannot be avoided (e.g. flashing lights on all four sides of the cage).

Mouse overstimulation chamber and experimental procedure.
Beginning at postnatal day 10 (P10), mice were randomly divided into two groups: (a) a control group which was reared according to approved and established protocols at the Seattle Children's Research Institute Vivarium; and (b) an overstimulated group which was treated identically to the control group except that they were exposed for 6 hours every night to auditory and visual stimulation intended to generate “non-normative sensory stimulation.” Exposure lasted for 42 days, which is comparable to the length commonly used in enriched environment studies8. Mice in both groups remained with their mother throughout this period to avoid any additional handling. Following the exposure period, lights and speakers were removed, but the mice remained in their familiar, regular mouse cages. Beginning 10 days later, we performed the following behavioral tests: the Open Field test32, Light Dark Latency test33, the Elevated Plus Maze34, the Novel Object Recognition test35 and the Barnes Maze36. For all tests, we made use of the VideoTrack (ViewPoint LS) tracking software to track mouse movements. The tracking software records where the mouse was, how long it was in certain areas, as well as the distance the mouse traveled in each area. Experiments were videotaped and in all cases, technicians blinded to research group made assessments.
We first assessed anxiety and activity. The Elevated Plus Maze measures the anxiety induced by open spaces, as well as height. The apparatus was an elevated maze consisting of four arms making the shape of a plus sign (Figure 2). Two of the arms had walls around them, whereas the other two arms project out of the center without walls. All mice were placed in the center of the maze and were allowed to run freely around the maze for 5 minutes. We calculated the amount of time the mouse spent in each arm, the distance traveled and the number of entries the mouse made into each arm. Overstimulated mice spent significantly more time in the open arm (Fig. 2E), had more entries into the open arm (Fig. 2F) and traveled greater distances in the open arm (Fig. 2G) compared to controls.

Results of Elevated Plus Maze (EPM) and Light Dark Latency (LDL) Tests.
2A & 2B demonstrate an illustrative example of a control and an overstimulated mouse's travel pathway on the EPM. Overstimulated mice spent more time in the open arm (mean±SEM; Controls: 9.93±2.11s, n = 48 and Overstimulated: 31.03±2.78s, n = 61, p < 0.001). (2E); had more entries in the open arms (mean±SEM; Controls: 7.97±1.22, n = 48 and Overstimulated: 11.92±1.06, n = 61, p < 0.001). (2F); and traveled greater distances in the open arms (mean±SEM; Controls: 73.55±23.61cm, n = 48 and Overstimulated: 176.23±14.62cm, n = 61, p < 0.001). (2G). 2C & 2D demonstrate an illustrative example of a control and an overstimulated mouse's travel pathway on the LDL. Overstimulated mice spent more time in the light chamber (mean±SEM; Controls: 53.79±4.17s, n = 48 and Overstimulated: 82.39±6.41s, n = 61, p < 0.001) (2H); took less time to fully enter the light chamber (mean±SEM; Controls: 223.00±17.49s, n = 48 and Overstimulated: 98.00±15.33, n = 61, p < 0.001) (2I) and traveled greater distances in the light chamber (mean±SEM; Controls: 276.42±42.03cm, n = 48 and Overstimulated: 436.89±40.06cm, n = 61, p <0.05) (2J).
The Light Dark Latency test measures risk-taking and anxiety in a related but different way. It is based on the rodents' innate aversion to brightly illuminated areas as well as their competing exploratory instincts37,38. The apparatus consisted of two compartments. One compartment is made of translucent fiberglass the other is painted black. The two compartments are separated by a black dividing wall with a small doorway for the mouse to travel between them. A 100 W light was directed at the translucent (light) chamber from approximately a 15 cm distance. The light is blocked out of the black chamber with a lid making it dark. Each mouse was placed in the dark chamber and was allowed to run freely between the two chambers for 5 minutes. Latency to enter (defined by all four paws entered), time spent, entries and distance traveled in the light chamber were recorded. Overstimulated mice were significantly different from control mice for all domains (Figure 2).
