
Abstract
Leukocytes and endothelial cells frequently cooperate to resolve inflammatory events. In most cases, these interactions are transient in nature and triggered by immunological insults. Here, we report that, in areas of disturbed blood flow, aortic endothelial cells permanently and intimately associate with a population of specialized macrophages. These macrophages are recruited at birth from the closing ductus arteriosus and share the luminal surface with the endothelium, becoming interwoven in the tunica intima. Anatomical changes that affect hemodynamics, such as in patent ductus arteriosus, alter macrophage seeding to coincide with regions of disturbed flow. Aortic resident macrophages expand in situ via direct cell renewal. Induced depletion of intimal macrophages leads to thrombin-mediated endothelial cell contraction, progressive fibrin accumulation and formation of microthrombi that, once dislodged, cause blockade of vessels in several organs. Together the findings reveal that intravascular resident macrophages are essential to regulate thrombin activity and clear fibrin deposits in regions of disturbed blood flow.
Similar content being viewed by others
Main
As a gatekeeper of cellular traffic between blood and tissues, the endothelium is well equipped to interact with hematopoietic cells. Endothelial cells enable the well-coordinated process of diapedesis that includes capture, rolling and finally transmigration of leukocytes across the vascular barrier1,2,3,4. These events rely on complex and sequential molecular interactions by which these cell types cooperate to mount and resolve inflammatory responses at the level of venules and capillaries2,3. In large arteries, interactions of the endothelium with inflammatory cells are mostly known for their association with atherosclerosis, a chronic inflammation of the vascular wall5,6. In this pathology, leukocytes occupy the subendothelial layer, forming a neointima that progressively expands by the constant influx and local proliferation of inflammatory cells5,7,8,9.
Other types of interactions between arterial endothelium and inflammatory cells have been shown, particularly a population of non-classical monocytes (Ly6CloCCR2loCX3CR1hi) better referred to as patrolling monocytes that are thought to promote endothelial integrity and vascular health10,11,12. In addition to patrolling monocytes, myeloid cells with a highly dendritic appearance were also described in the luminal aspect of the aorta, especially at sites prone to develop atherosclerosis, such as the aortic arch13,14. Despite their conspicuous location, the contribution of these myeloid cells to atherosclerosis was proved to be minimal, as their elimination only slightly delayed onset of the disease with no impact on duration or burden15. Thus, the mechanisms behind their peculiar distribution, specific seeding time and, more importantly, their function have remained puzzling.
Here we show that the emergence of aortic myeloid cells is not pathologically induced; instead, it is developmentally triggered as part of natural hemodynamic changes at birth that result in localized disturbed flow dynamics. Genetic ablation of this aortic myeloid resident population promotes fibrin deposition and microthrombus formation, clarifying its function as a critical regulator of hemostasis.
Results
Hemodynamics at birth promote seeding of myeloid cells in the tunica intima of the aorta
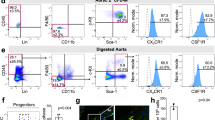
Fetal circulation includes two parallel circuits with equal left and right ventricular pressures. At birth, this balance changes drastically due to multiple concurrent events that include interruption of placental circulation, inflation of the lungs and a shift in pulmonary blood pressure. These changes result in high left ventricular pressure and closure of the ductus arteriosus, a fetal vessel that connects the pulmonary arteries to the aorta, an event that further magnifies oscillatory flow in the lower curvature of the aortic arch (Fig. 1a). These alterations in hemodynamics are quickly sensed by endothelial cells, which transition from an elongated to a polygonal shape in the lesser curvature of the aortic arch (Fig. 1b,c). Furthermore, evaluation of mouse embryos and neonates uncovered a burst of inflammatory cells exiting from the constricted ductus arteriosus that seeds the aorta, in tandem with the changes in hemodynamics (Fig. 1d–f). Interestingly, from the onset of birth, this population of CD45+ cells continued to reside in areas experiencing oscillatory and disturbed flow, including the lesser curvature of the aortic arch and branch openings (Fig.1g–i and Extended Data Fig. 1a). Initial characterization indicated that they also expressed CD11c (Fig. 1h,j). Curiously, CD11c+ cells were previously detected in the aortic arch of adult healthy mice13,14, raising the possibility that they might be the same population. We also found that CD11c+ cells progressively accumulated with age in the absence of pathologies or hypercholesterolemia (Extended Data Fig. 1a–c). However, they were not found in large veins, such as the vena cava (Extended Data Fig. 1d,e), indicating that an arterial niche, including flow patterns, may be required for their accumulation. Intimal immune cells were also absent from the carotid arteries of healthy, adult mice (Extended Data Fig. 1f) and from the descending young aorta, except for branches (Extended Data Fig. 1g). We also observed that the distribution of these cells progressively broadened with age. In fact, the descending aorta of 52- and 78-week-old mice revealed ongoing accumulation of intimal immune cells even in areas of laminar flow (Extended Data Fig. 1g–i), suggesting that vascular aging might also be a supportive niche for the seeding of intimal myeloid cells.

a, Embryonic day (E)18.5, connection between pulmonary arteries and aorta through the ductus arteriosus. Postnatal day (P)1, the ductus arteriosus constricts, contributing to disturbed flow. Eight weeks, independent aorta and pulmonary artery. Red, high oxygen levels; blue, low oxygen levels; purple, intermediate oxygen levels. b, Schema illustrating aorta dissection. GC, greater curvature; LC, lesser curvature; desc., descending aorta. c, Confocal images of the lesser curvature and descending aorta from mice at E18.5, P7 and 8 weeks. Elongation factor in the lesser curvature is on the adjacent graph (E18.5, n = 3; P7, n = 4; 8 weeks, n = 5 mice), Mann–Whitney t-test, mean ± s.d., two tailed, P < 0.0001 (exact), ****P ≤ 0.0001; scale bar, 10 μm). d, Bright-field images of ductus arteriosus (DA) remodeling (scale bars, 150 μm, E18.5 (n = 8 embryos); 200 μm, P1 (n = 10 mice); 500 μm, P3 (n = 12 mice); and 1 mm, 3 weeks (n = 12 mice)). e, Exit of immune cells (green) from the ductus arteriosus. VE-Cad or the transcription factor ERG are indicated in red. Scale bars, 300 μm and 70 μm (e(i–iv)); n = 12 mice per time point. f, Lumen of the ductus arteriosus (dotted white lines) at P0; immune cells are shown in green. Scale bar, 50 μm; n = 5 mice. g, Illustration indicating regions of disturbed flow where intimal immune cells (green) accumulate. Branch openings (dashed boxes) are shown in h,i. h, Large branch openings with accumulation of intimal myeloid cells (CD11c, red; CD45, green) (white arrows). ERG staining (white) identifies endothelial nuclei. BA, brachiocephalic trunk; LCCA, left common carotid artery; LSA, left subclavian artery. Scale bars, 200 μm (left) and 20 μm (right); n = 15 mice. i, Time course (1 week, 8 weeks, 78 weeks) of intimal CD45+ cell (green) deposition in intercostal arteries (yellow arrows). VE-Cad, red; intimal CD45+ cells, green. Scale bar, 50 μm; n = 12 (1 week), n = 498 (8 weeks), n = 6 (78 weeks) mice. j, In total, 99.4% of intimal CD45+ cells in the lesser curvature are also labeled as CD11c+ (n = 3 mice, mean ± s.d.). k, Bright-field images of control and Vim−/− aortae. Insets show the remnant (control) and PDA (Vim−/−) with blood (n = 8 mice per group; scale bar, 1,300 μm). l, Aortic arch of 10-week-old control and Vim−/− mice; immune cells in control (green arrows) and Vim−/− (dotted pink ovals) mice are shown. Right, high magnification of squares (l(i,ii)). Scale bars, 500 μm and 40 μm (l(i,ii)); n = 3 mice per group.
A definitive link between onset of oscillatory flow and recruitment of CD11c+ cells was established using mouse models of patent ductus arteriosus (PDA). Failure in PDA closure significantly alters cardiovascular hemodynamics16. While viable and fertile17, Vimentin−/− (Vim−/−) mice exhibit PDA in about 88% of adults, making them an ideal model to study myeloid cell distribution in adult aortae18. At 10 weeks of age, the ductus arteriosus in wild-type (WT) littermates becomes a solid fibrous structure that persists as the ligamentum arteriosum (Fig. 1k). By contrast, Vim−/− mice exhibit a viable ductus arteriosus, which impacts patterns of disturbed flow and the location of myeloid cells (Fig. 1l). The lesser curvature of the aortic arch in Vim−/− mice showed no intimal myeloid accumulation; instead, myeloid cells were noted, surrounding the openings of the ductus arteriosus, the subclavian artery and onset of the descending aorta (Fig. 1l).
The thin nature of the endothelial lining makes it difficult to ascertain the precise topology of myeloid cells in relation to the endothelium. Using Cdh5CreERT2;R26tdTomato reporter mice to label the endothelial monolayer (tdTomato combined with CD45 and vascular endothelial cadherin (VE-Cad) staining), we generated a three-dimensional (3D) rendering of the tunica intima. Myeloid cells were neither above nor below the endothelium but were instead interwoven within the endothelial monolayer, with cell processes projecting into and others below the lumen (Fig. 2a). En face scanning electron microscopy (SEM) of the aortic arch stained with anti-CD45 antibodies confirmed these findings (Fig. 2b). Additionally, 3D surface rendering using a reporter model that labels the tunica media using the Tagln (Sm22) promoter (Sm22Cre;R26tdTomato) combined with VE-Cad immunohistochemistry further validated that these CD45+ cells were located in the tunica intima (Fig. 2c). Finally, myeloid cells were detected by intravascular injection of anti-CD45 antibodies in vivo (Fig. 2d,e).

a, Cdh5CreERT2;R26tdTomato construct of the double transgenic mice used for 3D surface rendering of adult aortae to visualize the spatial location of intimal CD45+ cells (green) relative to the endothelium (tdTomato (tdTom)). VE-Cad is shown in white. Scale bars, 8 μm; n = 3 mice. Max-int. proj., maximum-intensity projection. b, Confocal and scanning electron microscopy (SEM) images of the same aorta were overlaid to determine the location of intimal CD45+ cells (green in the confocal view) in relation to the endothelium. ERG was used to visualize endothelial nuclei in confocal images. High-vac., high-vaccume. Scale bar, 10 μm; n = 3 mice. c, Sm22Cre;R26tdTomato construct of the double transgenic mice used for 3D rendering. Side view of a 3D surface image of adult aortae to visualize the spatial location of intimal CD45+ cells (green) relative to vascular smooth muscle cells (tdTom). Endothelial cells were visualized with anti-VE-Cad antibodies (white). Note that the green CD45+ cell is embedded in an area with white staining (VE-Cad). Scale bars, 5 μm; n = 3 mice. d, Schema of the experimental design related to e,g. Inj., injection; i.v., intravenous. e, Tail vein injection of rat anti-CD45 antibody (detected in green) was followed by euthanasia and fixation of the aorta 30 min later. Subsequent permeabilization and staining with an additional, distinct anti-CD45 antibody (detected using far red, labeled in white). The illustration was created using https://biorender.com/. e, En face images of the experiment described. Yellow arrowheads indicate the portions of immune cells exposed to the lumen (green). Mouse anti-CD45 biotinylated antibody labeled with streptavidin–A647 used in immunostaining after permeabilization is shown in white. PECAM1, red; 4,6-diamidino-2-phenylindole (DAPI), blue. Scale bar, 5 μm; n = 3 mice. f, En face images of the lesser curvature, staining for PECAM1 (green). Yellow arrowheads show faint PECAM1 positivity in CD45+ cells residing in the intima. Endothelial cells were detected in green with PECAM1. Scale bar, 5 μm; n = 6 mice. g, C57BL/6 mice were injected intravenously with a non-blocking PECAM1-specific (390) antibody to further examine exposure of cell bodies to the lumen. Mice were killed and harvested 15–30 min post-injection. 15 min after injection. CD45 (in green) was used to identify immune cells, and ERG (red) was used to identify endothelial cells. Yellow arrowheads indicate PECAM1+ regions of intimal immune cells. Scale bar, 4 μm; n = 3 mice.
During diapedesis, leukocytes adhere to endothelial cells through platelet endothelial cell adhesion molecule (PECAM1), preserving junctional integrity as they cross the endothelial barrier2,3,4. We also detected PECAM1 expression in these myeloid cells, indicating that they bind to the endothelium via homophylic, heterotypic interactions and prevent barrier disruption (Fig. 2f). En face images after injection of non-blocking PECAM1-specific antibodies revealed lumen-exposed regions in inflammatory cells (Fig. 2g). When combined, these experiments revealed that, in regions of disturbed flow, the constituency of the endothelial layer is enriched by a population of myeloid cells that intimately coexists with the endothelium in the absence of pathology and without breach of permeability.
scRNA-seq reveals the transcriptional identity of aortic intimal immune cells
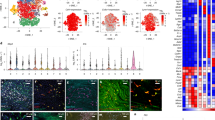
To recover cellular identities independently of defined labeling strategies, we turned to single-cell RNA sequencing (scRNA-seq) using specimens that were not sorted by flow cytometry19,20 (Extended Data Fig. 2a). Three independent scRNA-seq libraries were generated, capitalizing on regions with abundant numbers of intimal immune cells: the aortic arch of 8-week-old C57BL/6 mice (‘young arch’) and the descending aorta (thoracic and abdominal) of 78-week-old C57BL/6 mice (‘aged descending A,B’) (Extended Data Fig. 2b–e and Supplementary Table 1). Using dimensionality reduction by t-distributed stochastic neighbor embedding (t-SNE) analysis, we identified ten distinct cell types and assigned cellular identities (Extended Data Fig. 2f and Supplementary Tables 2,3) based on canonical lineage markers (Extended Data Fig. 2g,h). Within the ten cell types, two distinct macrophage populations were identified based on Fcgr1 (CD64) and Adgre1 (F4/80) expression (Extended Data Fig. 2h). One of the macrophage clusters expressed Lyve1 (encoding lymphatic endothelium hyaluronan receptor 1 (LYVE1)), F13a1 and Mrc1 (CD206), well-known markers of adventitia macrophages15,21,22,23,24 (Extended Data Fig. 2i,j), suggesting that this macrophage population came from the adventitia. While our isolation method enriches for cells in the tunica intima, few cells from the other aortic layers (adventitia and media) were also captured19,20 (Extended Data Fig. 2f,g). Therefore, we predicted that one of the two macrophage populations identified was from the adventitia. To test this prediction, we performed single-cell sequencing of dissected aortic adventitia (Extended Data Fig. 3a–g) and compared the two distinct myeloid populations identified. For this, we selected CD14-positive cells from the purified adventitia and intimal-enriched libraries (Extended Data Fig. 3c–e). This approach definitively confirmed the identity of the macrophage clusters from the aorta libraries. The Lyve1+Fcgr1+ group, also present in libraries from the adventitia, represented typical adventitial macrophages, while the second cluster with distinct expression of Mmp12 and Mmp13 was unique to intima-enriched aortic libraries and absent from the adventitia (Extended Data Fig. 3f,g). Furthermore, we compared the Mmp12+Mmp13+ aortic macrophage population to macrophages from the Tabula Muris Atlas25 (Extended Data Fig. 4a–h) and resident macrophages characterized in more recent publications22 (Extended Data Fig. 4i). From this analysis, it became clear that the aortic population of intimal macrophages was especially distinct from resident macrophages in other organs, with the exception of sympathetic nerve-associated macrophages found in the lung22.
