
Abstract
Seafood holds promise for helping meet nutritional needs at a low climate impact. Here, we assess the nutrient density and greenhouse gas emissions, weighted by production method, that result from fishing and farming of globally important species. The highest nutrient benefit at the lowest emissions is achieved by consuming wild-caught small pelagic and salmonid species, and farmed bivalves like mussels and oysters. Many but not all seafood species provide more nutrition at lower emissions than land animal proteins, especially red meat, but large differences exist, even within species groups and species, depending on production method. Which nutrients contribute to nutrient density differs between seafoods, as do the nutrient needs of population groups within and between countries or regions. Based on the patterns found in nutritional attributes and climate impact, we recommend refocusing and tailoring production and consumption patterns towards species and production methods with improved nutrition and climate performance, taking into account specific nutritional needs and emission reduction goals.
Similar content being viewed by others


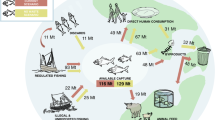
Introduction
Globally more seafood than ever is being produced and consumed and demand continues to grow with increasing wealth and population growth1. In 2017, seafood accounted for 17 percent of the global intake of animal protein1. There is substantial evidence that the health benefits of seafood consumption generally outweigh the potential negative health effects of contaminants or other safety risks2,3. Aquatic ecosystems play a crucial role in meeting human nutrition goals4, as seafood provides substantial amounts of protein, n-3 fatty acids and micronutrients such as vitamin D, vitamin B12, selenium, iodine, iron, zinc and phosphorus. Seafood is also important in the prevention of numerous non-communicable diseases and tackling widespread micronutrient deficiencies5,6,7,8,9, reasons why many governments recommend increased consumption. In addition, potential environmental benefits have been demonstrated from replacing other animal foods with seafood10,11,12. The public debate on future diets is currently strongly focused on the so called ‘green shift’- shifting consumption from terrestrial animal-based to plant-based foods, with far less attention devoted to a possible ‘blue shift’ where aquatic-sourced foods play an increasingly important role. Instead, seafood is often either omitted entirely from discussions or treated simplistically as an undifferentiated whole in studies analysing the combined health and environmental impact of diets13,14,15,16,17. To increase seafood consumption sustainably, better understanding of the performance of this diverse food category is needed.
While food sustainability is complex and multi-dimensional, climate change is one of the most urgent challenges confronting humanity, and greenhouse gas (GHG) emissions are readily quantifiable across production systems, making comparisons between diverse sources possible as long as methods are aligned18. Often but not always, climate impact is correlated with other environmental concerns and in such cases, emission reduction efforts will lead to broader improvement. Studies assessing and comparing the GHG emissions of seafood and other food products typically report emissions per kilogram of product, failing to account for the variation in nutritional value and food function of products. Some studies instead compared products based on portions or protein content19 but did not capture the wider nutritional variation. Recently, nutritional density indices were proposed to describe the nutritional profile of foods more comprehensively when comparing their environmental impacts20,21,22,23. Nutrient indices summarize densities of macro- and micronutrients24 and provide an indication of the extent to which foods contribute to average nutritional requirements25. A 2019 study by Hallström and colleagues estimated the nutritional value and GHG emissions of a range of seafood products representative of Swedish consumption patterns26. The nutrient density score in that study was based on data for 24 nutrients and it related content of each nutrient to the dietary reference intake (DRI) of desirable nutrients, or the maximum recommended intake (MRI) of undesirable nutrients. Care was taken to select and weight seafood products consumed in Sweden with regard to methodological consistency and representative production techniques. In a more recent effort, nutrient data for 12 nutrients was compiled along with available life cycle assessment (LCA) results for 35 food groups based on statistics and market categories27. Another seafood-focused study compiled nutrient content data for five nutrients, along with data on four environmental stressors on an aggregated level of species groups within aquaculture production systems28, without mentioning the large within-group variation in both dimensions26. Another study included seafood from both fisheries and aquaculture production systems29 while also quantifying the environmental impacts of providing the DRI of specific nutrients through different foods, the latter of which was also done by Koehn et al.27.
Here, we build on the work of Hallström et al.26 and analyse GHG emissions associated with production of globally important seafood species relative to their respective nutrient densities. We further refine the nutrient density score with regard to nutrients included, capping procedures and analysing variability both within and between species groups. We expand the species range to cover all species important in global production for which data are available and aggregate our results and present the potentially available global food supply from these species to place each in context of global production, converting liveweight into estimates of edible seafood mass. Although what is considered edible varies with the cultural context, most species are predominantly consumed as fillets and therefore the fillet yield serves as a rough proxy for food supply. We demonstrate how seafood products differ in nutrient density and climate impact both in relation to each other, and to terrestrial animal protein sources. Further, we identify those species that provide the most overall nutrients for the least emissions as well as those that provide the least nutritional value for the highest climate cost. Doing so, we add a nutritional dimension to our understanding of emissions from the seafood sector that can be further adapted to country- and population-specific dietary needs to identify opportunities for seafood to help satisfy human diets within carbon constraints.
Results and Discussion
Seafood more nutritious at lower emissions than terrestrial animal protein sources
At the highest level, we find that while a two-fold variability exists between the average nutrient density scores of major seafood species groups, median GHG emissions vary by over an order of magnitude between sources of seafood—both in terms of species groups but also in terms of how seafood is produced (Fig. 1). The global average performance of all seafood assessed, weighted by species production volume (the two lines in Fig. 1), has a higher nutrient density than beef, pork and chicken and lower GHG emissions than beef and pork. Importantly, the weighted average performance of all seafood species analysed is useful only for comparative purposes—as it does not per se indicate whether this average represents a high or low value. This is true also for the nutrition and environmental data shown in a recent study28, whose relative values do not reveal whether the difference between the best and the worst-performer is small or large.