The Open Field Test measures hyperactivity and anxiety in yet a third way. The open field box consisted of a square black box made out of plexiglass with an outlined center area. Each animal was placed in the box for ten minutes. Overall activity in the box (measured with videotrack) was measured as well as the amount of time and distance traveled in the center area of the maze. Mice naturally prefer to be near a protective wall rather than exposed to danger out in the open, but a competing foraging instinct will motivate them to explore. (Figure 3A). Overstimulated mice spent significantly more time in the center (Fig. 3A,B), entered significantly more times into the center and traveled more distance within the center than control mice (Figure 3B). Moreover, the overall distance traveled by the overstimulated mice was significant higher than the distance traveled by the control mice, suggesting that overstimulated mice are hyperactive. (Figure 3C).

Results of Open Field Test: Tests.
3A & 3B demonstrate an illustrative example of a control and an overstimulated mouse's travel pathway on the Open Field Test. Overstimulated mice spent more time in the center of the open field (mean±SEM; Controls: 24.97±2.04, n = 64 and Overstimulated: 44.25±3.76, n = 72, p < 0.001) (3C), had more entries into the center of the open field (mean±SEM; Controls: 20.69±1.72, n = 72 and Overstimulated: 34.94±2.72, n = 72, p < 0.001) (3D) and they were more active as measured in distance traveled in the center of the open field field (mean±SEM; Controls: 505.79±47.45, n = 72 and Overstimulated: 883.03±71.82, n = 72, p <0.001), (3E) They also traveled greater distance overall (mean±SEM; Controls: 5836.66±194.26, n = 72 and Overstimulated: 6612.43±200.5, n = 72, p< 0.001) (3F).
We next assessed memory and learning. The Novel Object Recognition Test assesses short term memory by testing a mouse's ability to remember if it had previously encountered an object or not39. It is based on the idea that a mouse will spend more time investigating and exploring an object which it has never seen (novel object), compared to an object it has encountered before (familiar object). The same box, which was used in the Open Field Test, was used for the Novel Object Recognition test. Mice were habituated to the box a day before the test, during the Open Field Test. On the test day, mice were placed in the box for the acquisition period with two identical objects and were allowed to explore and familiarize themselves with the objects for 5 minutes. The mice were given a one hour inter-trial interval (ITI) and were then placed back in the test box. Everything was the same as during the acquisition period except that one of the two identical objects was replaced with a new, novel object. During the testing period, mice were allowed to explore both of the objects for 5 minutes. This trial was recorded with a video camera and scored by a trained and blinded experimenter to determine the time the mouse spent investigating each object. During the testing period, overstimulated mice spent less time on the novel object compared to controls. (Figure 4E).

Results of Barnes Maze Test (BM) and Novel Object Recognition Test (NORT).
4A & B demonstrate an illustrative example of a control and an overstimulated mouse's travel pathway on the BM. 4C summarizes the results of the BM over 4 days of successive testing. Overstimulated mice trended towards finding the escape hole faster than controls during the first day of the experiments (mean±SEM; Controls: 30.60±4.64, n = 12 and Overstimulated: 20.08±2.35, n = 72, p = 0.09) which is consistent with our findings that exposed animals travel greater distances and thus are more likely to encounter the escape hole. However, on day 4, control mice took significantly less time to find the target hole compared to overstimulated mice (mean±SEM; Controls: 3.44±0.39, n = 12 and Overstimulated: 6.33±0.67, n = 10, p < 0.001) Overstimulated mice continued to make more errors before finding the hole suggesting impaired memory and learning. Figure 4D summarizes the mean times mice spent on both objects during the acquisition (mean±SEM; Controls: 44.35±4.61, n = 39 and Overstimulated: 57.49±4.01, n = 42) and the test trial (mean±SEM; Controls: 44.87±5.64, n = 39 and Overstimulated: 53.81±3.77, n = 42). Figure 4E shows the discrimination ratio for the controls and the overstimulated mice. This shows how well the mice were able to discriminate between the novel and the familiar object. Overstimulated mice spent less time with a novel object compared to controls. Accordingly the discrimination ratio calculated as ((Time Spent on the Novel Object – Time Spent On the Familiar Object)/Total Time) is significantly reduced compared to controls (mean±SEM; Controls: 0.32±0.07, n = 39 and Overstimulated: 0.16±0.05, n = 42, p < 0.05).