Subsequently, we compared the Mmp12+ Mmp13+ macrophage population to a recently identified group of macrophages isolated from whole aortas and referred to as MacAIR cells15. Comparisons between the intimal Mmp12+Mmp13+ population presented here to the pre-hypercholesterolemic aortic MacAIR population15 revealed that they were transcriptionally identical (Supplementary Table 4). Interestingly, we also found that the young arch and old thoracic endothelial-associated macrophage populations were identical (Extended Data Fig. 4j and Supplementary Table 5), suggesting that the endothelial niche is responsible for the underlying tissue-specific imprinting of these macrophages. This realization prompted two immediate questions: were these populations progeny of CD45+ cells exiting the ductus arteriosus? And, more importantly, what was their biological relevance?
Aortic intima resident macrophages (MacAIR) are distinct macrophages that seed the aorta at birth
To clarify the function of MacAIR cells and delve into their developmental origin, we first performed differential expression analysis to seek as many unique markers as possible. As shown by direct comparisons with adventitial macrophages and the Tabula Muris Atlas, we found that MacAIR cells expressed significantly higher transcriptional levels of matrix metalloproteinases (MMPs) (Mmp12 and Mmp13; Extended Data Fig. 2i,j) and exhibited increased expression of immune cell activation genes (Il1b, Ccl3, Ccl4, Tnf, Cxcl2 and Cxcl16; Extended Data Fig. 2i,j and Supplementary Tables 6–8). In addition, MacAIR cells displayed expression of genes involved with wound healing or identification of dying cells (Axl, Tyrobp, Cd44 and Cd74; Extended Data Fig. 2i,j and Supplementary Tables 6–8) and showed higher expression levels of antigen-presenting genes, such as major histocompatibility complex class II (MHC class II)-encoding genes (H2-Aa, H2-Ab1, H2-Eb1 and H2-M2; Extended Data Fig. 2i,j and Supplementary Tables 6–8). By contrast, adventitia Lyve1hi macrophages expressed genes associated with M2-like macrophages (F13a1, Folr2 and Mrc1 (CD206); Extended Data Fig. 2i,j and Supplementary Tables 7 and 8).
To match the expression profile to their presumed progenitors, immunohistochemistry was performed on aortae of mice at postnatal day (P)7 and adult mice for targets unique to MacAIR cells (MMP13, CXCL16 and CD11c). Intimal CD45+ cells colocalized with MacAIR markers (Extended Data Fig. 5a,b); whereas, in the adventitia, no CD45+ cells showed expression of MacAIR markers (Extended Data Fig. 5c). We also found that MacAIR cells expressed Cx3cr1 transcripts (Extended Data Fig. 5d) and CX3CR1 (fractalkine receptor) protein (Extended Data Fig. 5e). Moreover, using an inducible Cx3cr1 reporter model (Cx3cr1CreERT2;R26tdTomato), 98% of all intimal CD45+ cells in adult aortae were labeled with tdTomato (Extended Data Fig. 5f), which also colocalized with CXCL16 (Extended Data Fig. 5g), thus demonstrating the activity of the Cx3cr1 promoter, which was later used for lineage tracing. Additionally, using this transgene, intimal CD45+ cells in aortae of mice at P5 were also labeled by the reporter after tamoxifen treatment (P1 and P3), further indicating that MacAIR cells seed the aorta immediately after birth (Extended Data Fig. 5h). Overall, these findings support the conclusion that immune cells accumulating and residing in the tunica intima after birth were MacAIR cells.
To more definitively confirm the origin and the macrophage identity of MacAIR cells (versus dendritic cells (DCs)), we assessed recombination labeling using the Csf1rMerCreMer;R26tdTomato macrophage fate-mapping model. In this transgenic model, we found all intimal immune cells (CD45+) to also be positive for tdTomato (Extended Data Fig. 5i). Moreover, all intimal immune cells expressed CD68 (Extended Data Fig. 5j). Additionally, MacAIR cells did not express the DC master regulator transcription factor encoded by Zbtb46 (ref. 26) nor classical DC markers Cd8a, Ccr7 (Extended Data Fig. 5k), Cd103, Dcir2, or Mycl (not detected). Furthermore, MacAIR cells were shown to phagocytose dying (Annexin+) endothelial cells in vivo (Extended Data Fig. 5l), providing functional evidence of their macrophage identity. Thus, MacAIR cells are a transcriptionally unique macrophage population that takes residency in the tunica intima of the aorta shortly after birth in regions of disturbed flow.
MacAIR cells are self-maintained throughout adulthood
It is now understood that many tissue-resident macrophages are maintained through self-renewal without contribution from circulating monocytes27,28. Incorporation of 5-ethynyl-2-deoxyuridine (EdU) demonstrated that, in a 2-h pulse, 5.4% of MacAIR cells were undergoing DNA replication in situ in the adult arch (Extended Data Fig. 6a–c), suggesting that MacAIR cells followed the trend of other resident macrophages. Long-term replenishment was assessed by lineage tracing with pulse-labeling Cx3cr1CreERT2;R26tdTomato mice and evaluation 10 and 20 weeks after labeling (Extended Data Fig. 6d,e). To control for the possible contribution of circulating cells, we also quantified tdTomato-positive blood cells. Flow cytometry analyses indicated that, after 3 weeks, CD45+tdTomato+ peripheral blood was negligible (less than 0.5%; Extended Data Fig. 6f,g), thus negating the possibility that cells from the circulation contributed to reporter positive MacAIR cells 20 weeks after tamoxifen treatment. In this manner, the absence of circulating tdTomato+ cells allowed us to ask whether monocytes participated in replenishment of MacAIR cells. Although the labeling frequency of MacAIR cells decreased by 10% 10 weeks after tamoxifen injection, we noted that reporter expression remained constant for an additional 10 weeks (20 weeks in total) (Extended Data Fig. 6e,h). This finding supports the conclusion of local self-renewal with negligible contribution from monocytes.
To further confirm these findings, we performed parabiosis experiments using adult GFP+ and WT mice in which both mice shared chimeric circulation for 5 weeks (Extended Data Fig. 6i). The experiment allowed all circulating cells to access the aortic regions of interest (lesser curvature). The presence of GFP+ intimal immune cells incorporated in WT mice would infer monocyte contribution and vice versa (Extended Data Fig. 6i). Only mice with excellent level of parabiosis were used (Extended Data Fig. 6j). Evaluation of the lesser curvature in WT mice revealed no GFP+ intimal immune cells (Extended Data Fig. 6k), proving further strength to the conclusion that MacAIR cell maintenance was independent of monocytes. This was also supported by clonal analysis of Cx3cr1CreERT2;R26Rainbow mice in which inducible Cre recombinase in cells expressing Cx3cr1 would randomly recombine, generating three fluorescently labeled populations (Extended Data Fig. 6l). The data strongly support the notion that MacAIR cells are self-maintained.
MacAIR cells originate from definitive hematopoietic cells and expand through self-renewal
MacAIR cells colonize the aorta as they exit from the ductus arteriosus and migrate to areas of disturbed flow. After this initial wave of migration, MacAIR cells expand in number by local proliferation (Fig. 3a). Nonetheless, lineage tracing was required to fully ascertain adult progeny. Thus, we performed lineage analysis using the inducible Cx3cr1CreERT2;R26tdTomato reporter model to label MacAIR cells immediately after birth and follow descendants over time (Fig. 3b,c). After administration of tamoxifen at birth, 63% of intimal CD45+ cells were positive for tdTomato by P7. At 4 weeks, 91% of intimal CD45+ cells in aortae retained tdTomato labeling (Fig. 3d–f). We interpret the increase from P7 (63%) to 4 weeks (91%) to indicate that only those inflammatory cells expressing Cx3cr1 were successfully retained in the endothelium. Importantly, at 4 weeks, less than 0.5% of peripheral blood cells were positive for tdTomato (Fig. 3f), supporting the absence of contributions from circulating cells. Therefore, these data indicate that the MacAIR cells that enter the aorta postnatally expand via self-renewal and colonize the lesser curvature with minimal (if any) input from circulating monocytes.

a, Experimental design. EdU injection at P7 2 h before euthanasia. Intimal CD45+ cells (green) proliferate (EdU+, white) in situ (yellow arrowheads). Endothelial cells are shown in red (ERG). Scale bar, 20 μm; n = 3 mice. b, For lineage tracing, Cx3cr1CreERT2;R26tdTomato neonates were injected with tamoxifen (tam) at P1, P3 and P5. At P7, aortae were collected from half of the litter as a baseline control. At 4 weeks, aortae were collected from the remaining littermates for retention of the reporter in MacAIR cells. c, Illustration depicting possible outcomes. tdTomato levels are negligible in blood (as shown in g). PB, peripheral blood. d, tdTomato+ immune cells in aortae at P7 (top) and 4 weeks (bottom) of Cx3cr1CreERT2;R26tdTomato mice after tamoxifen administration. MacAIR cells in aortae of Cx3cr1CreERT2;R26tdTomato mice at 4 weeks retained reporter (tdTomato+) expression when labeled postnatally, as shown by the comparison between P7 (baseline) and 4 weeks (lineage traced). CD45, green; VE-Cad, white. Scale bars, 80 μm and 10 μm, n = 5 (P7); 300 μm and 20 μm, n = 4 (4-week-old) mice. e, Percentage of intimal tdTomato+CD45+ cells in aortae of Cx3cr1CreERT2;R26tdTomato mice at P7 and 4 weeks when labeled with tamoxifen postnatally (n = 4 (4-week-old), n = 5 (P7) mice; Mann–Whitney t-test, P = 0.0159, two tailed, mean ± s.d., *P ≤ 0.05). f, Flow cytometry plots of tdTomato labeling of circulating cells at P7 and 4 weeks (n = 3 (4-week-old), n = 4 (P7) mice). g, Experimental design for the clonal tracing model and possible outcomes. Tamoxifen injection of Cx3cr1CreERT2;R26Rainbow mice at P1, P3 and P5. Aortae were collected at 8 weeks. Cer., Cerulean; mOr., mOrange; mCh., mCherry. h, Clones of MacAIR cells labeled at 8 weeks (initial tamoxifen injections at P1, P3 and P5). CD45+ cells are shown in white. Scale bar, 100 μm; n = 5 mice. i, Intimal CD11c+CD45+ cells from control and Ccr2−/− mice at P5 (top, former ductus arteriosus region) and 8 weeks (bottom, lesser curvature). CD11c, red; CD45, green; ERG, white. Scale bar, 100 μm and 50 μm; n = 7 mice per time point. j, Comparison of CD11c+CD45+ cells in aortae from control and Ccr2−/− mice at P5 (n = 6 (control), n = 7 (Ccr2−/−) mice; Mann–Whitney t-test, mean ± s.d., P > 0.9999 (exact), two tailed) or in aortae from mice at 8 weeks (n = 7 (control) or n = 6 (Ccr2−/−) mice; Mann–Whitney t-test, mean ± s.d., P = 0.0872, two tailed). NS, not significant. k, Summary of Figs. 1–3. MacAIR cells enter the aorta via the ductus arteriosus, expand through self-renewal and populate the lesser curvature. Macrophages also accumulate in descending aortae from aged mice. Illustrations (a,b,g) were created using https://biorender.com/.
To assess clonal expansion of MacAIR cells at early time points, we again used the inducible Cx3cr1CreERT2;R26Rainbow model (Fig. 3g). Transgenic pups were treated with tamoxifen immediately after birth to induce stochastic recombination, yielding permanent expression of one of three mutually exclusive fluorescent proteins: Cerulean, mOrange and mCherry (Fig. 3g). At 8 weeks of age, clones of labeled MacAIR cells were observed (Fig. 3h), confirming that MacAIR cells self-expand in situ after seeding the aorta immediately after birth. We also examined MacAIR cells in chemokine receptor 2 (CCR2)-deficient (Ccr2−/−) mice, in which Ly6Chi (classical) monocyte emigration from the bone marrow is defective29. Compared to control mice, we found no difference in the proportion and number of intimal CD11c+CD45+ (MacAIR) cells in the aortae of either young (P7) or adult (8-week-old) mice (Fig. 3i,j). This further solidifies the notion that postnatal aortic intimal macrophages self-expand and maintain their population independently of monocytes at steady state (Fig. 3k).
Assessment of whether MacAIR cells were derived from primitive or definitive hematopoietic lineages was determined through experiments with a defined lineage-tracing model (Flt3Cre;R26mTmG) that labels cells arising from definitive hematopoietic stem cells as GFP+ (refs. 27,30,31) (Extended Data Fig. 6m). Aortae from Flt3Cre;R26mTmG mice at P5 showed that a large proportion of intimal CD45+ cells were GFP+ relative to circulating CD11bhiGr1lo monocytes (Extended Data Fig. 6n,o). This labeling, combined with the observation that MacAIRs were detected post-birth, strongly indicates that these cells were derived from a definitive hematopoietic lineage and not from the yolk sac, as is known to occur for some resident macrophage populations27,28.
