
Abstract
The pleiotropic benefits of statins in cardiovascular diseases that are independent of their lipid-lowering effects have been well documented, but the underlying mechanisms remain elusive. Here we show that simvastatin significantly improves human induced pluripotent stem cell-derived endothelial cell functions in both baseline and diabetic conditions by reducing chromatin accessibility at transcriptional enhanced associate domain elements and ultimately at endothelial-to-mesenchymal transition (EndMT)-regulating genes in a yes-associated protein (YAP)-dependent manner. Inhibition of geranylgeranyltransferase (GGTase) I, a mevalonate pathway intermediate, repressed YAP nuclear translocation and YAP activity via RhoA signaling antagonism. We further identified a previously undescribed SOX9 enhancer downstream of statin–YAP signaling that promotes the EndMT process. Thus, inhibition of any component of the GGTase–RhoA–YAP–SRY box transcription factor 9 (SOX9) signaling axis was shown to rescue EndMT-associated endothelial dysfunction both in vitro and in vivo, especially under diabetic conditions. Overall, our study reveals an epigenetic modulatory role for simvastatin in repressing EndMT to confer protection against endothelial dysfunction.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
All the raw data files generated from this study including for RNA-seq, ATAC-seq and ChIP–seq were deposited at GEO (GSE157895). The public datasets used in this study include the following: human EC H3K36me3, https://www.encodeproject.org/experiments/ENCSR993KKI/; human EC H3K27me3, https://www.encodeproject.org/experiments/ENCSR436KET/; human EC H3K4me3, https://www.encodeproject.org/experiments/ENCSR306WYL/; human EC H3K4me3, https://www.encodeproject.org/experiments/ENCSR622UQY/; human EC H3K27ac, https://www.encodeproject.org/experiments/ENCSR616PRQ/; YAP1 ChIP–seq, GSE66081; and YAP–TAZ ChIP–seq, GSE94862. Other data were included in the article and source files.
Code availability
No custom-made code was used in this study.
References
Arnett, D. K. et al. 2019 ACC/AHA Guideline on the primary prevention of cardiovascular disease: executive summary: a report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 140, e563–e595 (2019).
Chou, R., Dana, T., Blazina, I., Daeges, M. & Jeanne, T. L. Statins for prevention of cardiovascular disease in adults: evidence report and systematic review for the US Preventive Services Task Force. JAMA 316, 2008–2024 (2016).
Sen-Banerjee, S. et al. Kruppel-like factor 2 as a novel mediator of statin effects in endothelial cells. Circulation 112, 720–726 (2005).
Laufs, U., La Fata, V., Plutzky, J. & Liao, J. K. Upregulation of endothelial nitric oxide synthase by HMG CoA reductase inhibitors. Circulation 97, 1129–1135 (1998).
Greenwood, J. & Mason, J. C. Statins and the vascular endothelial inflammatory response. Trends Immunol. 28, 88–98 (2007).
Jiang, Y. Z. et al. Hemodynamic disturbed flow induces differential DNA methylation of endothelial Kruppel-like factor 4 promoter in vitro and in vivo. Circ. Res. 115, 32–43 (2014).
Karlic, H. et al. Inhibition of the mevalonate pathway affects epigenetic regulation in cancer cells. Cancer Genet. 208, 241–252 (2015).
Tikoo, K. et al. Tissue specific up regulation of ACE2 in rabbit model of atherosclerosis by atorvastatin: role of epigenetic histone modifications. Biochem. Pharmacol. 93, 343–351 (2015).
Musunuru, K. et al. Induced pluripotent stem cells for cardiovascular disease modeling and precision medicine: a scientific statement from the American Heart Association. Circ. Genom. Precis. Med. 11, e000043 (2018).
Sayed, N. et al. Clinical trial in a dish using iPSCs shows lovastatin improves endothelial dysfunction and cellular cross-talk in LMNA cardiomyopathy. Sci. Transl. Med. 12, eaax9276 (2020).