Finally, we tested the mice with the Barnes Maze which is a circular platform with an escape hole that leads to a small chamber attached underneath the platform. There are 19 other imitation holes circled around the maze in order to distract from the real hole. The imitation holes look like the escape hole, but do not lead to an escape chamber. The Maze is lit up with a 100 w light bulb. On day one, the mouse was placed in an opaque box in the center of the maze. After ten seconds had elapsed, the opaque box was removed and the mouse was guided by hand to the escape hole. The first trial began after 15 minutes. On trial one, the mouse was once again placed in the start box for 10 seconds. Once released, the mouse had three minutes to find the escape hole. As soon as the mouse found the hole the light was turned off. The mouse spent 1 minute in the escape chamber and was then returned to its original housing cage. This procedure was recorded with the video camera and tracking software and latency to find the escape hole, as well as distance traveled was recorded. Mice were given four learning trials a day with 15-minute ITI's. On each day, the mean time to find the target hole was calculated. Overstimulated mice performed poorly compared to control mice on day 4 of testing. (Figure 4C).
Discussion
We have demonstrated in a controlled way that providing non-normative, excessive audiovisual stimulation during the early life of mice results in significant deficits in cognition and behavior. Specifically, overstimulated mice demonstrate increased risk taking/decreased anxiety, poorer short-term memory and impaired learning. These results are in marked contrast to what has been shown in the setting of “enriched” environments.
There are several limitations to this study that warrant consideration. First, it is possible that the overstimulation affected maternal rearing habits as they too were exposed to it. Future experiments should assess this directly. Prior studies of maternal stress and behavioral outcomes have yielded mixed results with some showing that maternal compensatory behaviors when the stress is removed eliminates untoward effects and others showing increased anxiety in offspring39,40. Neither of these is consistent with our observed results40,41. Furthermore, for a separate experiment, we collected weights on 24 adult mice (14 overstimulated and 10 control). There was no significant difference in their weights: Control 46.79 g vs Overstimulated 43.22 (p = .31). Prior studies have found maternal stress to lead to reduced adult weight in offspring42. Second, we cannot be certain that the observed effects were the result of overstimulation and not stress since it is conceivable that the lights and sounds were stress-inducing. However, we deliberately and set the volume to 70 decibels, which is well below the levels of 100–115 db that are typically used for acoustic stress models and at a level that has been deemed acceptable by federal guidelines in the United States26,27,28,29,30,31. Furthermore, our observation of decreased anxiety and increased travel in our overstimulated mice runs contrary to what has been demonstrated in newborn stress models where increased anxiety is typically seen27,28,29,30,31,43,44. Finally, stress may be in the causal pathway and mediate the effects of overstimulation. That is, overstimulation is stressful and that leads to untoward effects. If this is the case, controlling for it would not make sense although it would be worthwhile from a scientific standpoint to assess it. In future studies, we intend to explicitly explore this pathway by among other things, measuring serial serum cortisol levels on both groups of mice.
To what extent these results can be applied to human populations is unknown especially given that this is the first study to our knowledge to associate overstimulation with diminished cognitive performance and behavioral consequences. It should be noted however that the benefits of enriched environments previously demonstrated on mice have been replicated in humans suggesting that early experiences help tune the mind and improve executive function45,46. It appears then that a coherent model whereby early experiences establish neuronal circuits or habits of the mind which can either enhance or impede subsequent performance may be emerging.