MacAIR cells blunt thrombin activity in areas of oscillatory flow
An inducible diphtheria toxin (DTx) model (Cx3cr1CreERT2;Csf1rlslDTR) in which dual tamoxifen and DTx injections result in elimination of cells expressing both Cx3cr1 and Csf1r was used to deplete MacAIR cells and clarify their biological relevance (Fig. 4a). Efficient loss of MacAIR cells was noted 24 h after DTx injection (Fig. 4b) and was associated with altered endothelial morphology and apparent reduction in cell size (Fig. 4c and Extended Data Fig. 7a). To clarify whether cell size changes were due to cell contraction, we evaluated the expression of phosphorylated myosin light chain 2 (pMLC2). In control mice, MacAIR cells showed high expression of pMLC2 (Fig. 4d and Extended Data Fig. 7b). Upon MacAIR depletion, endothelial cells became positive for pMCL2, indicating a highly contractile functional state, likely the reason behind the drastic change in cell size (Fig. 4c,d and Extended Data Fig. 7b). Together, these findings suggested that MacAIR cells prevented a contractile endothelial phenotype that would otherwise manifest in areas of disturbed flow through an unclear mechanism.

a, Transgenes (Cx3cr1CreERT2;Csf1rlslDTR) used to deplete MacAIR cells require both tamoxifen and DTx to promote apoptosis of double-positive cells. DTR, diptheria toxin receptor; DTx, diptheria toxin. b, Images and quantification of intimal CD45+ cells in the lesser curvature from controls and Cx3cr1CreERT2;Csf1rlslDTR mice (both exposed for 24 h to tamoxifen and DTx). CD45+ (green) cells were effectively depleted. Scale bar, 50 μm; n = 5 mice per group, Mann–Whitney t-test, mean ± s.d., P = 0.0079, two tailed, **P ≤ 0.01. c, Surface area and elongation factor measurements of endothelial cells in the lesser curvature of Cx3cr1CreERT2;Csf1rlslDTR mice and littermate controls after MacAIR depletion (n = 3 (control) or n = 4 (Cx3cr1CreERT2;Csf1rlslDTR) mice; Mann–Whitney t-test, line at median, P < 0.0001 (exact, surface area and elongation factor), two tailed, ****P ≤ 0.0001). d, pMLC2 staining (red) in the lesser curvature 1 d after tamoxifen and DTx injections of control and Cx3cr1CreERT2;Csf1rlslDTR mice. CD45, green; VE-Cad, white. Scale bar, 10 μm; n = 5 mice per group. e, Images of co-staining for MMP12 or MMP13 (red), CD45 (green) and VE-Cad (white). Note the colocalization of MMP12 and MMP13 with CD45+ cells. Scale bars, 20 μm and 10 μm; n = 3 mice. f, Thrombin in areas of oscillatory and disturbed flow in aortae of 8-week-old C57BL/6 mice (IC, intercostal artery); scale bar, 20 μm; n = 3 mice. g–i, Effects of dabigatran on aortae from Cx3cr1CreERT2;Csf1rlslDTR and control mice. g, Experimental design. Cx3cr1CreERT2;Csf1rlslDTR and littermate controls treated with tamoxifen and DTx for depletion of MacAIR cells. Vehicle or dabigatran was administered for 5 d. The illustration was created using https://biorender.com/. h, Images of the lesser curvature 5 d after treatments as indicated. pMCL2, red; CD45, green; VE-Cad, white. Loss of pMLC2 in dabigatran-treated Cx3cr1CreERT2;Csf1rlslDTR MacAIR-depleted mice. Scale bar, 7 μm; n = 5 mice per group. i, Surface area measurements of endothelial cells in the lesser curvature of Cx3cr1CreERT2;Csf1rlslDTR mice and littermate controls 5 d after dabigatran treatment (n = 7 mice per group, Mann–Whitney t-test, line at median, P < 0.0001 (exact), two tailed, ****P ≤ 0.0001). j, Western blot of human aortic endothelial cells (HAECs) treated with thrombin or MMP13 or pretreated with MMP13 (time course), followed by thrombin treatment for an additional 5 min (n = 4 biological replicates, Mann–Whitney t-test, mean ± s.d., P = 0.0286 (vehicle versus thrombin; vehicle versus MMP13, 5 min; MMP13 versus thrombin; thrombin versus MMP13; thrombin versus MMP13, 1 min; thrombin versus MMP13, 5 min), two tailed, *P ≤ 0.05).
It is well known that exposure of cultured endothelial cells to thrombin results in rapid Rho activation, phosphorylation of myosin light chain 2 (MLC2), stress fiber formation, contraction and disruption of endothelial junctions32,33 (Extended Data Fig. 7c). These events trigger downstream signaling by GPCR protease-activated receptor (PAR)1, a thrombin receptor highly expressed by aortic endothelial cells (Extended Data Fig. 7d) and activated upon cleavage32,33. Interestingly, plasmin and other proteases, including MMP12 and MMP13 (both highly expressed by MacAIR cells) (Fig. 4e), can alter this signaling pathway34,35,36,37. Specifically, MMP13 can cleave PAR receptors on the carboxyl-terminal side of the thrombin cleavage site and thus prevent thrombin-mediated endothelial contraction34,36,37. Consequently, we posit that, in absence of MacAIR cells, intact PAR1 could be activated by thrombin, leading to endothelial contraction. This assumption implied accumulation of thrombin in areas of disturbed flow. Indeed, immunocytochemistry revealed thrombin deposits at the lesser curvature and branch openings (Fig. 4f). Additionally, to further confirm thrombin activity in regions of disturbed flow, we treated control and Cx3cr1CreERT2;Csf1rlslDTR mice with vehicle or dabigatran (thrombin inhibitor) and assessed endothelial cell contraction (Fig. 4g). Five days after treatment, dabigatran-treated control mice still expressed pMLC2 in MacAIR cells, indicating that macrophage contraction was thrombin independent (Fig. 4h). However, depletion of MacAIR cells with concurrent dabigatran treatment prevented endothelial cell contraction, as shown by pMLC2 expression and cell size quantification (Fig. 4h,i), demonstrating that endothelial contraction was thrombin dependent but mitigated by MacAIR cells. Thrombin was still present in areas of oscillatory flow after macrophage depletion (Extended Data Fig. 7e). These findings indicate that MacAIR cells prevent endothelial contraction driven by thrombin in regions of disturbed flow. Importantly, we tested other aspects of clotting in the presence and absence of macrophages, including tail bleed times (Extended Data Fig. 7f) as well as other mediators or regulators of clotting such as nitric oxide and prostacyclin (Extended Data Fig. 7g,h). None of these were altered by depletion of MacAIR cells.
The next critical question was to sort out the molecular mechanism by which MacAIR cells impaired thrombin action and, in particular, pMLC2. It is well established that thrombin mediates signals on endothelial cells through PAR1, PAR2 and PAR4)32,33, all expressed by aortic endothelial cells but not by MacAIR cells (Extended Data Fig. 7d). Interestingly, PAR receptors can be cleaved by thrombin, mediating signaling, but also by other proteases such as MMP12 and MMP13 blunting these signals by cleaving PAR1 on the carboxyl-terminal side of the thrombin site34,35,36,37. Considering the high levels of MMP12 and MMP13 expressed by MacAIR cells, it was only logical to predict that MMPs secreted by MacAIR cells impaired thrombin-mediated contraction. While testing this hypothesis in mouse aortae was impossible, we evaluated thrombin signaling, as shown by pMLC2 levels on cultured endothelial cells in the presence and absence of MMP13 (Fig. 4j). The findings showed that thrombin activates pMLC2 in the endothelium and that this effect is impaired by co-incubation with MMP13.
MacAIR residency coincides with deposition of fibrin(ogen)
The presence of thrombin in areas of disturbed flow led us to inquire whether fibrinogen, a thrombin substrate, was also found in these regions. Indeed, en face immunostaining revealed buildup of fibrin(ogen) in the lesser curvature (Fig. 5a). Higher magnification images (Fig. 5b) and 3D surface rendering (Fig. 5c) showed fibrin(ogen) decorating the surface of MacAIR cells. To confirm that fibrin(ogen) was deposited in regions of disturbed flow, we depleted endogenous fibrinogen in vivo and mice with fluorescently tagged fibrinogen to assess binding and accumulation (Fig. 5d). Knockdown of Fbg (fibrinogen gamma chain) was accomplished by delivery of small interfering (si)RNA targeting hepatic fibrinogen mRNA (siFibrinogen), which was encapsulated in lipid nanoparticles containing an ionizable cationic lipid38. Seven days after treatment, circulating fibrinogen was ~90% depleted (Fig. 5e), and, at this time, fibrinogen–Alexa Fluor (A)488 (fbg–a488) avidly bound to MacAIR cells and accumulated throughout the lesser curvature 3 h after injection (Fig. 5f). By contrast, fbg–a488 did not accumulate in the greater curvature (Extended Data Fig. 8a), confirming the predilection of fibrinogen for regions of disturbed flow.

a, Whole-mount, en face images of fibrin(ogen) (white) and CD45+ cells (green) in the aortic arch of 8-week-old C57BL/6 mice. Scale bars, 500 μm and 50 μm; n = 20 mice. b, Fibrin(ogen) (white) was overlaid with CD45 staining (green) in the adult aortic arch. Higher magnification (bottom) of blue dotted inserts highlights fibrin(ogen) staining in an area that resembles the cellular process of a MacAIR cell (blue dotted ellipse). Scale bars, 10 μM and 5 μM; n = 20 mice. c, En face images of fibrin(ogen) location in the aortic arch displayed using maximum-intensity projections (top) and 3D surface rendering (bottom). Scale bars, 7 μm and 4 μm; n = 16 mice. d, Experimental design. Eight-week-old C57BL/6 mice were injected via the tail vein with either siFibrinogen (siFbg) or siLuciferase (siLuc) (control). Seven days later, mice were injected via the tail vein with fbg–a488. Three hours after fbg–a488 injection, aortae were collected for analysis. The illustration was created using https://biorender.com/. e, Plasma levels of fibrinogen 7 d after mRNA injection, measured by ELISA (n = 4 mice per group, Mann–Whitney t-test, mean ± s.d., P = 0.0286, two tailed, *P ≤ 0.05). f, Distribution of intravenously injected fbg–a488 in relation to MacAIR cells. CD45 is shown in white. Yellow arrowheads indicate MacAIR surface-bound fbg–a488 (n = 4 mice per group; scale bar, 4 μm). g, Intercostal artery from adult (8-week-old) mice exhibiting fibrin(ogen) deposition (white) and intimal CD45+ cell accumulation (green). VE-Cad (red) identifies endothelial cells (scale bar, 70 μm, n = 15 mice). h, En face images of the aortic arch comparing fibrin(ogen) accumulation (green) and immune cell (CD45, white) deposition in control (h(i); n = 8 mice), Fbg−/− (h(ii); n = 5 mice) and FbgAEK (h(iii); n = 6 mice) mice. VE-Cad identifies endothelial cells in red. Scale bars, 300 μm and 8 μm. The adjacent graph represents quantification of MacAIR cells in the lesser curvature of the aortic arch in Fbg−/− and FbgAEK mice compared to colony controls (n = 3 (FbgWT/WT), n = 5 (Fbg−/−), n = 5 (FbgWT/WT), n = 6 (FbgAEK) mice per group; Mann–Whitney t-test, mean ± s.d., P = 0.3929, P = 0.6623, two tailed; P > 0.05 is not significant).
We validated concurrent associations of MacAIR cells and fibrin(ogen) in intercostal arterial openings (Fig. 5g). The correlation was also noted in descending aortae of 78-week-old mice (Extended Data Fig. 8b). Moreover, a time course evaluation showed progressive fibrin(ogen) accumulation in the aortic arch of mice at P7, 3 weeks and 8 weeks, coinciding with the age-dependent expansion of MacAIR cells (Extended Data Fig. 8c). As fibrinogen is a substrate for macrophages, we predicted that fibrinogen deposits might be required for seeding MacAIR cells in the regions of disturbed flow. Thus, we analyzed MacAIR accumulation in fibrinogen-deficient mice (Fbg−/−)39 and in mice expressing a mutant form of fibrinogen that could not be converted to fibrin polymer (FbgAEK)40. Our prediction was incorrect, as no difference in the abundance or distribution of MacAIR cells was found in either mouse model (Fig. 5h). These findings revealed that, while overlapping in location, fibrin(ogen) was dispensable for seeding or anchoring MacAIR cells to the tunica intima.
MacAIR cells are necessary to clear fibrin(ogen) deposits in regions of disturbed flow
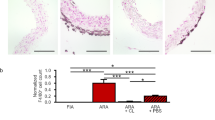
The concurrent presence of both thrombin and fibrin(ogen) in areas of disturbed flow implied that MacAIR cells might be involved in preventing fibrin formation; therefore, we explored fibrin(ogen) accumulation in the Cx3cr1CreERT2;Csf1rlslDTR model over time (Fig. 6a). Evaluation of fibrin(ogen) levels at 7 and 14 d after continued depletion showed progressive and significant accumulation in the lesser curvature (Fig. 6b–d and Extended Data Fig. 8d). Additionally, fibrin fibrils, which were not detected in any control mice in our experiments, were clearly visible in the aortae of mice depleted of macrophages for 14 d (Fig. 6c). The data imply that MacAIR cells promote clearance of fibrin(ogen) and/or prevention of fibrin formation. Further, en face SEM images of aortae (Fig. 6e and Extended Data Fig. 8e) revealed microthrombi and polymerized fibrin decorating the lesser curvature of Cx3cr1CreERT2;Csf1RrlslDTR mice but not present in littermate controls. Microthrombi were also noted by confocal microscopy along with rupture of the endothelial lining (Fig. 6f,g). As a direct readout of disseminated microthrombi, we evaluated d-dimer levels and found markedly elevated levels in macrophage-depleted mice compared to undetectable levels in control mice (Fig. 6h), further supporting hemostatic imbalance. Histological examination of tissues from MacAIR-depleted mice revealed hemorrhagic foci in multiple tissues, including the kidney, liver and lung (Fig. 6i). Images of kidneys from Cx3cr1CreERT2;Csf1rlslDTR mice that had to be euthanized due to health decline exhibited abundant fibrin(ogen) throughout the tissue (Fig. 6j), consistent with vascular rupture. Based on the data presented, it is likely that dislodged microthrombi traveling through the circulatory system were responsible for occluding smaller-diameter vessels, leading to hemorrhagic foci and organ damage. Additional support for the requirement of MacAIR cells in clearing fibrin(ogen) deposits also emerged from evaluation of Cd11c−/− mice. The absence of CD11c resulted in a significant reduction in MacAIR numbers, indicating that that CD11c was necessary, albeit not fully sufficient, for anchorage of MacAIR cells to the intimal niche, as we could still detect some macrophages (approximately 45% in comparison to controls) in the tunica intima of this mouse. Importantly, this mouse model also exhibited an impressive accumulation of fibrin(ogen) (Extended Data Fig. 8f).