Oda, N., Abe, M. & Sato, Y. ETS-1 converts endothelial cells to the angiogenic phenotype by inducing the expression of matrix metalloproteinases and integrin β3. J. Cell. Physiol. 178, 121–132 (1999).
Savant, S. et al. The orphan receptor Tie1 controls angiogenesis and vascular remodeling by differentially regulating Tie2 in tip and stalk cells. Cell Rep. 12, 1761–1773 (2015).
Sayed, N. et al. Transdifferentiation of human fibroblasts to endothelial cells: role of innate immunity. Circulation 131, 300–309 (2015).
Atkins, G. B. & Jain, M. K. Role of Kruppel-like transcription factors in endothelial biology. Circ. Res. 100, 1686–1695 (2007).
Feig, J. E. et al. Statins promote the regression of atherosclerosis via activation of the CCR7-dependent emigration pathway in macrophages. PLoS ONE 6, e28534 (2011).
Calo, E. & Wysocka, J. Modification of enhancer chromatin: what, how, and why? Mol. Cell 49, 825–837 (2013).
Lin, K. C., Park, H. W. & Guan, K. L. Regulation of the Hippo pathway transcription factor TEAD. Trends Biochem. Sci. 42, 862–872 (2017).
Jin, E. et al. Differential roles for ETS, CREB, and EGR binding sites in mediating VEGF receptor 1 expression in vivo. Blood 114, 5557–5566 (2009).
Zhang, H. et al. TEAD transcription factors mediate the function of TAZ in cell growth and epithelial–mesenchymal transition. J. Biol. Chem. 284, 13355–13362 (2009).
Zhao, B. et al. Inactivation of YAP oncoprotein by the Hippo pathway is involved in cell contact inhibition and tissue growth control. Genes Dev. 21, 2747–2761 (2007).
Mauviel, A., Nallet-Staub, F. & Varelas, X. Integrating developmental signals: a Hippo in the (path)way. Oncogene 31, 1743–1756 (2012).
Maruyama, J. et al. Novel YAP1 activator, identified by transcription-based functional screen, limits multiple myeloma growth. Mol. Cancer Res. 16, 197–211 (2018).
Juarez, D. & Fruman, D.A. Targeting the mevalonate pathway in cancer. Trends. Cancer. 7, 525–540 (2021).
Sorrentino, G. et al. Metabolic control of YAP and TAZ by the mevalonate pathway. Nat. Cell Biol. 16, 357–366 (2014).
Kaur, R., Kaur, M. & Singh, J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: molecular insights and therapeutic strategies. Cardiovasc. Diabetol. 17, 121 (2018).
Haffner, S. M., Lehto, S., Ronnemaa, T., Pyorala, K. & Laakso, M. Mortality from coronary heart disease in subjects with type 2 diabetes and in nondiabetic subjects with and without prior myocardial infarction. N. Engl. J. Med. 339, 229–234 (1998).
Cholesterol Treatment Trialists' (CTT) Collaborators et al. Efficacy of cholesterol-lowering therapy in 18,686 people with diabetes in 14 randomised trials of statins: a meta-analysis. Lancet 371, 117–125 (2008).
Ong, S. B. et al. Calpain inhibition restores autophagy and prevents mitochondrial fragmentation in a human iPSC model of diabetic endotheliopathy. Stem Cell Reports 12, 597–610 (2019).
Li, Y., Lui, K. O. & Zhou, B. Reassessing endothelial-to-mesenchymal transition in cardiovascular diseases. Nat. Rev. Cardiol. 15, 445–456 (2018).
Antonopoulos, A. S., Margaritis, M., Lee, R., Channon, K. & Antoniades, C. Statins as anti-inflammatory agents in atherogenesis: molecular mechanisms and lessons from the recent clinical trials. Curr. Pharm. Des. 18, 1519–1530 (2012).