Methods
Animals and Housing
Both males and female CD1 mice where used for all experiments. Mice were housed in groups (4–5 per cage) and provided with standard mouse chow and water at libitum. Mice were housed in a room that had a 12 h light/dark cycle (light on at 7:00 am) with controlled temperature (21 +/− 1 Celsius). The protocol was approved by the Seattle Children's Research Institutional Animal Care and Use Committee.
Overstimulation
The mice were housed with mother in regular housing conditions until they were 10 days old. At postnatal day (p) 10, the cage was moved into the overstimulation rack. The overstimulation rack consisted of two loud speakers, - suspended two inches above the top of the cage. Exposure sounds came from children's Television shows such as Pokemon, Powerpuff girls, Bakugan etc., which were layered on top of each other with one pitch shifted and one non pitch shifted track in order to better accommodate the higher frequency hearing rage of the mice. The volume was kept below 70 db the level used in acoustic stress models. To better simulate a television paradigm, LED lights (red, green, yellow and blue) were synchronized to go off in rhythm to match the sound output of the speakers. Mice were exposed during the night for 6 six hours a day (from 11 PM to 5 AM), starting at p. 10 until p. 52. After this they had a ten-day resting period before behavioral tests began.
Behavioral Tests
Light Dark Latency Test
This test detects anxiety-like behaviors in mice. The apparatus consisted of two compartments one which was made of translucent fiberglass (15 cm x 30 cm x 30 cm) and one, which was painted black (15cm x 30cm x 30cm). A Black-dividing wall with a small (5cm x 5cm) doorway for the mouse to travel through freely divided the two compartments. A 100 W light was directed at the translucent (light) chamber from approximately a 15 cm distance. The light is blocked out of the black chamber with a lid making it dark. Each mouse was placed in the dark chamber and was allowed to run freely between the two chambers for 5 minutes. The trial was video taped and a tracking device (Videotrack from ViewPoint Life Sci) recorded all of the mouse's movements. Latency to enter (defined by all four paws entered), time spent, entries and distance traveled in the light chamber were recorded.
The Elevated Plus Maze
The Elevated plus maze measures another type of anxiety, the anxiety induced by open spaces, as well as height related anxiety. The apparatus was an elevated (50 cm) maze consisting of four arms (30x5cm each) making the shape of a plus sign (+). Two of the arms had 20 cm walls around them, whereas the other two arms project out of the center without walls. All mice were placed in the center of the maze and were allowed to run freely around the maze for 5 minutes. Mice were recorded and tracked by videotrack (ViewPoint LS) in order to determine the amount of time the mouse spent in each arm, the distance traveled and the number of entries the mouse made into each arm.
The Open Field Test
The Open Field Test measures hyperactivity through locomotion and anxious behavior. The open field box consisted of a square black box (60cm x 60cm x 25cm), made out of plexiglass with an outlined center area. The center area (30cm x 30cm) was made of Vinyl Electrical Tape (Tartan 1710). Each animal was placed in the box for ten minutes. Overall activity in the box (measured with videotrack) was measured as well as the amount of time and distance traveled in the center area of the maze. This paradigm is based on the idea that mice will naturally prefer to be near a protective wall rather than exposed to danger out in the open.
The Novel Object Recognition Test
The Novel Object Recognition assesses a mouse's ability to remember if it haspreviously encountered an object or not. It is based on the idea that a mouse will spend more time investigating and exploring an object which it has never seen (novel object), than an object it has encountered before (familiar object). The same box, which was used in the Open Field Test, was used for the Novel Object Recognition test. Mice were habituated to the box a day before the test, during the Open Field Test. On the test day, mice were placed in the box for the acquisition period with two identical objects (A) and were allowed to explore and familiarize themselves with the objects for 5 minutes. The mice were given a one hour inter trial interval (ITI) and where then placed back in the test box. Everything was the same as during the acquisition period except that one of the two identical objects (A) was replaced with a new, novel object (B). During the testing period, mice were allowed to explore both of the objects for 5 minutes. This trial was recorded with a video camera and scored by a trained and blinded experimenter to determine the time the mouse spent investigating each object. Interaction was considered when the mouse's nose touched the object or was pointed towards the object within a 1cm radius. The discrimination ratio was calculated using the following formula: [(Time Spent on the Novel Object – Time Spent on the Familiar Object)/Total Time)].