a, Experimental design for long-term MacAIR depletion. Cx3cr1CreERT2;Csf1rlslDTR mice and littermate controls were injected every other day, alternating between tamoxifen and DTx. b, Accumulation of fibrin(ogen) (green) in the lesser curvature 14 d after MacAIR depletion in Cx3cr1CreERT2;Csf1rlslDTR mice compared to in control mice (scale bars, 500 μm (left) and 150 μm (right); n = 4 (control), n = 11 (Cx3cr1CreERT2;Csf1rlslDTR) mice per group. c, Fibrin fibrils (green) after MacAIR depletion (14 d) (yellow dotted ovals). VE-Cad, white; CD45, pink (scale bar, 10 μm; n = 4 (control), n = 11 (Cx3cr1CreERT2;Csf1rlslDTR) mice per group). Fb(g), fibrin(ogen). d, Mean fluorescence intensity (MFI) of fibrin(ogen) in the lesser curvature of Cx3cr1CreERT2;Csf1rlslDTR and control mice 14 d after depletion (n = 4 mice per group, Mann–Whitney t-test, mean ± s.d., P = 0.0286, two tailed, *P ≤ 0.05). AU, arbitrary units. e, SEM showing microthrombi in the lesser curvature of control and Cx3cr1CreERT2;Csf1rlslDTR mice 14 d after depletion. Scale bars, 20 μm and 5 μm; n = 3 mice per group. f, Microthrombi (green) and intima ruptures (yellow arrowheads) in a Cx3cr1CreERT2;Csf1rlslDTR mouse that was euthanized due to declined health 7 d after depletion. Scale bar, 20 μm. g, Intima ruptures with high fibrinogen (yellow arrowheads) in Cx3cr1CreERT2;Csf1rlslDTR mice 14 d after depletion (fibrinogen, green; VE-Cad, red). Scale bar, 20 μm; n = 4 mice. h, d-dimer plasma levels in Cx3cr1CreERT2;Csf1rlslDTR and control mice 14 d after depletion (n = 6 (Cx3cr1CreERT2;Csf1rlslDTR), n = 13 (control) mice; Mann–Whitney t-test, mean ± s.e.m., P = 0.0024, two tailed, **P ≤ 0.01). i,j, Kidneys, lung and liver from a Cx3cr1CreERT2;Csf1rlslDTR mouse with 7 or 14 d of macrophage depletion. i, Multiple hemorrhagic foci (blue arrowheads). j, Fibrin(ogen) (green) on kidney sections of control and Cx3cr1CreERT2;Csf1rlslDTR mice. aSMA, alpha smooth muscle actin. Scale bar, 50 μm; n = 4 (control), n = 11 (Cx3cr1CreERT2;Csf1rlslDTR) mice per group. k, Flow cytometry to assess the percentage of CX3CR1+CSF1R+CD45+ cells in bone marrow (BM), lung, liver, kidney and peripheral blood in control and Cx3cr1CreERT2;Csf1RlslDTR mice 1 d after depletion (n = 5 mice per group (n = 3 mice, peripheral blood), Mann–Whitney t-test, mean ± s.e.m.; P = 0.007937 (bone marrow), P = 0.0065 (peripheral blood); two tailed, **P ≤ 0.01). Q, quartile. l, Transcript levels of plasminogen receptors (Eno1 and Plgrkt) in adventitial macrophages (adv Mφ) and MacAIR cells from scRNA-seq (Extended Data Fig. 2). m, Left, experimental strategy for intravenous injection of FITC-conjugated albumin or plasminogen. Right, distribution of injected FITC-conjugated albumin (control, top) or plasminogen (bottom) in relation to MacAIR cells (yellow arrowheads). CD45, white; PECAM1, red. Scale bar, 5 μm; n = 3 per group. The illustration was created using https://biorender.com/.
An alternative explanation for fibrin(ogen) accumulation and thrombosis upon MacAIR depletion could relate to breach of barrier integrity in areas of oscillatory flow, implicating MacAIR cells in the maintenance of endothelial junctional integrity in these areas. We tested this possibility by examining fluorescent microsphere (40 nm) deposition in the tunica intima of littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice that both received tamoxifen and DTx injections (1 d of MacAIR depletion) (Extended Data Fig. 8g). While the positive control (buffered EDTA intracardiac injection for 5 min) resulted in robust deposition of fluorescent beads in between cells, no accumulation of fluorescent beads was found in any of the other groups (Extended Data Fig. 8g). Thus, MacAIR cells do not play a role in maintaining barrier integrity; instead, they appear to be necessary to clear fibrin(ogen) deposits and mitigate PAR1–thrombin signaling in regions of disturbed flow.
The drastic phenotype observed from depletion of MacAIR cells, a relatively small population, prompted the question of whether the approach might affect a broader group of macrophages. Thus, we evaluated alterations in the macrophage populations of multiple organs by flow cytometry analysis (Fig. 6k). The findings revealed that, while bone marrow and peripheral blood were affected by the dose of tamoxifen and DTx used, none of the other CX3CR1+CSF1R+ macrophage populations in the evaluated organs were altered. The implication is that areas of oscillatory and disturbed flow in the arterial tree are sites of fibrin accumulation that absolutely depend on MacAIR cells for clearance with critical consequences. A few last important pieces of evidence to solidify this conclusion are still pending, including how MacAIR cells degrade fibrin(ogen).
Macrophages are known to express plasminogen receptors to degrade fibrin(ogen) extracellularly41. Importantly, MacAIR cells expressed plasminogen receptors (Fig. 6l) and bound to fluorescently conjugated plasminogen when injected intravenously (Fig. 6m). Thus, in addition to MMP12 and MMP13, MacAIR cells are capable of generating cell surface-associated plasmin, particularly with the aid of endothelial cells, which express high levels of tissue-type plasminogen activator (Extended Data Fig. 8h,i). We also confirmed that neither endothelial cells nor MacAIR cells expressed plasminogen (Extended Data Fig. 8j). These findings indicate that MacAIR cells are required to clear fibrin(ogen) deposits, prevent fibrin formation and maintain an anti-thrombotic state in areas of disturbed flow.
Additional support for the conclusion that MacAIR cells are responsible for clearing fibrin(ogen) deposits in regions of disturbed flow came from experiments in which MacAIR cells were allowed to return after a 2-week depletion. Much like resident macrophages in other organs27,28,42,43,44, we found that, upon removal of MacAIR cells and elimination of depletion pressure (tamoxifen and DTx), monocytes seeded areas of disturbed flow and reconstituted the MacAIR population (Fig. 7a–d). A gradual increase in macrophage number was noted 1 and 2 weeks after depletion (Fig. 7d). Importantly, the increase in MacAIR numbers was associated with a reduction of accumulated fibrin(ogen) (Fig. 7c,e). Moreover, circulating d-dimer levels (Fig. 7f) and endothelial cell size in the lesser curvature of the aorta (Fig. 7g) also returned to control levels, demonstrating full rescue of the phenotype. Overall, these findings further support the conclusion that MacAIR cells are required to clear fibrin(ogen) deposits in regions of disturbed flow.

a, Images of littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice 1 week after a single DTx injection. Stopping tamoxifen and DTx injections relieves the pressure of cell ablation and allows for the repopulation of intimal immune cells. Aortae were stained for MacAIR markers to validate the monocyte-derived population that repopulates the arch after the resident population is depleted (MacAIR identity, CD11c+MMP12+CD68+CD45+) (n = 3 mice per stain; scale bars, 15 μm (left) and 10 μm (right)). b, Schematic of macrophage rescue experiments. Littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice were both injected with tamoxifen and DTx for 2 weeks of continued macrophage depletion. After 2 weeks, we stopped administration of tamoxifen and DTx, allowing MacAIR cells to repopulate the aorta. Aortae were evaluated en face at 2 weeks of depletion (baseline) and after ceasing tamoxifen or DTx treatment (1 week after depletion and 2 weeks after depletion). c, En face images evaluating fibrin(ogen) (green) and MacAIR (red) accumulation in aortae of littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice at the given time points. Fibrin fibrils are indicated by yellow arrowheads. VE-Cad, white; 1 week after, 1 week after depletion; 2 weeks after, 2 weeks after depletion. Scale bars, 300 μm and 7 μm; n = 6 mice per group. d, Total number of intimal CD45+ cells in the lesser curvature of the aortic arch in littermate controls and Cx3cr1CreERT2;Csf1rlslDTR mice at 2 weeks of depletion, 1 week after depletion and 2 weeks after depletion (n = 5 (control, 1 week after), n = 6 (DTR, 2 weeks after), n = 7 mice per group; Mann–Whitney t-test, mean ± s.d., P = 0.00116 (2-week depletion), two tailed, **P ≤ 0.01). e, Mean fluorescence intensity of fibrin(ogen) in the lesser curvature of the aortic arch (n = 5, 6 (DTR, 2 weeks after), n = 10 (DTR, 1 week after) mice per group; Mann–Whitney t-test, mean ± s.e.m., P = 0.007937 (2-week depletion), two tailed, **P ≤ 0.01). f, d-dimer plasma levels (n = 5 (control, 2 weeks after), n = 7 (control, 2-week depletion and 1 week after), n = 8 (DTR, 1 week after), n = 9 (DTR, 2 weeks after), n = 10 (DTR, 1-week depletion) mice; Mann–Whitney t-test, mean ± s.d., P = 0.0034 (2-week depletion), two tailed, **P ≤ 0.01). g, Surface area measurements of endothelial cells in the lesser curvature of Cx3cr1CreERT2;Csf1rlslDTR mice and littermate controls (n = 4 mice per group, Mann–Whitney t-test, line at median; P < 0.0001 (exact, 2-week depletion), ****P ≤ 0.0001).
Discussion
Vascular endothelial and hematopoietic cells are well known to coordinate inflammatory responses. Here, we have expanded these functions to also include regulation of intravascular hemostasis. Indeed, our findings indicate that, while endothelial cells provide a non-thrombogenic surface, facilitating blood circulation in areas of disturbed flow, this function is challenged by the accumulation of fibrinogen and thrombin (aortic arch and branches with rapid flow). In these regions, the presence of a population of intima resident macrophages (MacAIR), summoned to areas of disturbed flow from birth, is critical to effectively clear fibrinogen and prevent intravascular clotting.
Macrophage association with the endothelium is not necessarily surprising, and, when seen in aortic tissue sections, the assumption is that this heterotypic interaction might be part of an inflammatory or pre-atherosclerotic lesion. However, the unusual feature that captured our attention was that seeding of this macrophage population occurred after birth; furthermore, these cells displayed a unique topology in relation to the endothelium. These elements indicated that the presence of these macrophages in the luminal aspect of the aorta was not part of an inflammatory response; instead, the process was a normal developmental program by which MacAIR cells become a constitutive component of the tunica intima in regions of disturbed flow. While here we addressed multiple points related to origin, lineage and self-renewal, questions such as why these cells are attracted to areas of disturbed and oscillatory flow and how and why do they migrate from the closing ductus arteriosus to these regions remain unanswered. A logical assumption is that disturbed flow alters the endothelium, creating unique niche conditions that attract monocytes to these sites and promote MacAIR differentiation into a stable population capable of self-renewal.
The ability of macrophages to cleave fibrinogen and participate in the remodeling of the provisional matrix formed during wound healing has been established long ago45. However, the repertoire of proteases is distinct in different macrophage subtypes; some proteases are expressed only upon induction and in situations of wound healing. We found that, unlike adventitial and other macrophage populations, MacAIR cells constitutively express MMP12 and MMP13, known to cleave fibrinogen46,47,48. In addition, MacAIR cells also bind to plasminogen, and this anchorage enables endothelial tissue plasminogen activator (tPA) to generate plasmin, which also degrades fibrinogen and fibrin. Together, these proteases enable intravascular macrophages to efficiently remove fibrinogen deposits and antagonize fibrin accumulation driven by procoagulant pathways and the presence of thrombin in areas of oscillatory and disturbed flow.
Supporting the biological role attributed to MacAIR cells, it is pertinent to remember that, while compatible with development and reproduction, plasminogen deficiency results in a severe thrombotic phenotype in both normal and inflamed tissues of adult animals39,49,50. Homozygous plasminogen-deficient mice display spontaneous thrombotic lesions in multiple organs and a median survival of 176 d, with about 40% of mice succumbing to death. Additionally, these death phenotypes are effectively reversed by the simultaneous imposition of fibrinogen deficiency39. These findings underlie a critical constitutive function of the plasminogen-activation system for fibrin surveillance and clearance in non-pathological settings and resonate extremely well with the findings described here.
Given the association of MacAIR cells with areas of disturbed flow, also known to be pro-atherogenic sites, their potential contribution to atherosclerotic lesions is an important question. Specifically, one could predict that MacAIR cells are particularly poised to be the progenitors of foam cells. This question was the focus of a recently published elegant study15. The authors found that, while MacAIR cells can take up lipids, elimination of these cells only slightly delays but does not alter the burden of atherosclerotic lesions, nor does it change the accumulation of foam cells in lesions. Altogether, the conclusions of that study indicate that, while participating in the process, it is the influx of circulating monocytes into the neointima that is the main source of foam cells. In context, these data together with the fact that MacAIR cells embed themselves in the tunica intima immediately after birth concur with the notion that the presence of these macrophages is not a response to a pathological insult.