Wang, K. C. et al. Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis. Proc. Natl Acad. Sci. USA 113, 11525–11530 (2016).
Monroe, T. O. et al. YAP partially reprograms chromatin accessibility to directly induce adult cardiogenesis in vivo. Dev. Cell 48, 765–779 (2019).
von Gise, A. & Pu, W. T. Endocardial and epicardial epithelial to mesenchymal transitions in heart development and disease. Circ. Res. 110, 1628–1645 (2012).
Zhao, J. et al. Sox9 and Rbpj differentially regulate endothelial to mesenchymal transition and wound scarring in murine endovascular progenitors. Nat. Commun. 12, 2564 (2021).
Lin, Q. Q., Zhao, J., Zheng, C. G. & Chun, J. Roles of Notch signaling pathway and endothelial–mesenchymal transition in vascular endothelial dysfunction and atherosclerosis. Eur. Rev. Med. Pharmacol. Sci. 22, 6485–6491 (2018).
Xin, M. et al. Regulation of insulin-like growth factor signaling by Yap governs cardiomyocyte proliferation and embryonic heart size. Sci. Signal. 4, ra70 (2011).
Wang, X. et al. YAP/TAZ orchestrate VEGF signaling during developmental angiogenesis. Dev. Cell 42, 462–478 (2017).
Chen, J. & Harris, R. C. Interaction of the EGF receptor and the Hippo pathway in the diabetic kidney. J. Am. Soc. Nephrol. 27, 1689–1700 (2016).
Ikeda, S. et al. Yes-associated protein (YAP) facilitates pressure overload-induced dysfunction in the diabetic heart. JACC Basic Transl. Sci. 4, 611–622 (2019).
Chen, L. & Liu, Z. Multifaceted function of YAP/TEAD on chromatin:prospects of ‘a non-canonical role of YAP/TEAD is required for activation of estrogen-regulated enhancers in breast cancer’. J. Mol. Cell Biol. 11, 1101–1103 (2019).
Wang, L. et al. Integrin–YAP/TAZ–JNK cascade mediates atheroprotective effect of unidirectional shear flow. Nature 540, 579–582 (2016).
Zhang, X. et al. The essential role of YAP O-GlcNAcylation in high-glucose-stimulated liver tumorigenesis. Nat. Commun. 8, 15280 (2017).
Kovacic, J. C. et al. Endothelial to mesenchymal transition in cardiovascular disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 73, 190–209 (2019).
Liu, C., Oikonomopoulos, A., Sayed, N. & Wu, J. C. Modeling human diseases with induced pluripotent stem cells: from 2D to 3D and beyond. Development 145, dev156166 (2018).
Saha, K. & Jaenisch, R. Technical challenges in using human induced pluripotent stem cells to model disease. Cell Stem Cell 5, 584–595 (2009).
Lau, E., Paik, D. T. & Wu, J. C. Systems-wide approaches in induced pluripotent stem cell models. Annu. Rev. Pathol. 14, 395–419 (2019).
Mandegar, M. A. et al. CRISPR interference efficiently induces specific and reversible gene silencing in human iPSCs. Cell Stem Cell 18, 541–553 (2016).
Wei, T. T. et al. Cannabinoid receptor 1 antagonist genistein attenuates marijuana-induced vascular inflammation. Cell 185, 1676–1693 (2022).
Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10, 1213–1218 (2013).
Acknowledgements
We thank S. Khanamiri (Stanford Cardiovascular Institute) for technical assistance on wire myography experiments and N.-Y. Shao for his constructive advice on transcriptomic analysis. We thank A. Mueller and X. Wu (Stanford Cardiovascular Institute) for their help in proofreading. This study was supported by research grants from the National Institutes of Health (NIH) (R01 HL113006, R01 HL150693, R01 HL163680, R01 HL145676, P01 HL141084, R01 HL141371 and R01 HL141851 to J.C.W.); NIH R01 HL158641, R01 HL161002 and the American Heart Association SFRN grant 869015 to N.S.; AHA Career Development Awards 19CDA34760019 to C.L.; NIH R01 HL155282 and 18CDA34110293 to M.-T.Z.; and the Tobacco-Related Disease Research Program (30FT0852 to M.S.). L.S.Q. is a Chan Zuckerberg Biohub–San Francisco Investigator. Schematic figures were created using https://BioRender.com.