The Barnes Maze
The Banes Maze is a (90 cm elevated) circular platform (100 cm diameter) with an escape hole (5 cm diameter), which leads to a small chamber attached underneath the platform. There are 19 other imitation holes circled around the maze in order to distract from the real hole. The imitation holes look like the escape hole, but do not lead to an escape chamber. The Maze is lit up with a 100 w light bulb. On day one, the mouse was placed in an opaque box in the center of the maze. After ten seconds had elapsed, the opaque box was removed and the mouse was guided by hand to the escape hole. The first trial began after 15 minutes. On trial one, the mouse was once again placed in the start box for 10 seconds. Once released, the mouse had three minutes to find the escape hole. As soon as the mouse found the hole the light was turned off. The mouse spent 1 minute in the escape chamber and was then returned to its original housing cage. This procedure was recorded with the video camera and tracking software and latency to find the escape hole, as well as distance traveled was recorded. Mice were given four learning trials a day with 15-minute ITI's. After four days of learning, a probe trial was performed where the mouse once again was placed on the maze. This time, the escape hole was covered, now virtually looking like all the other holes. The probe trial was recorded and time spent around the escape hole was recorded. We also looked at the amount of other holes that were searched. After 90 seconds the mouse was placed in its original housing cage.
Data Coding
Entries and time in each area was assessed by research assistants blinded to group assignment.
Statistical Analyses
All comparisons of means used two tailed student t tests. Means are reported with Standard Errors as shown in the figures.
References
Kempermann, G., Kuhn, H. G. & Gage, F. H. Experience-induced neurogenesis in the senescent dentate gyrus. J Neurosci 18, 3206–3212 (1998).
Kempermann, G. & Gage, F. H. Experience-dependent regulation of adult hippocampal neurogenesis: effects of long-term stimulation and stimulus withdrawal. Hippocampus 9, 321–332 (1999).
Kempermann, G., Kuhn, H. G. & Gage, F. H. More hippocampal neurons in adult mice living in an enriched environment. Nature 386, 493–495 (1997).
Rampon, C. et al. Effects of environmental enrichment on gene expression in the brain. Proc Natl Acad Sci U S A 97 (2000).
Tang, Y. P., Wang, H., Feng, R., Kyin, M. & Tsien, J. Z. Differential effects of enrichment on learning and memory function in NR2B transgenic mice. Neuropharmacology 41, 779–790 (2001).
Fuchs, J. L., Montemayor, M. & Greenough, W. T. Effect of environmental complexity on size of the superior colliculus. Behavioral & Neural Biology 54, 198–203 (1990).
Greenough, W. T., Larson, J. R. & Withers, G. S. Effects of unilateral and bilateral training in a reaching task on dendritic branching of neurons in the rat motor-sensory forelimb cortex. Behavioral & Neural Biology 44, 301–314 (1985).
Choi, S. H. et al. Non-cell-autonomous effects of presenilin 1 variants on enrichment-mediated hippocampal progenitor cell proliferation and differentiation. Neuron 59, 568–580 (2008).
Schloesser, R. J., Lehmann, M., Martinowich, K., Manji, H. K. & Herkenham, M. Environmental enrichment requires adult neurogenesis to facilitate the recovery from psychosocial stress. Mol Psychiatry 15, 1152–1163, mp201034 [pii] 10.1038/mp.2010.34 (2010).