The present report adds to the long list of studies that have recently identified self-renewing tissue-resident macrophages in multiple organs21,22,28,51,52,53,54, now including the aortic endothelium in this list. These tissue-resident macrophages were found to seed multiple organs either during embryonic development or shortly thereafter and are derived from either yolk sac erythroid–myeloid or circulating myeloid progenitors27,28.
The findings presented here shift several paradigms. First, they challenge the concept that associations between the endothelium and leukocytes are always transient and triggered by acute inflammatory events. In fact, our results highlight a long-term partnership between endothelial cells and macrophages that is not dependent on immune responses; instead, it is triggered by the drastic hemodynamic changes associated with birth. Second, they change the view that a homotypic endothelial layer forms the luminal side of vessels, which now needs to be amended to include macrophages in areas of disturbed flow and aged arteries. Lastly, our findings indicate that intravascular clotting in arteries is constantly antagonized in regions of disturbed flow and the ‘aged’ endothelium. While the non-thrombogenic function of vessels was attributed exclusively to the endothelium, the data presented here provide clear evidence that, in some regions of the vascular tree, this can only be accomplished with the aid of macrophages.
Methods
Mice
All animal procedures were approved and performed in accordance with the University of California, Los Angeles (UCLA) and Northwestern University (NU) Institutional Animal Care and Use Committee. All other mouse information can be found in Supplementary Table 9. Mouse strains were maintained on a C57BL/6J background, with the exception of Csf1rmerCremer mice, which had a mixed background (FVB:C57BL/6). Mice were genotyped by Transnetyx. Male and female mice were used in approximately equal numbers for all experiments except for scRNA-seq experiments in which only male C57BL/6 mice were used to minimize potential sex and strain differences at the transcriptional level for the arch (8 weeks) and ‘aged descending (78 weeks) A,B’ datasets. However, validation of transcripts was performed in male and female mice. Unless specified, all adult mice used were 8–10 weeks of age.
Aorta en face collection
Adult mice were injected intraperitoneally (i.p.) with 10 mg methacholine to promote smooth muscle cell relaxation and facilitate en face imaging. Immediately after injection, mice were killed and perfused with 10 ml 2% paraformaldehyde (PFA) through the left ventricle (for embryos and neonates, 0.5–3 ml 2% PFA was used). Following perfusion, aortae were removed, and the adventitia was dissected under a microscope. Aortae were opened longitudinally, transferred to a 35-mm silicon-coated dish filled with 2% PFA and pinned to lay flat, exposing the endothelium. Fixation proceeded for 1 additional hour at 4 °C followed by washes with PBS.
Aortic en face immunostaining
For immunostaining, tissue was washed three times with 1× Hank’s balanced salt solution (HBSS) and incubated in blocking–permeabilization buffer (0.3% Triton X-100, 0.5% Tween-20, 3% normal donkey serum) for 1 h. The primary antibody cocktail was prepared in blocking–permeabilization buffer and incubated overnight at 4 °C (the endothelial marker ERG, VE-Cad or PECAM1 were always used in conjunction with other markers to label the endothelium). Aortae were washed three times with 1× HBSS and incubated with secondary antibodies for 1 h, washed with 1× HBSS and mounted on glass slides with ProLong Gold (Thermo, P36930). Antibodies and dilutions used are listed in Supplementary Table 9. Aortae were imaged using either an LSM880 confocal microscope (Zeiss) or an A1R HD25 confocal microscope (Nikon). Z-stack and tile scan features were used to image the large, wavy surfaces of the aortae. Resulting tiles were then stitched into a single large image (ZEN 2.0 Black software, Zeiss or NIS-Elements, Nikon), which enabled visualization of the large aortic arch at high resolution. Imaris software (Imaris 9.5.1 and 9.7.0, Bitplane) was used to visualize images in 3D. For a list of software used for analysis, see Supplementary Table 9. Additionally, Denoise.AI (Nikon) was employed to remove Poisson shot noise. Images were acquired using either ×20, ×63 or ×100 objectives.
In vivo labeling
Mice were injected via the tail vein with either rat anti-mouse CD45 antibody or non-blocking PECAM1-specific (390)–daylight 650 antibody diluted in sterile PBS to label the lumen-facing surfaces of MacAIR cells. Mice were killed 15 min after injection, and aortae were fixed, collected and stained with additional antibodies. For in vivo labeling of CD45, the following antibodies were additionally used: mouse anti-mouse CD45–biotin and streptavidin–A647, donkey anti-rat–A488 and anti-PECAM1 (2H8)–A568. For in vivo labeling of mice injected with PECAM1-specific (390) antibody, the following antibodies were used: anti-ERG and donkey anti-rabbit–A568, and anti-rat CD45 and donkey anti-rat–A488 (Supplementary Table 9).
Immunostaining and imaging of sections
PFA-fixed, paraffin-embedded specimens from kidneys were sectioned at 4 µm. Antigen retrieval was performed using 1× citrate buffer, and then samples were incubated in blocking–permeabilization buffer for 1 h. Sections were then incubated with primary antibodies (Supplementary Table 9) overnight at 4 °C. The following day, samples were incubated with species-specific secondary antibodies for 1 h before mounting in ProLong Gold. Samples were evaluated using an A1R HD25 confocal microscope (Nikon).
Bright-field imaging of aortae
Aortae from mice at E18.5, P1, P3, 1 week, 3 weeks and 10 weeks were dissected, and the adventitia was removed under a dissecting microscope. Aortae were pinned onto a 35-mm silicon-coated dish filled with PBS. Aortae were imaged using an Echo Revolve microscope (RVL-100-G).
Single-cell RNA sequencing
Isolation of intima cells was performed as previously described20. In summary, mice were anesthetized and perfused with 10 ml Versene buffer through the left ventricle. Under a dissecting microscope, the adventitia was removed, and the aorta was cut open in Versene buffer, exposing the endothelium. After Versene washes, aortae were bathed in 1× trypsin and incubated twice for 5 min at 37 °C. The endothelium was then gently removed using a microscalpel (EMS, 72046-30) and repeat pipetting, with sc-HBSS (containing 0.04% BSA and 2% FBS to inactivate trypsin and actinomycin D at 1 µg ml−1 to block transcription) applied to obtain a single-cell suspension (Extended Data Fig. 2a). Cells were pelleted at 300g and then treated with 1× red blood cell (RBC) lysis buffer (eBioscience, 00-4333-57) for 1 min and washed twice with 0.04% BSA. To obtain enough cells, six male (78-week-old; ‘aged descending’) or eight male (8-week-old; arch) C57BL/6 mice were used per library.
To isolate adventitia cells into a single-cell suspension, mice were anesthetized and perfused with 10 ml DMEM. The adventitia was dissected from aortae and dissociated using the Miltenyi Adipose Tissue Dissociation kit (130-105-808). Cells were pelleted at 300g. Following the Miltenyi protocol, the single-cell suspension was additionally treated with 1× RBC lysis buffer (eBioscience, 00-4333-57) and 1 U DNase. The final cell suspension was washed multiple times and resuspended in 0.04% BSA.
scRNA-seq libraries were generated using the 10x Genomics Chromium Single Cell 3′ Library & Gel Bead kit version 2. Cells were loaded accordingly following the 10x Genomics protocol with an estimated targeted cell recovery of 5,000 cells. Sequencing was performed on the Illumina HiSeq 4000 (paired end, 100 bp per read, 8-week-old mice, arch and ‘aged descending A,B’). The digital expression matrix was generated by demultiplexing, processing barcodes and counting gene unique molecular indices using the Cellranger count pipeline (version 4.0.0, 10x Genomics). Multiple samples were merged using the Cellranger aggr pipeline. To identify different cell types and find signature genes for each cell type, the R package Seurat (version 3.1.2) was used. Cells that expressed <100 genes or <500 transcripts were filtered out. We ran the DoubletFinder algorithm and set the doublet rate at 2% as recommended by the vendor (10x Genomics). The algorithm predicted that only 0.01% of doublets were captured. Variable genes were selected using the FindVariableGenes function for further analysis. Data were normalized using the NormalizeData function with a scale factor of 10,000. Genes were then scaled and centered using the ScaleData function. Principal-component analysis and t-SNE were used to reduce the dimensionality of the data. Cluster marker genes were found using the FindAllMarkers function. Cell types were annotated based on cluster marker genes. Heatmaps, violin plots and gene expression plots were generated by DoHeatmap, VlnPlot and featurePlot functions, respectively.
Flow cytometry of peripheral blood
To measure reporter labeling (lineage tracing) and donor chimerism (parabiotic mice) of circulating cells, blood was collected by retro-orbital bleeding into tubes containing FACS Buffer at 4 °C. Blood cells were pelleted at 300g and treated with 1× RBC lysis buffer. Additionally, cells were stained on ice with anti-CD45–APC-Cy7 (BD, 557659) and then analyzed on a BD Fortessa machine.
Flow cytometry of organs
One day after DTx injection (MacAIR depletion), littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice were euthanized and perfused with 10 ml Versene to remove blood and then perfused with 10 ml DMEM. Lung, liver, kidney and leg bones (femur and tibia) were dissected and washed with PBS. For bone marrow collection, muscle was removed, and bones were cleaned, crushed in FACS Buffer using a motor pestle to release bone marrow cells and treated with 1× RBC lysis buffer. For generation of single-cell suspensions from lung, liver and kidney, tissues were digested using Miltenyi’s Lung Dissociation kit (130-095-927), Liver Dissociation kit (130-106-807) and multidissociation kit 2 (130-110-203), following the vendor’s protocols. Cells were pelleted and treated with 1× RBC lysis buffer. Additionally, bone marrow, lung, liver and kidney cells were stained on ice with anti-CD45–APC-Cy7 (BD, 557659), anti-CX3CR1–APC (BioLegend, 149008), anti-CSF1R–BV421 (BioLegend, 135513) and anti-TER119–PE-Cy7 (eBioscience, 25-5921-82) antibodies and then analyzed on a BD Fortessa machine.
Lineage tracing
Homozygous lox-stop-lox-tdTomato reporter mice were crossed with homozygous Cx3cr1CreERT2 mice. Tamoxifen induction of Cre activity in the resulting F1 compound heterozygotes was initiated by dissolving tamoxifen (free base, MP Biomedicals, 0215673891) in sunflower seed oil (Sigma, S5007) and administering 0.01 mg tamoxifen via oral gavage to neonates at P1, P3 and P5. The penetration of intimal tdTomato+CD45+ cells in the aorta at P7 ranged from 40% to 62%, determined by en face confocal imaging (Fig. 3f); thus, analysis of 4-week-old adult mice needed to be normalized to data from littermates at P7. For baseline controls, half of the litter was killed, and blood as well as aortae were collected to determine reporter labeling efficiency. At 4 weeks, the remaining littermates were killed, and blood as well as aortae were collected to determine reporter expression.
Reporter expression in adult Cx3cr1CreERT2;R26lox-stop-lox-tdTomato mice was induced after i.p. injection of 1 mg tamoxifen three times, every other day. The penetration of tdTomato-positive intimal CD45+ cells in the aorta ranged from 97% to 100%. Aortae and blood were collected at 1 week after tamoxifen treatment as a baseline control for normalization purposes. Littermates were killed 10 weeks after injection to assess retention of reporter expression in adulthood.
For clonal tracing, Cx3cr1CreERT2;R26Rainbow (ref. 55) neonates were administered 0.01 mg tamoxifen via oral gavage at P1, P3 and P5 to yield permanent expression of three mutually exclusive fluorescent proteins: Cerulean, mOrange and mCherry. At 8 weeks, mice were killed, and reporter labeling was assessed. Adult Cx3cr1CreERT2;R26Rainbow mice were injected with one dose of 0.02 mg tamoxifen, and aortae were assessed 9 months later (at 11 months of age).
In vivo macrophage depletion
To deplete macrophages, adult (8–10-week-old) Cx3cr1CreERT2;Csf1rlslDTR mice were first injected with tamoxifen i.p. to promote Cre-dependent induction of the DTR. The following day, mice were injected with 200 ng DTx (Sigma, D0564) in sterile 1× PBS to induce apoptosis of DTR-expressing cells. This resulted in cell death of this population in as quickly as 24 h. However, as intimal, aortic myeloid cells are replenished quickly, evaluation of longer time points required continuous injection with tamoxifen followed by DTx every 3 d until the end of the experiment.
EdU-incorporation assay
Mice were injected i.p. with EdU (Fisher Scientific, A10044 at 10 mM). Two hours after injection, mice were terminally anesthetized and perfused with 2% PFA. The aorta was removed, and the adventitia was dissected. Aortae were longitudinally cut to exposed the endothelium. Following fixation, EdU was revealed using A647 following the manufacture’s protocol (Invitrogen, C10640). Additionally, aortae were stained for the nuclear endothelial marker ERG and the pan-hematopoietic marker CD45 and with DAPI before being imaged (Supplementary Table 9).
Scanning election microscopy
Mice were anesthetized and perfused with 10 ml 2% PFA through the left ventricle. Aortae were first immunostained en face following protocols listed above. High- and low-resolution images were then obtained by confocal microscopy. Aortae were then washed with 1× HBSS, dehydrated with increasing concentrations of ethanol and subjected to critical point drying followed by gold and/or palladium coating using a sputter coater. High- and low-resolution SEM images were also taken to use landmarks and find macrophages identified with the confocal images. Confocal and SEM images were then overlaid together using Adobe Photoshop (22.5.0).
To image microclots in aortae from macrophage-depleted (Cx3cr1CreERT2;Csf1rlslDTR) and control mice, mice were perfused with 4% glutaraldehyde through the left ventricle. Aortae were dissected and treated for an additional hour with 4% glutaraldehyde at room temperature and then washed several times with 1× PBS. Aortae were incubated in 1% osmium tetroxide for 1 h, dehydrated with a series of ethanol, dried to the critical point, mounted on pins and coated with 10-nm gold particles for SEM. Aortae were then imaged using a JEOL NeoScope microscope at 10 kV or 15 kV.
Dabigatran treatment
Mice were injected twice daily (morning and night) i.p. with 300 µg dabigatran (BIBR 953, Selleck, S2196) per injection for 5 d. Dosage was determined by the clotting test. After 5 d of treatment, mice were sacrificed, and aortae were analyzed.