Author information
Authors and Affiliations
Contributions
C.L. designed the study with J.C.W. and N.S. and performed most of the experiments and data analysis with M.S. W.L.W.T. processed the sequencing data and performed most of the bioinformatic analysis with Yu Liu’s help. I.Y.C. supplied iPSC-ECs for the experiments. X.Y. performed most of the wire myography experiments. H.Y. and A.Z. processed the EC function analysis. Yanxia Liu, M.-T.Z., M.A., M.Z. and E.R.G. discussed the results and strategy. C.L., M.S. and J.C.W. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
J.C.W. is a founder and A.Z. is a consultant of Greenstone Biosciences. The other authors declare no competing interests.
Peer review
Peer review information
Nature Cardiovascular Research thanks Peter Libby, Philip Marsden and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Statins improve endothelial function and alter epigenetic associated genes in iPSC-ECs.
a, NOS3 expression levels in iPSC-ECs after being treated with seven statins at different concentrations (0.1 μM, 1 μM, and 10 μM). Data are normalized to that of the vehicle control group; prava, pravastatin; atorva, atorvastatin; rosuva, rosuvastatin; meva, mevastatin; fluva, fluvastatin; lova, lovastatin; and simva, simvastatin (n = 6 biological samples). b, Simvastatin decreases the viability of iPSC-ECs at 10 µM (n = 3 biological samples). c, Simvastatin increases ROS production in iPSC-ECs at 10 µM (n = 3 biological samples). d, Representative images showing higher densities of capillary-like networks formed by iPSC-ECs from three healthy donors treated with simvastatin versus vehicle. Scale bars, 250 µm. e, Differentially expressed genes (DEGs) in simvastatin- versus vehicle-treated iPSC-ECs (FDR < 0.05). f, Simvastatin had no effect on iPSC-EC proliferation compared to vehicle (DMSO) (n = 4 biological samples). g, Enrichment (cellular components) analysis of upregulated DEGs shown in e. h, Enrichment (cellular components) analysis of downregulated DEGs shown in (e). All data are presented as mean ± SEM. Unpaired two-sided Student’s t-test (a,b,c).
Extended Data Fig. 2 Simvastatin alters chromatin accessibility in iPSC-ECs.
a, Annotation of ATAC peaks with differential chromatin accessibility. b and c, Motif enrichment analysis at the ATAC-seq sites with differential chromatin accessibility at enhancer regions. d, KEGG pathway analysis of the ATAC-seq peaks with KLF motifs. e and f, GO pathway analysis of ATAC-seq peaks with KLF motifs and TEAD motifs.
Extended Data Fig. 3 RNA-seq and ATAC-seq analyses of iPSC-ECs treated with simvastatin at different timepoints.
a, Heatmap of RNA-seq showing changes in transcriptomic patterns of iPSC-ECs after being treated with simvastatin at 0 h, 12 h, 24 h, and 72 h. b, Normalized expression levels of endothelial marker genes (KLF4, CDH5, and KDR) from RNA-seq of simvastatin-treated iPSC-ECs at different timepoints. c, Normalized expression levels of YAP downstream genes (SMAD3, CTGF, GLI2, MCL1, RUNX1, BIRC2, TGFB2, and BIRC5) from RNA-seq of simvastatin-treated iPSC-ECs at different timepoints. d, Normalized expression levels of mesenchymal genes (TGFBR1, TWIST1, SNAI2, and SOX9) from RNA-seq of simvastatin-treated iPSC-ECs at different timepoints. All data are presented as mean ± SEM. n = 2 RNA-seq biological samples.