Kuzumaki, N. et al. Hippocampal epigenetic modification at the brain-derived neurotrophic factor gene induced by an enriched environment. Hippocampus 21, 127–132 (2011).
Goshen, I. et al. Environmental enrichment restores memory functioning in mice with impaired IL-1 signaling via reinstatement of long-term potentiation and spine size enlargement. J Neurosci 29, 3395–3403 (2009).
Nithianantharajah, J. & Hannan, A. J. Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat Rev Neurosci 7, 697–709 (2006).
Burrows, E. L., McOmish, C. E. & Hannan, A. J. Gene-environment interactions and construct validity in preclinical models of psychiatric disorders. Prog Neuropsychopharmacol Biol Psychiatry 35, 1376–1382 (2011).
Liu, N., He, S. & Yu, X. Early Natural Stimulation through Environmental Enrichment Accelerates Neuronal Development in the Mouse Dentate Gyrus. PLoS One 7, e30803 (2012).
Reynolds, S., Lane, S. J. & Richards, L. Using animal models of enriched environments to inform research on sensory integration intervention for the rehabilitation of neurodevelopmental disorders. J Neurodev Disord 2, 120–132 (2010).
Institute of Medicine. From Neurons to Neighborhoods: The Science of Early Childhood Development. (National Academy Press, 2000).
Garrison, M. & Christakis, D. A teacher in the living room?: Educational media for babies, toddlers and preschoolers. (Kaiser Family Foundation, Menlo Park CA, 2005).
Christakis, D. A., Ebel, B. E., Rivara, F. P. & Zimmerman, F. J. Television, video and computer game usage in children under 11 years of age. J Pediatr 145, 652–656 (2004).
Christakis, D. A. & Zimmerman, F. J. The Elephant in the Living Room : Make Television work for your Kids. (Rodale, 2006).
Vandewater, E. A. et al. Digital childhood: electronic media and technology use among infants, toddlers and preschoolers. Pediatrics 119, e1006–1015 (2007).
Zimmerman, F. J. & Christakis, D. A. Associations between content types of early media exposure and subsequent attentional problems. Pediatrics 120, 986–992 (2007).
Tandon, P. S., Zhou, C., Lozano, P. & Christakis, D. A. Preschoolers' total daily screen time at home and by type of child care. J Pediatr 158, 297–300 (2011).
Goodrich, S. A., Pempek, T. A. & Calvert, S. L. Formal Production Features of Infant and Toddler DVDs. 163, 1151–1156 (2009).
Christakis, D. A., Zimmerman, F. J., DiGiuseppe, D. L. & McCarty, C. A. Early television exposure and subsequent attentional problems in children. Pediatrics 113, 708–713 (2004).
Lillard, A. S. & Peterson, J. The immediate impact of different types of television on young children's executive function. Pediatrics 128, 644–649 (2011).
Committee for the update of the Guide for the Care and use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals. Eighth Edition edn, (National Academies Press, 2009).
Berridge, C. W. & Dunn, A. J. CRF and restraint-stress decrease exploratory behavior in hypophysectomized mice. Pharmacol Biochem Behav 34, 517–519 (1989).
Berridge, C. W. & Dunn, A. J. Restraint-stress-induced changes in exploratory behavior appear to be mediated by norepinephrine-stimulated release of CRF. J Neurosci 9, 3513–3521 (1989).
Gilhotra, N. & Dhingra, D. GABAergic and nitriergic modulation by curcumin for its antianxiety-like activity in mice. Brain Res 1352, 167–175 (2010).
Heinrichs, S. C. et al. Anti-stress action of a corticotropin-releasing factor antagonist on behavioral reactivity to stressors of varying type and intensity. Neuropsychopharmacology 11, 179–186 (1994).
Dunn, A. J. & Berridge, C. W. Physiological and behavioral responses to corticotropin-releasing factor administration: is CRF a mediator of anxiety or stress responses? Brain Res Brain Res Rev 15, 71–100 (1990).