In vivo siFibrinogen knockdown
Mice were injected with either siFibrinogen or an siRNA targeting luciferase (siLuciferase, control) at 1 mg siRNA per kg body weight via tail vein injection. siFibrinogen and siLuciferase were each encapsulated in lipid nanoparticles, composed of an ionizable lipid (DLin-MC3-DMA), phosphatidylcholine, cholesterol and a polyethylene glycol lipid, using methods previously described56. To determine knockdown efficiency, we quantified fibrinogen protein levels in plasma 7 d after injection. For this, we collected peripheral blood in EDTA-coated tubes (BD, 365974) and isolated plasma by centrifugation. Plasma was diluted (1:50), and fibrinogen levels were measured using a Mouse Fibrinogen ELISA kit (Abcam, ab213478).
Intravenous administration of fbg–a488 or plasminogen–FITC
Mice were injected intravenously with 400 µg fbg–a488 (Thermo, F13191), plasminogen–FITC (Abcam, ab92770) or albumin–FITC (control, Thermo, A23015). Mice injected with fbg–a488 were killed 3 h after injection, and mice injected with plasminogen–FITC were harvested 4 h after injection to assess local accumulation relative to the MacAIR cells.
d-dimer measurements
Mice were anesthetized, and blood was collected via right ventricle puncture with a 25G syringe. For d-dimer measurements, blood was collected in citrate buffer with a final concentration of 3.2% citrate (for d-dimer). Blood was centrifuged, and citrate plasma was collected and shipped to IDEXX on dry ice for quantification.
Comparison to the Tabula Muris Atlas and the Chakarov et al. study
MacAIR cells were compared to monocytes or macrophages from the Tabula Muris Atlas25. Specifically, expression values for all monocytes and macrophages were extracted from the Tabula Muris Atlas and merged with expression values of MacAIR cells from our dataset. The Seurat pipeline described above was applied to cluster cells. As the datasets were generated by different laboratories, the RunHarmony function from the R package Harmony57 was applied to remove potential batch effects among different tissues. After clustering, MacAIR marker genes were used to calculate a module score, which was used to identify the cell population from the Tabula Muris Atlas that was similar to MacAIR cells. The module score was calculated based on the average expression of genes in the list, subtracted by the aggregated expression of randomly chosen control genes.
Our monocyte and macrophage scRNA-seq datasets were also overlapped with data from the two macrophage populations obtained by Chakarov et al.23. For this marker, genes extracted from the comparison between monocytes, adventitial macrophages and MacAIR cells in our study were compared to data from Lyve1loMHCIIhi and Lyve1hiMHCIIlo cells (MHCII complex contains multiple genes: M2-Aa, M2-Ab1, M2-Eb2, M2-Eb1) from the Chakarov et al. study. The Jaccard index between each pair for cell types from the two datasets was calculated and plotted in the heatmap.
Parabiosis
Mice of similar weight and sex were housed together for 2 weeks before surgery to assess compatibility. Surgeries were performed as described previously58. In short, mice were anesthetized, and matching skin incisions were made from the olecranon to the knee joint of each mouse, and the subcutaneous fascia was bluntly dissected to create ~0.5 cm of free skin. The right olecranon of one animal was attached to the left olecranon of the other by a single 3-0 nylon suture and tie. The partners’ knee joints were similarly connected. Dorsal and ventral skins were closed, approximated by staples, and the animals were warmed with heating pads and monitored until recovery. Parabiotic pairs were housed one pair per cage and given acidified water (pH 2.5). After 4 weeks of anastomosis, blood samples from each animal in a parabiont pair were analyzed using flow cytometry. Animal pairs with <30% blood chimerism were excluded from our studies.
Annexin staining in aortae
Adult mice were killed and perfused with 3 ml 5% Annexin–A488 conjugate (Invitrogen, A13201) through the left ventricle. Five minutes later, the aorta was dissected and transferred to a 35-mm silicon-coated dish filled with 20% Annexin–A488. Annexin–A488 (20%) was also used to flush the Annexin inside the intact vessel and then incubated at 37 °C for 10 min. Following incubation, tissue was fixed in 2% PFA for 1 additional hour at room temperature, followed by multiple washes with PBS. Aortae were then opened and pinned to lay flat, exposing the endothelium and immunostained as described previously with anti-ERG and anti-CD45 antibodies and DAPI (Supplementary Table 9).
In vitro thrombin and MMP13 treatment and thrombin treatment with shear stress
HAECs were isolated as described from aortic trimmings of donor hearts at UCLA59. Cells were de-identified and exempt from consideration as human individuals by institutional regulatory boards at the UCLA. Additional HAECs were purchased from Lonza (CC-2535). HAECs were grown on gelatin-coated, tissue culture-treated six-well plates (Falcon, 353046) to confluency in complete MCDB-131 medium (Vec Technologies, MCDB131-WOFBS) with 10% FBS (Omega Scientific, FB-11). Cells were cultured in a humidified incubator at 37 °C with 5% CO2 and used between passages 4 and 9. Once confluent, cells were washed with PBS to remove serum and incubated with serum-free MCDB-131 medium. The next day, cells were treated with vehicle, MMP13 (1 µg, Abcam, ab227435) or thrombin (0.625 U ml−1, Sigma, 10602400001) for 5 min. After treatment, cells were washed with phosphate buffered saline (PBS) containing Na3OV4, and lysates were collected using mRIPA (50 mM Tris, 0.1% SDS, 0.5% sodium deoxycholate, 1% Triton X-100, 150 mM NaCl, 1 mM Na3VO4, protease inhibitor cocktail). Lysates were denatured in Laemmli buffer with 2-mercaptoethanol at 95 °C for 10 min. Proteins were separated with an SDS–PAGE gradient (4–20%, Bio-Rad, 4561094DC) gel, transferred onto nitrocellulose membranes (Bio-Rad, 1704271) and incubated overnight at 4 °C with primary antibodies. Primary antibodies were anti-pMLC2 (3671S), anti-MLC2 (CST, 3672S), anti-PAR1 (Thermo, PA5-19102) and anti-γ-tubulin (Abcam, ab11321). HRP-conjugated secondary antibodies were applied in a species-dependent manner for 1 h. Immunocomplex detection was performed with the enhanced chemiluminescence SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Fisher Scientific, 34095) using the ChemiDoc XRS+ Molecular Imager (Bio-Rad Laboratories). Quantification of bands was performed using Image Lab Software (Bio-Rad Laboratories).
For thrombin and shear stress experiments, HAECs were grown on gelatin-coated, glass-bottom, six-well plates (Cellvis, P06-1.5H-N) to confluency in complete MCDB-131 medium supplemented with 10% FBS. Once confluent, cells were washed with PBS to remove serum, and medium containing MCDB-131 and 4% dextran (Sigma-Aldrich, 31392) was added to confluent HAECs. Vehicle (PBS) or thrombin (Sigma, 10602400001) at a final concentration of 0.625 U ml−1 was then applied. Monolayers were subjected to unidirectional constant laminar flow for 48 h at 130 r.p.m. in a horizontal circular orbit (Benchmark, BT302). Static monolayers used the same dextran-containing medium and were cultured alongside flow-treated monolayers. After flow treatment (48 h), cells were fixed in 2% PFA for 15 min at room temperature and then washed with PBS.
For immunostaining, cells were incubated in blocking–permeabilization buffer (0.3% Triton X-100, 0.5% Tween-20, 3% normal donkey serum) for 1 h at room temperature. The primary antibody cocktail was prepared in blocking–permeabilization buffer and incubated with samples overnight at 4 °C. The following day, aortae were washed three times with 1× PBS and incubated with secondary antibodies for 1 h at room temperature.
Microsphere permeability
One day after DTx injection (MacAIR depletion), littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice were injected in the left ventricle with 200 µl 40-nm microspheres (1:10, Thermo, F8795). Beads circulated for 5 min, and then mice were euthanized and perfused with 10 ml PBS, followed by 10 ml 2% PFA. Aortae were collected, and whole-mount staining was performed as described above. For the positive control, C57BL/6 mice were injected with 50 mM EDTA buffer solution in the left ventricle, which was allowed to circulate for 5 min to challenge endothelial junctions. Next, these positive control mice were injected with microspheres and harvested as described.
Tail bleeding
One day after DTx injection (MacAIR depletion), littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice with same age and weight were anesthetized with ketamine–xylazine at a dose of 0.1 ml per 20 g. Using scalpel blade #11, tails of mice were resected exactly 3 mm distal from the tail end. Tails were quickly placed in tail-bleeding buffer (10 mM Tris-HCl, 2 mM CaCl2, warmed to 37 °C), and a stopwatch was started. The stopwatch was stopped after the blood stream halted.
Ex vivo nitric oxide measurements
A 5 mM stock solution of 4-amino-5-methylamino-2′,7′-difluorofluorescein (DAF-FM) diacetate (Invitrogen, D3844) was diluted to 10 µM in medium without phenol red (EBM, Lonza, CC-3129). One day after DTx injection (MacAIR depletion), littermate control and Cx3cr1CreERT2;Csf1rlslDTR mice were euthanized and perfused with 10 ml PBS followed by 3 ml 10 µM DAF-FM diacetate. Aortae were then quickly and gently collected in 10 µM DAF-FM. Intact aortae were then incubated in DAF-FM for 10 min at 37 °C. After 10 min, DAF-FM was replaced with fresh DAF-FM, and the sample was incubated for an additional 15 min. Aortae were then quickly filleted open and mounted with PBS. Confocal imaging occurred immediately after to evaluate nitric oxide levels.
Quantification and statistical analysis
Treatments were randomized; investigators were blinded to allocation for outcome assessment. No samples or animals were excluded from this study. Sampling size for animal experiments was determined by power analysis with a type 1 error rate of 5% and a minimum detectable effect of 20%. For cell surface experiments, we used sampling similar to previous published reports. For experiments for which the outcome was immunofluorescence, figures show representative images; however, the number of independent times that the experiment was reproduced using biological replicates is provided in the legend.
Quantification of intimal immune cells in aortae was performed using the spots function in Imaris 9.5.1 or 9.7.0 (Bitplane) on maximum-intensity Z projections. Only cells with clearly distinguishable bodies and nuclei (DAPI, not shown) were quantified. No samples or animals were excluded from this study.
Cell surface area and elongation factors of endothelial cells in aortae of control and macrophage-depleted (Cx3cr1CreERT2;Csf1rlslDTR) mice was determined in several regions of 44.1 mm2 within the lesser curvature of each whole-mount, flat-mounted aorta. Measurements were performed with NIS-Elements, using a combination of manual and automated cell shape identification. VE-Cad or PECAM1 staining was used to define endothelial cell borders. For macrophage-depletion experiments, we used littermate controls that received both tamoxifen and DTx injections. For evaluation of fluorescence, mean fluoresce intensity was determined using the surface function in Imaris 9.5.1 or 9.7.0 on maximum-intensity Z projections. Subsequently, the mean for at least three independent samples and s.d. was determined. To assess whether two datasets were significantly different, we calculated P values with unpaired, nonparametric Student’s t-test followed by the Mann–Whitney test; P < 0.05 was considered significant (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001). Statistical analyses were performed with Prism 8 (GraphPad Software).
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All data generated or analyzed are included in the main article and associated files. scRNA-seq data were deposited in the GEO database under accession numbers GSE161787, GSE125691 (ref. 22) and GSE154817 (ref. 15). The Tabula Muris Atlas dataset is deposited at https://www.czbiohub.org/tabula-muris/25. Source data are provided with this paper.
References
Diacovo, T. G., Roth, S. J., Buccola, J. M., Bainton, D. F. & Springer, T. A. Neutrophil rolling, arrest, and transmigration across activated, surface-adherent platelets via sequential action of P-selectin and the β2-integrin CD11b/CD18. Blood 88, 146–157 (1996).
Ley, K., Laudanna, C., Cybulsky, M. I. & Nourshargh, S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat. Rev. Immunol. 7, 678–689 (2007).
Liew, P. X. & Kubes, P. The neutrophil’s role during health and disease. Physiol. Rev. 99, 1223–1248 (2019).
Mamdouh, Z., Chen, X., Pierini, L. M., Maxfield, F. R. & Muller, W. A. Targeted recycling of PECAM from endothelial surface-connected compartments during diapedesis. Nature 421, 748–753 (2003).
Gisterå, A. & Hansson, G. K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 13, 368–380 (2017).
Quintar, A. et al. Endothelial protective monocyte patrolling in large arteries intensified by western diet and atherosclerosis. Circ. Res. 120, 1789–1799 (2017).
Libby, P., Nahrendorf, M. & Swirski, F. K. Leukocytes link local and systemic inflammation in ischemic cardiovascular disease: an expanded “cardiovascular continuum”. J. Am. Coll. Cardiol. 67, 1091–1103 (2016).
Robbins, C. S. et al. Local proliferation dominates lesional macrophage accumulation in atherosclerosis. Nat. Med. 19, 1166–1172 (2013).
Ross, R. Rous-Whipple Award Lecture. Atherosclerosis: a defense mechanism gone awry. Am. J. Pathol. 143, 987–1002 (1993).
Auffray, C. et al. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 317, 666–670 (2007).
Carlin, L. M. et al. Nr4a1-dependent Ly6Clow monocytes monitor endothelial cells and orchestrate their disposal. Cell 153, 362–375 (2013).
Hanna, R. N. et al. Patrolling monocytes control tumor metastasis to the lung. Science 350, 985–990 (2015).
Choi, J.-H. et al. Identification of antigen-presenting dendritic cells in mouse aorta and cardiac valves. J. Exp. Med. 206, 497–505 (2009).
Jongstra-Bilen, J. et al. Low-grade chronic inflammation in regions of the normal mouse arterial intima predisposed to atherosclerosis. J. Exp. Med. 203, 2073–2083 (2006).
Williams, J. W. et al. Limited proliferation capacity of aortic intima resident macrophages requires monocyte recruitment for atherosclerotic plaque progression. Nat. Immunol. 21, 1194–1204 (2020).