Extended Data Fig. 4 Simvastatin inhibits YAP and TEAD activity in iPSC-ECs.
a, Comparative analysis showing that simvastatin exposure to iPSC-ECs for 12 h, 24 h, and 72 h induces highly correlated changes in DEGs and ATAC signal alterations (values indicate log2(fold change)). b, Immunofluorescence of YAP subcellular localization in iPSC-ECs treated with simvastatin versus vehicle control for 24 h. c, Quantification of nuclear/cytoplasmic YAP ratios in iPSC-ECs treated with simvastatin versus vehicle control for 24 h (n = 14 cells). d, iPSC-ECs treated with simvastatin (n = 101 cells) but not vehicle (n = 138 cells) for 24 h showing significantly decreased nuclear TEAD activity. e, Representative images of capillary-like tubular networks formed by iPSC-ECs treated with simva, simva + MA, simva + GGPP, simva + squalene, GGTi298, RhoAi, and vehicle control. All data are presented as mean ± SEM. Unpaired two-sided Student’s t-test (c,d).
Extended Data Fig. 5 Simvastatin reverses hyperglycemia (HG)-induced endothelial dysfunction by repressing YAP-mediated EndMT process.
a, ROS levels in iPSC-ECs treated with HG, HG + simvastatin, HG + GGTi298, HG + RhoAi, and vehicle control (n = 3 biological samples). b, Gene set enrichment analysis (GSEA) of the differentially regulated genes in HG versus vehicle control (top panel) and HG versus HG + simvastatin (bottom panel) reveals that HG-upregulated EndMT process is blunted by simvastatin. c, Heatmaps of epithelial-mesenchymal-transition gene sets in HG versus vehicle control (left) and HG versus HG + simvastatin (right) based on GSEA analysis. Representative western blot data (d) and densitometric quantification (e) showing the hyperglycemic condition (HG) decreases phosphorylated/total YAP in iPSC-ECs compared to the control condition. GAPDH serves as a loading control (n = 3 biological samples). All data are presented as mean ± SEM. Unpaired two-sided Student’s t-test (a).
Extended Data Fig. 6 In vivo validation of the endothelial protective effects of simvastatin in a diabetic mouse model.
a, A representative trace of isometric tension in the mouse aorta. The aortic rings were equilibrated for 30 min under a resting tension of 10 mN after two sessions of pre-constriction with the vasoconstrictor PGF2α (1 µM). For a vasoconstriction response, endothelin-1 (ET-1, 0.1 nM to 1 µM) was used to induce a contractile response. At the plateau of maximal contraction, acetylcholine (Ach, 1 nM to 1 mM) was added accumulatively to initiate relaxation. b, Concentration-response relationship for ET-1-induced aortic constriction in mice treated with simvastatin (left panel) and GGTi298 (right panel). Developed tension was the force generated by aortic rings normalized to aortic tissue dry weight (mN/mg). Each point represents the mean developed contractile force ± SEM (n = 4 biological samples). c, Concentration-response relationship for ET-1-induced aortic constriction in wildtype mice treated with vehicle (saline), simvastatin (25 mg/kg), or GGTi298 (5 mg/kg) for 8 weeks (left panel). Concentration-response relationship for Ach-induced endothelial-dependent aortic relaxation in wildtype mice treated with vehicle (saline), simvastatin (25 mg/kg), or GGTi298 (5 mg/kg) for 8 weeks (right panel). Each point represents the mean constriction/relaxation response ± SEM (n = 4 biological samples). All data are presented as mean ± SEM.