Prut, L. & Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. European Journal of Pharmacology 463, 3–33 (2003).
Bourin, M. & Hascoët, M. The mouse light/dark box test. European Journal of Pharmacology 463, 55–65 (2003).
Pellow, S., Chopin, P., File, S. E. & Briley, M. Validation of open : closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. Journal of Neuroscience Methods 14, 149–167 (1985).
Dere, E., Huston, J. P. & De Souza Silva, M. A. The pharmacology, neuroanatomy and neurogenetics of one-trial object recognition in rodents. Neuroscience & Biobehavioral Reviews 31, 673–704 (2007).
Paul, C.-M., Magda, G. & Abel, S. Spatial memory: Theoretical basis and comparative review on experimental methods in rodents. Behavioural Brain Research 203, 151–164 (2009).
Costall, B., Jones, B. J., Kelly, M. E., Naylor, R. J. & Tomkins, D. M. Exploration of mice in a black and white test box: Validation as a model of anxiety. Pharmacology Biochemistry and Behavior 32, 777–785 (1989).
Crawley, A. M., Anderson, D. R., Wilder, A., Williams, M. & Santomero, A. Effects of repeated exposures to a single episode of the television program Blue's Clues on the viewing behaviors and comprehension of preschool children. Journal of Educational Psychology 91, 630–638 (1999).
Ennaceur, A. & Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav Brain Res 31, 47–59 (1988).
Millstein, R. A. & Holmes, A. Effects of repeated maternal separation on anxiety- and depression-related phenotypes in different mouse strains. Neuroscience & Biobehavioral Reviews 31, 3–17 (2007).
Schechter, M., Pinhasov, A., Weller, A. & Fride, E. Blocking the postpartum mouse dam's CB1 receptors impairs maternal behavior as well as offspring development and their adult social–emotional behavior. Behavioural Brain Research 226, 481–492 (2012).
Gao, P. et al. Maternal stress affects postnatal growth and the pituitary expression of prolactin in mouse offspring. J Neurosci Res 89, 329–340, 10.1002/jnr.22550 (2011).
Chesworth, R. et al. The response of neuregulin 1 mutant mice to acute restraint stress. Neurosci Lett 515, 82–86 (2012).
Naqvi, F., Haider, S., Batool, Z., Perveen, T. & Haleem, D. J. Sub-chronic exposure to noise affects locomotor activity and produces anxiogenic and depressive like behavior in rats. Pharmacol Rep 64, 64–69 (2012).
Diamond, A., Barnett, W. S., Thomas, J. & Munro, S. Preschool program improves cognitive control. Science 318, 1387–1388(2007).
Diamond, A. & Lee, K. Interventions shown to aid executive function development in children 4 to 12 years old. Science 333, 959–964 (2011).
Author information
Authors and Affiliations
Contributions
DAC conceived of the idea and drafted the manuscript. JSBR designed and performed the behavioral experiments. All three authors analyzed the data and edited the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Christakis, D., Ramirez, J. & Ramirez, J. Overstimulation of newborn mice leads to behavioral differences and deficits in cognitive performance. Sci Rep 2, 546 (2012). https://doi.org/10.1038/srep00546
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00546
This article is cited by
-
A signalling pathway for transcriptional regulation of sleep amount in mice
Nature (2022)
-
Auditory processing remains sensitive to environmental experience during adolescence in a rodent model
Nature Communications (2022)
-
Early childhood screen time as a predictor of emotional and behavioral problems in children at 4 years: a birth cohort study in China
Environmental Health and Preventive Medicine (2021)
-
Effects of Excessive Screen Time on Neurodevelopment, Learning, Memory, Mental Health, and Neurodegeneration: a Scoping Review
International Journal of Mental Health and Addiction (2021)
-
Does physical activity moderate the association between screen time and psychosocial development in early childhood? Analysis of a longitudinal infant cohort study in Ireland
European Journal of Pediatrics (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