Broadhouse, K. M. et al. Assessment of PDA shunt and systemic blood flow in newborns using cardiac MRI. NMR Biomed. 26, 1135–1141 (2013).
Colucci-Guyon, E. et al. Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell 79, 679–694 (1994).
Salvador, J. et al. Transcriptional evaluation of the ductus arteriosus at the single-cell level uncovers a requirement for vimentin for complete closure. Preprint at bioRxiv https://doi.org/10.1101/2021.10.30.466605 (2021).
Ma, F., Hernandez, G. E., Romay, M. & Iruela-Arispe, M. L. Single-cell RNA sequencing to study vascular diversity and function. Curr. Opin. Hematol. 28, 221–229 (2021).
McDonald, A. I. et al. Endothelial regeneration of large vessels is a biphasic process driven by local cells with distinct proliferative capacities. Cell Stem Cell 23, 210–225 (2018).
Ensan, S. et al. Self-renewing resident arterial macrophages arise from embryonic CX3CR1+ precursors and circulating monocytes immediately after birth. Nat. Immunol. 17, 159–168 (2016).
Chakarov, S. et al. Two distinct interstitial macrophage populations coexist across tissues in specific subtissular niches. Science 363, eaau0964 (2019).
Lim, H. Y. et al. Hyaluronan receptor LYVE-1-expressing macrophages maintain arterial tone through hyaluronan-mediated regulation of smooth muscle cell collagen. Immunity 49, 326–347 (2018).
Hernandez, G. E. & Iruela-Arispe, M. L. The many flavors of monocyte/macrophage–endothelial cell interactions. Curr. Opin. Hematol. 27, 181–189 (2020).
Tabula Muris Consortium et al. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 562, 367–372 (2018).
Satpathy, A. T. et al. Zbtb46 expression distinguishes classical dendritic cells and their committed progenitors from other immune lineages. J. Exp. Med. 209, 1135–1152 (2012).
Epelman, S., Lavine, K. J. & Randolph, G. J. Origin and functions of tissue macrophages. Immunity 41, 21–35 (2014).
Ginhoux, F. & Guilliams, M. Tissue-resident macrophage ontogeny and homeostasis. Immunity 44, 439–449 (2016).
Serbina, N. V. & Pamer, E. G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 7, 311–317 (2006).
Boyer, S. W., Schroeder, A. V., Smith-Berdan, S. & Forsberg, E. C. All hematopoietic cells develop from hematopoietic stem cells through Flk2/Flt3-positive progenitor cells. Cell Stem Cell 9, 64–73 (2011).
Epelman, S. et al. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity 40, 91–104 (2014).
Mehta, D. & Malik, A. B. Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 86, 279–367 (2006).
Rigor, R. R., Shen, Q., Pivetti, C. D., Wu, M. H. & Yuan, S. Y. Myosin light chain kinase signaling in endothelial barrier dysfunction. Med. Res. Rev. 33, 911–933 (2013).
Austin, K. M., Covic, L. & Kuliopulos, A. Matrix metalloproteases and PAR1 activation. Blood 121, 431–439 (2013).
Hou, H.-H. et al. MMP-12 activates protease-activated receptor-1, upregulates placenta growth factor, and leads to pulmonary emphysema. Am. J. Physiol. Lung Cell. Mol. Physiol. 315, L432–L442 (2018).
Jaffré, F., Friedman, A. E., Hu, Z., Mackman, N. & Blaxall, B. C. β-adrenergic receptor stimulation transactivates protease-activated receptor 1 via matrix metalloproteinase 13 in cardiac cells. Circulation 125, 2993–3003 (2012).
Willis Fox, O. & Preston, R. J. S. Molecular basis of protease-activated receptor 1 signaling diversity. J. Thromb. Haemost. 18, 6–16 (2020).
Kulkarni, J. A., Witzigmann, D., Chen, S., Cullis, P. R. & van der Meel, R. Lipid nanoparticle technology for clinical translation of siRNA therapeutics. Acc. Chem. Res. 52, 2435–2444 (2019).
Bugge, T. H. et al. Loss of fibrinogen rescues mice from the pleiotropic effects of plasminogen deficiency. Cell 87, 709–719 (1996).
Prasad, J. M. et al. Mice expressing a mutant form of fibrinogen that cannot support fibrin formation exhibit compromised antimicrobial host defense. Blood 126, 2047–2058 (2015).
Motley, M. P. et al. A CCR2 macrophage endocytic pathway mediates extravascular fibrin clearance in vivo. Blood 127, 1085–1096 (2016).
Bonnardel, J. et al. Stellate cells, hepatocytes, and endothelial cells imprint the Kupffer cell identity on monocytes colonizing the liver macrophage niche. Immunity 51, 638–654 (2019).
Sakai, M. et al. Liver-derived signals sequentially reprogram myeloid enhancers to initiate and maintain Kupffer cell identity. Immunity 51, 655–670 (2019).
Scott, C. L. et al. Bone marrow-derived monocytes give rise to self-renewing and fully differentiated Kupffer cells. Nat. Commun. 7, 10321 (2016).
Colvin, R. B. & Dvorak, H. F. Fibrinogen/fibrin on the surface of macrophages: detection, distribution, binding requirements, and possible role in macrophage adherence phenomena. J. Exp. Med. 142, 1377–1390 (1975).
Hiller, O., Lichte, A., Oberpichler, A., Kocourek, A. & Tschesche, H. Matrix metalloproteinases collagenase-2, macrophage elastase, collagenase-3, and membrane type 1-matrix metalloproteinase impair clotting by degradation of fibrinogen and factor XII. J. Biol. Chem. 275, 33008–33013 (2000).
Motterle, A. et al. Influence of matrix metalloproteinase-12 on fibrinogen level. Atherosclerosis 220, 351–354 (2012).
Tardif, G., Reboul, P., Pelletier, J.-P. & Martel-Pelletier, J. Ten years in the life of an enzyme: the story of the human MMP-13 (collagenase-3). Mod. Rheumatol. 14, 197–204 (2004).
Bugge, T. H., Flick, M. J., Daugherty, C. C. & Degen, J. L. Plasminogen deficiency causes severe thrombosis but is compatible with development and reproduction. Genes Dev. 9, 794–807 (1995).
Goguen, J. D., Bugge, T. & Degen, J. L. Role of the pleiotropic effects of plasminogen deficiency in infection experiments with plasminogen-deficient mice. Methods 21, 179–183 (2000).
Gomez Perdiguero, E. et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 518, 547–551 (2015).
Hoeffel, G. et al. C-Myb+ erythro-myeloid progenitor-derived fetal monocytes give rise to adult tissue-resident macrophages. Immunity 42, 665–678 (2015).
Schulz, C. et al. A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336, 86–90 (2012).
Yona, S. et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 38, 79–91 (2013).
Rinkevich, Y., Lindau, P., Ueno, H., Longaker, M. T. & Weissman, I. L. Germ-layer and lineage-restricted stem/progenitors regenerate the mouse digit tip. Nature 476, 409–413 (2011).
Strilchuk, A. W. et al. Sustained depletion of FXIII-A by inducing acquired FXIII-B deficiency. Blood 136, 2946–2954 (2020).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289–1296 (2019).
Ali, S. R. et al. Developmental heterogeneity of cardiac fibroblasts does not predict pathological proliferation and activation. Circ. Res. 115, 625–635 (2014).
Navab, M. et al. Monocyte migration into the subendothelial space of a coculture of adult human aortic endothelial and smooth muscle cells. J. Clin. Invest. 82, 1853–1863 (1988).
Acknowledgements
We are grateful to G. Randolph and J. Williams for extensive discussions about intima resident macrophages and for the generous transfer of the Cx3cr1CreERT;Csf1rlslDTR transgenic mouse. We are also thankful to W. Mueller for anti-PECAM1 antibodies. We are thankful to K. Ridge for the generous transfer of Vim−/− mice. We also thank the Broad Stem Cell Research Center and the Jonsson Comprehensive Center at the UCLA for sequencing scRNA-seq libraries; the Broad Stem Cell Research Center flow core at the UCLA and the Robert H. Lurie Comprehensive Cancer Center flow core at NU, Feinberg School of Medicine; the Mouse Histology and Phenotyping core at NU; and the Center for Advanced Microscopy at NU. Special thanks to M. Steel and S. Mirkov for managing animal colonies and for assistance with husbandry and animal experiments. Additionally, we thank S. Buchanan for help with cell size quantification. We also thank F.V. Korobova from the Center for Advanced Microscopy at NU for support with SEM. We thank members of the Arispe laboratory and the Feinberg Cardiovascular and Renal Research Institute at NU for extensive discussions. Illustrations were created by C. Bauista or with https://biorender.com/. This work was supported by R35HL140014 to M.L.I.-A. and a Howard Hughes Medical Institute Gilliam Fellowship (GT11560) to G.E.H. J.S. is supported by the NU Molecular and Translational Cardiovascular Training Program (T32HL134633, SP0040691). D.A.L. is supported by the NIH–NICHD training grant T32HD007491.
Author information
Authors and Affiliations
Contributions
G.E.H. performed and designed experiments and wrote and edited the manuscript; G.M., N.B.F., J.S. and D.A.L. performed experiments and quantification; L.J.J., J.L. and C.J.K. provided siRNA–nanoparticles; P.Z. performed parabiosis experiments, supervised by R.A. R.A. provided transgenic mice and provided intellectual input and discussion; A.E.B. and M.J.F. provided key transgenic mice and provided intellectual input for experimental design and analysis. M.P. provided intellectual input for experiments using scRNA-seq data; F.M. performed bioinformatic analysis of scRNA-seq data; M.L.I.-A. conceived the study, designed experiments and wrote and edited the manuscript. All authors discussed the results and had the opportunity to comment on the manuscript.
Corresponding author
Ethics declarations
Competing interests
C.J.K. is a director and shareholder of NanoVation Therapeutics Inc., which is commercializing RNA-based therapies. C.J.K., L.J.J. and J.L. have filed intellectual property on RNA-based therapies, with the intention of commercializing these inventions. All other authors declare no competing interests.
Peer review
Peer review information
Nature Cardiovascular Research thanks Klaus Ley, John Hwa and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Intimal immune cell distribution in the aorta and vena cava of young and aged mice.
a, Accumulation of intimal immune cells, as per CD45 (green arrows) in adult (n = 498 mice) and old 78wk (n = 6 mice) aortic arch. VE-Cadherin (red). Scale bars, 500μm & 15μm (aI-II). b, High magnification of en face distribution of CD45 + (green) cells in the lesser curvature (LC) of adult and aged aortae. Scale bar, 20μm. c, Quantification of intimal CD45 + cells in LC of 8wk and 78wk old mice. (n = 4 mice per group, Mann-Whitney T-test, ± SD, p = 0.0286, two-tailed, *p ≤ 0.05). d, e Images of the vena cava of 8wk old mice. d, Low magnification of vena cava (VE-Cadherin in white). Scale bar, 500μm. e, No intimal CD11c + (red) CD45 + (green) cells were found in the vena cava, even in regions of disturbed blood flow (Scale bar, 10μm, n = 3 mice). f, Few intimal CD45 + cells (green) accumulate in carotid arteries (8wk), VE-Cadherin in red (Scale bar, 300μm, 50μm (fI-II), n = 8 mice). High magnification inserts on the right showing carotid branches (disturbed flow). g, Whole mount scans of the descending aortae (thoracic and abdominal) of 8wk and 78wk old C57BL/6 mice. Except for branches, few intimal CD45 + cells (green) were detected in 8wk old descending aortae, (gI) whereas 78wk old mice display large number of intimal CD45 + cells (gII). Scale bar, 500μm (8wk thoracic), 700μm (8wk abdominal), 1000μm (78 wk thoracic and abdominal), and 50μm (gI-II), n = 5 (8wk) and 6 (78wk) mice. h, Topographic map to guide the quantification of immune cells shown in ‘i’. i, Intimal CD45 + cells per aortic region at 8wk, 52wk, and 78wk old mice (n = 3 mice per timepoint, two-tailed T-test, ± SD, p = 0.0022 (2: 8wk vs 52wk), p = 0.0031 (2: 8wk vs 78wk), p = 0.0175 (2: 52wk vs 78wk), p = 0.0396 (3: 8wk vs 78wk), p = 0.0052 (5: 8wk vs 52wk), (*p ≤ 0.05, **p ≤ 0.01).
Extended Data Fig. 2 scRNA-seq uncovers the macrophage identity and unique transcriptional signature of the intimal immune cell population.
a, Schematic to illustrate the process for isolation of intimal, aortic cells for single-cell RNA sequencing. b-d, Violin plots representing the Quality Control (QC) metrics for each scRNAseq library: Arch and Aged Thoracic A and B. Note, Aged Thoracic A & B represent two independent libraries, not technical replicates, each composed of cells isolated from 6-8 aortae. Shown are: the number of genes per cell (nGene), the absolute number of observed transcripts per cell (nUMI, Unique Molecular Identifier), and the fraction of transcripts arising from the mitochondria genome (fration of mito). b, Aortic arch library (red); c, Aged thoracic library A (green); d, Aged thoracic library B (blue). e, tSNE plot of cells from three independent libraries: aortic arch in red (8 wk) and aged descending (descen.) A (in green) and B (in red) (78wk). 6-8 aortas were used per library. f, Ten distinct cell types were identified, and cellular identities were assigned to each cluster. g, tSNE analysis and heat map-style representation of non-hematopoietic vascular lineage markers (color key indicates expression level). h, Violin plots showing expression levels of the most prominent hematopoietic lineage markers shown as normalized gene expression per cell. i, Top differentially expressed genes expressed by monocytes, adventitia macrophages (Adv. Mϕ), aortic intima-resident macrophages (MacAIR) (color key indicates expression level). j, Violin plots showing expression levels of selected genes that distinguished monocytes (left column), Adv. Mϕ (center left column), and MacAIR (right columns).