Extended Data Fig. 7 RNA-seq analysis shows potent rescue effects of both simvastatin and GGTi298 on diabetes-induced vascular dysfunction in mice.
a, Venn diagram showing overlapped DEGs of db/+ (vehicle), db/db + simvastatin, and db/db + GGTi298 compared to the db/db + vehicle group, respectively. b, EndMT-associated genes, such as Ctgf, Vcam1, Tgfb2, and Smad3, were rescued by simvastatin and GGTi298 in diabetic mouse aortas (n = 2 RNA-seq biological samples). c, KEGG pathway analysis showing both simvastatin and GGTi298 improved genes associated with endothelial functions that were downregulated in db/db mouse aortas. d, Endothelial marker genes, such as Klf2, Nr2f2, and Klf5, were restored by simvastatin (simva) and GGTi298 in db/db mouse aortas (n = 2 RNA-seq biological samples). e, Representative immunofluorescent images showing YAP nuclear localization patterns in iPSC-ECs treated with disturbed flow, disturbed flow + simvastatin, laminar flow, and laminar flow + simvastatin. Blue: DAPI. Green: YAP. Scale bars: 50 μm. All data are presented as mean ± SEM.
Extended Data Fig. 8 Simvastatin-regulated enhancers identified by YAP ChIP-seq.
a, Western blot analysis showing simvastatin downregulated SOX9 expression in iPSC-ECs under disturbed and laminar flow patterns. b, Densitometric quantification of SOX9 protein expression changes in simvastatin- and vehicle-treated iPSC-ECs under disturbed and laminar flow patterns (n = 2 biological samples). c, Genome Browser snapshots of decreased binding loci at the NOTCH1 enhancer region showing simvastatin and GGTi298 treatment inhibited YAP binding at this region (highlighted in a yellow frame). d, Normalized gene expression levels (from RNA-seq) of Notch1 and Sox9 in aortas (n = 2 RNA-seq biological samples) from diabetic (db/db) and control mice (db/+). All data are presented as mean ± SEM. Unpaired two-sided Student’s t-test (b,d).
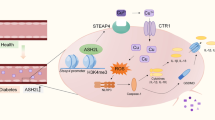
Extended Data Fig. 9 Schematic summary demonstrates that simvastatin rescues endothelial dysfunction by repressing YAP-mediated chromatin remodeling of the EndMT process.
A schematic overview showing our proposed model. In normal endothelial cells (ECs), geranylgeranyltransferase (GGTase) mediates YAP activity through the mevalonate pathway. Active YAP creates an ‘open chromatin status’ at the enhancer regions of genes regulating EndMT, such as SOX9. The diabetic condition further enhances YAP activity, thereby exacerbating endothelial functions by further upregulating EndMT genes. In contrast, statins decrease mevalonate levels via the inhibition of HMG-CoA reductase, followed by suppression of GGTase-mediated RhoA geranylgeranylation, consequently attenuating YAP activity (dashed line). Reduced YAP activity makes chromatin less-open (‘closed status’) at the enhancer regions of genes associated with EndMT. Diabetes-induced endothelial dysfunction can be alleviated by suppressing YAP activity with statins, GGTi298 or RhoA inhibitor (RhoAi), leading to the downregulation of EndMT genes. P, phosphorylation.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 2
Unprocessed blots.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 6
Unprocessed blots.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 5
Unprocessed blots.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 8
Unprocessed blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Liu, C., Shen, M., Tan, W.L.W. et al. Statins improve endothelial function via suppression of epigenetic-driven EndMT. Nat Cardiovasc Res 2, 467–485 (2023). https://doi.org/10.1038/s44161-023-00267-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s44161-023-00267-1
This article is cited by
-
Selenopeptide nanomedicine ameliorates atherosclerosis by reducing monocyte adhesions and inflammations
Nano Research (2024)
-
Rescue of EndMT-associated endothelial dysfunction by modulating the YAP pathway
Nature Cardiovascular Research (2023)
-
Simvastatin protects against endothelial dysfunction via epigenetic mechanisms
Nature Reviews Cardiology (2023)