Extended Data Fig. 3 scRNAseq on dissected adventitia.
a, Illustration displaying the different aortic layers and highlighting the tissue that was dissected and used to generate independent scRNAseq libraries for comparison: tunica adventitia vs tunica intima, (red text). b, tSNE projections of cells from three independent libraries: aged adventitia (78 wk) and aged thoracic A and B (78wk). c, Expression of CD14 to identify myeloid cells in aged thoracic and aged adventitia data sets (color key indicates expression level). d, tSNE plot displaying only the CD14 expressing cells from c. Pink dotted circle highlights a cluster of cells that are exclusively found in intimal-enriched libraries (Aged Thoracic A and B). e, tSNE plot identifying unique cell clusters presented in d. Note, cluster 7 (pink dotted cluster) is a clearly distinct population. f, Expression of Lyve1 was used to identify adventitia macrophages and Fcgr1 was used to identify macrophages. Note, Cluster 7 expresses Fcgr1 and also highly expresses Mmp12 and Mmp13 but it does not express Lyve1 (color key indicates expression level). g, Cellular identities assigned to each cluster identified based on transcriptional expression patterns.
Extended Data Fig. 4 MacAIR comparison to other published macrophage data sets.
a, Uniform manifold approximation and projection (UMAP) plot of monocytes and macrophages extracted from the Tabula muris atlas and merged with the aortic intima-macrophage (MacAIR) data set (‘Aorta’) from this work. b, UMAP plot identifying distinct cell clusters based on transcriptional signatures. c, MacAIR marker genes were used to calculate a module score, which was used to identify cells from the Tabula muris atlas that were similar to MacAIR. The MacAIR module score was applied and represented by heat map-style UMAP plot. d, Number (top graph) and percentage (bottom graph) of cells from each tissue that are in each cluster. e, UMAP heat map-style representation of MMP12 (MacAIR marker), Cx3cr1, and Csf1r (top row) with violin plot representation of each given gene below. f, Heat map comparing the top 50 MacAIR markers to all the clusters found in b. g, UMAP plotting only cluster 2 and showing tissue origin. h, UMAP heat map-style representation of top MacAIR markers: Mmp12, Mmp13, Cxcl16, Itgax (CD11c), i, Differential genes from the comparison among monocytes, adventitia macrophages, and MacAIR (our data – this study) were overlapped with genes extracted from the comparison between the Lyvel°wMHCIIhigh and LyvehighMHCIIl°w macrophages from the Chakarov et al. dataset. The jaccard index between each pair for cell types from the two datasets were calculated and plotted in the heatmap. j, Comparison of MacAIRs identified in the tunica intima-enriched young arch vs aged descending data sets.
Extended Data Fig. 5 Validation of MacAIR markers identified by scRNAseq.
a, b, Validation of unique MacAIR genes at P7 (a) and adult (b) aortae. MacAIR markers in red (CXCL16 MMP13 and CD11c) and CD45 in green. Scale bar, 5μm and 10μm, n = 3 (P7), 5 (adult) mice. c, Evaluation of the same markers as in panels a and b, but now in the adventitia. Note that no CD45 + cells in adult adventitia expressed the MacAIR markers CXCL16, MMP13, and CD11c. Scale bar, 10μm; n = 3 mice per marker. d-e, MacAIRs express Cx3cr1 at both the (d) transcript level and (e) protein level (in red) (Scale bar, 5μm, n = 3 mice). f, Cx3cr1-reporter (Cx3cr1creERT2;R26tdT°mat°) showing tdTomato reporter expression in intimal immune cells one-day post-tamoxifen (Scale bar, 100μm and 30μm). 98% of intimal CD45 + cells in the lesser curvature (LC) of adult Cx3cr1creERT2;R26tdT°mat° mice were tdTomato + ; represented on adjacent graph (n = 3 mice, ± SD, GC, greater curvature). g, Intimal tdTomato+ cells colocalize with MacAIR marker Cxcl16, post-tamoxifen injection in Cx3cr1creERT2;R26tdT°mat° aortae. Scale bar, 10μm, n = 3 mice. h, CD45 + cells found in P5 Cx3cr1creERT2;R26tdT°mat° aortae were also tdTomato + (yellow arrowheads) after tamoxifen at P1 and P3 (Scale bar, 50μm and 10μm, n = 3 mice). VE-Cadherin (white). (DA, Ductus arteriosus). i, Csf1rmerCremer;R26tdT°mat° macrophage fate-mapping. Intimal CD45 + cells in the lesser curvature (LC) labeled by tdTomato. Adult mice received a single injection of tamoxifen. ERG (white) (Scale bar, 20μm; n = 2 mice). j, Images co-staining for CD68 (white), CD45 (red) and VE-Cadherin (green). Scale bar, 30μm and 7μm, n = 3 mice. k, Expression of classical dendritic cell (DC) markers (Zbtb46, Cd8a, Ccr7) and macrophage (Mφ) markers (Fcgr1, Adgre1, Cd68) in adventitia macrophages (Adv. Mφ), intima-resident macrophages (MacAIRs), and monocytes from scRNA-seq data in Extended Data Fig. 2. l, 3D surface rendering of an intimal CD45 + cell (white) phagocytosing an annexin + (green) endothelial cell (ERG + , red) in the LC. Scale bar, 10μm.
Extended Data Fig. 6 Aortic intimal macrophages are a self-replicating cell population of the tunica intima residing in areas of disturbed flow.
a, Timeline of 5-ethynyl-2′-deoxyuridine (EdU) incorporation. 8wk and 78wk old mice were injected with EdU and harvested 2-hrs later. b, EdU + (white) and CD45 + (green) co-expressing cells (yellow arrowheads) (arch and descending=Desc.). Scale bar, 20μm, n = 3 (78wk), 8 (8wk) mice. c, Intimal EdU+ CD45 + cells found in LC at 8wk (n = 4mice, ± SD). d, Experimental design for lineage tracing. 8wk old Cx3cr1creERT2;R26tdTomato mice were pulse-labeled by Tam injection (inj.). Mice were harvested at 1wk-post-tam to establish reporter baseline. Evaluation was done at 10- and 20-wks post-tam. e, Images of CD45 (green) and tdTomato+ at 1-, 10-, and 20-wks post-tamoxifen. Endothelial cell (EC) labeled with VE-cadherin (1- and 10-wk) and ERG (20-wk). Scale bar, 80μm; n = 4 (20wk), 5 (1wk,10wk) mice. GC = greater curvature. f, Representative fluorescence activated cell sorting (FACS) plot of tdTomato + / CD45 in the peripheral blood of Cx3cr1creERT2;R26tdTomato mice weeks after tamoxifen injection. g, Percentage of peripheral blood positive for tdTomato during the time course of the study post-tamoxifen measured by flow cytometry (n = 3 (2wk, 3wk, 4wk,10wk), 5(20wk), and 6 (1wk) mice, ±SD). h, Percentage of intimal tdTomato+CD45 + cells found the aortic arch (LC) of Cx3cr1creERT2;R26tdTomato mice at 1-, 10-, and 20-wks post-tamoxifen injection (n = 4 (20wk), 5 (1wk, 10wk) mice, Mann-Whitney t-test, ± SD, p = 0.0159 (1wk vs 10wk), p = 0.0159 (1wk vs 20wk), two-tailed, *p ≤ 0.05). i, Experimental design and timeline indicating the possible outcomes of the parabiosis experiment. GFP + and wild-type (WT) mice were surgically paired together to share chimeric circulation for 5wks prior to analyses. j, Representative FACS plot and histogram showing blood chimerism after parabiosis. k, Quantification of intimal GFP + CD45 + cells found in the LC of the WT mice (n = 2 pairs of mice, ± SD). PB = peripheral blood; WT = wild-type. l, Clonal tracing model, design, and possible outcomes. 2month old Cx3cr1creERT2;R26Rainbow were administered a single dosage of tamoxifen. At 11months (9 months later), aortae were evaluated clonal expansion and retention of labeling of MacAIRs overtime. Images revealed clones of MacAIRs labeled and CD45 (white) 9months after tamoxifen labeling. Scale bar, 30μm; n = 4 mice. Cerulean (Cer), mOrange (mOr), mCherry (mCh). m, Lineage tracing (Flt3cre;R26mTmG) that labels cells derived from definitive hematopoiesis. Flt3-driven cre recombinase (cre) expression leads to an irreversible switch from Tomato (tom) to GFP expression in Flt3-expressing cells. As this is an irreversible switch, progeny will also be labeled GFP. n, GFP expression in CD45 + cells of P5 Flt3cre;R26mTmG aortae relative to circulating monocytes from the same mice (n = 4 mice, Mann-Whitney t-test, ± SD, p = 0.0286, *p ≤ 0.05). o, Images of P5 Flt3cre;R26mTmG aortae showing GFP expression (green) of intimal CD45 + (blue) cells exiting the ductus arteriosus (DA) (n = 4 mice).
Extended Data Fig. 7 Thrombin distribution and signaling in areas of disturbed flow.
a, VE-cadherin (white) of control and macrophage-depleted (Cx3cr1creERT2;CSF1RlslDTR) aortae (upper panel). Changes in cell area and cell number were quantified by filling in red the area limited by VE-Cadherin staining. Note changes in shape and in cell number upon deletion of MacAIRs Scale bar, 15μm, n = 3 (control) or 4 (Cx3cr1creERT2;CSF1RlslDTR) mice. b, Low (upper panels) and high (lower panels) magnifications of control and Cx3cr1creERT2;CSF1RlslDTR aortae staining for phosphorylation of myosin light chain 2 (pMLC2) (in red) and CD45 (green) one day post-diphtheria (DTx) toxin injection. Scale bar, 500μm (upper panels), 50μm (lower panels); n = 5 mice per group. c, Effect of thrombin on human aortic endothelial cells (HAECs) pMLC2 (red) cultured under laminar flow. VE-cadherin (white) provides information on cell shape. DAPI (blue). Scale bar, 10μm. d, F2r (PAR1), F2rl1 (PAR2), and F2rl3 (PAR4) expression in endothelial cells (EC) and in MacAIRs from the arch scRNAseq data sets (Extended Data Fig. 2). e, En face images of the lesser curvature (LC) and greater curvature (GC) staining for thrombin (white) in control and Cx3cr1creERT2;CSF1RlslDTR aortae one-day after DTx (Scale bar, 50μm). f, Time of hemostatic plug formation measured following transverse amputation of the tip of the tail (3 mm) to assess hemostasis function in control and Cx3cr1creERT2;CSF1RlslDTR mice one-day post-DTx injection (n = 11 mice per group). g, Ex vivo mean fluorescent intensity (MFI) measurements of nitric oxide in the lesser curvature (LC) and thoracic aorta from control and Cx3cr1creERT2;CSF1RlslDTR mice one-day post-DTx injection (n = 2 mice per group and 3-7 number of areas quantified per mouse; Mann-Whitney T-test, ± SD, p = 0.0005, two-tailed ****p ≤ 0.0001). DAF-FM Diacetate (4-Amino-5-Methylamino-2’,7’-Difluorofluorescein Diacetate) was used to detect and quantify NO. h, Plasma levels of prostacyclin in control and Cx3cr1creERT2;CSF1RlslDTR one-day post-DTx injection, measured by ELISA. No statistically significant changes noted (n = 4 mice per group, Mann-Whitney t-test, ± SD).
Extended Data Fig. 8 MacAIRs are necessary to prevent fibrin accumulation in areas of turbulent flow.
a, Images of the greater curvature after treatment with siFibrinogen or siLuciferase (control) for 7 days and then exposed to Fbg-a488 intravenously for 3 hrs. No fibrinogen binds to the greater curvature (unlike Fig. 5f). Scale bar, 7μm; n = 4 mice per group. b, Descending aortae (78wk old) stained for fibrin(ogen) (white) and CD45 (green). Higher magnifications in bI and bII, right panels. Scale bars, 1000μm and 100μm (bI-II), n = 3 mice. c, Time-course of fibrin(ogen) (in green) and CD45 (white) at P7 (n = 3 mice), 3wk (n = 5 mice), and 8wk (n = 20 mice) old mice. Scale bar, 300μm. d, Fibrin(ogen) (green) in Cx3cr1creERT2;CSF1RlslDTR 7-days post-macrophage depletion. Scale bar, 500μm, 150μm (dI), and 8μm (bottom row). e, Scanning electron microscopy of aortae from control and Cx3cr1creERT2;CSF1RlslDTR 14-days post-macrophage depletion (n = 3 mice per group). f, Images of the aortic arch comparing fibrin(ogen) (green) and CD45 (white) in Control and CD11c-/- mice (Scale bar, 300μm and 8μm, n = 4 mice per group). Higher magnifications (fI and fII) on the right. Graph with number of CD45 + cells in the LC of control and CD11c-/- mice. (n = 4 mice per group, Mann-Whitney T-test, ± SD, p = 0.0286, two-tailed, *p ≤ 0.05). g, Schema of littermate control (negative control) and Cx3cr1creERT2;CSF1RlslDTR that received Tam/DTx (24 hr prior) and C57BL/6 EDTA treated mice (positive control). High magnification shown below. VE-Cadherin (white), CD45 (red) (Scale bar, 7μm; n = 3 mice per group). h, Plat (tPA), Plau (urokinase), and Plaur (urokinase receptor) expression in endothelial cells (EC) and MacAIRs from data sets in Extended Data Fig. 2. i, Immunostaining for tPA (white) in the LC and in the thoracic aorta in relation to CD45 (green). Endothelial cells (red) (Scale bar, 10μm; n = 3 mice). j, Expression of plasminogen (Plg) undetected in EC and MacAIRs by scRNA-seq.
Supplementary information
Supplementary Information
Supplementary Figs. 1 and 2.
Supplementary Tables
Supplementary Tables 1–9.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hernandez, G.E., Ma, F., Martinez, G. et al. Aortic intimal resident macrophages are essential for maintenance of the non-thrombogenic intravascular state. Nat Cardiovasc Res 1, 67–84 (2022). https://doi.org/10.1038/s44161-021-00006-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s44161-021-00006-4
This article is cited by
-
Macrophages in cardiovascular diseases: molecular mechanisms and therapeutic targets
Signal Transduction and Targeted Therapy (2024)
-
Heat transfer mechanism in idealized healthy and diseased aortas using fluid-structure interaction method
Biomechanics and Modeling in Mechanobiology (2023)
-
The why and how of adaptive immune responses in ischemic cardiovascular disease
Nature Cardiovascular Research (2022)
-
Aortic-intima-resident macrophages are guardians of arterial health
Nature Cardiovascular Research (2022)