Relative nutrient density and production-related GHG emissions (i.e. post-harvest emissions are excluded) per edible weight of globally important seafood groups from fisheries (solid colours) and aquaculture (striped) at the point of landing or harvest, along with beef (B), chicken (C) and pork (P). Relative size of seafood group bubbles is proportionate to 2015 global edible weight production volumes, and GHG and nutrient density values are weighted by species. Both GHG values and nutrition scores are log-weighted and positioned relative to the weighted average of all included seafood species (3.7 kg CO2e per kg edible weight and NDS 4.0). For graphs showing species in each species group individually, see Supplementary Figs. 1–8.
Pelagics, wild salmonids and farmed bivalves best performers
Amongst the seafood groups defined, wild-caught salmonids (pink and sockeye salmon) and the small pelagic species (e.g. herrings, mackerels, and anchovies) and farmed bivalves have the lowest GHG emissions per nutrient density ratio (Fig. 1, Table 1), and comprise the top tertile of species considered (Table 1). These are not the most consumed seafood species, though. Harvest of wild salmonids is relatively low and functionally constrained by limited stocks. A large portion of landings from many small pelagic fisheries is currently destined for other uses (e.g. inputs to aquaculture and livestock feeds), largely due to insufficient demand for direct human consumption, but also as a result of incentives created by regulations. Farmed bivalves (e.g. oysters, mussels etc.) are found among top performers in terms of GHG emissions, but provide slightly lower nutrition density. In contrast, crustaceans, both farmed (primarily tropical shrimp species), and wild-caught (various shrimp species, American lobster, etc) and cephalopods all result in higher than average emissions while providing lower than average nutritional scores. Our findings with regard to best- and worst performing species and species groups confirm previous findings26,27,28. Unlike other seafood groups that are defined phylogenetically and presumably are nutritionally more similar9 (see also Fig. 1), species grouped as ‘whitefish’ simply share desirable human consumption characteristics (e.g. firm, pale flesh, mild flavour). As both wild-caught and farmed whitefish represent substantial production tonnages, their group-specific nutritional and GHG emission scores have a commensurate influence on the overall weighted average scores for all seafood products analysed. Despite this, the wild-caught whitefish species considered resulted in the lowest nutrient density scores of all groups assessed (Fig. 1). In all comparisons between species groups and production forms in Fig. 1, it is important to keep in mind that as the observations within each group and methods used to characterize attributes are not fully consistent across data sources, it was not considered feasible to conduct formal statistical testing. The differences observed should therefore be interpreted as indicative based on currently available data.
Large variability within species groups
Each species group consists of 2-10 species with considerable variability within each group either in terms of nutrient density scores, production-related GHG emissions, or both, with larger variation observed in GHG emissions (Fig. 2, Table 1, see also Supplementary Figs. 1–8). The lower variability in nutrient density is partly due to the choice of capping nutrient content at the dietary reference intake (DRI, i.e. nutrient content exceeding the DRI does not influence the nutrition score, see Methods). This choice also reduces the contribution of nutrition to the combined score, which is driven moreso by GHG emissions due to the greater variability in emission rates. Likely reflecting the large diversity of species and production sources contained within the whitefish group, it encompasses the widest species-specific variability of both GHG emissions and nutritional density (Fig. 2).

Nutrient density scores are based on the 21 nutrients common to all species (full bars) and, where possible, 23 nutrients (grey lines) (for nutrients see Methods). GHG emissions of individual seafood species are representative of the dominant production method for each (or weighted if multiple major production method is employed globally). Solid bars indicate species from fisheries, and striped bars species from aquaculture. Comparisons to land-based animal proteins are based on nutritional content of averaged meat cuts for beef and pork, and fillets for chicken. GHG emissions of beef are beyond the scale at 56 kg CO2e per kg edible product.
Within most species groups, the large observed variability in GHG emissions (Fig. 2) mainly determined by the production technique, suggests the potential for substantial reduction opportunities related to production methods and practices. Small pelagics are an exception and exhibit greater variation in nutrient density values than in GHG emissions. While the nutritional content of species can only be influenced to a limited degree (e.g., through changes in feed composition, timing of harvest), substantial opportunities to reduce GHG emissions exist by increasing use of low-emission technologies or inputs (e.g. energy sources, feed inputs), and by rebuilding stocks30,31. In particular, changes in feed composition and use represent major opportunities to reduce emissions from fed aquaculture32. Greenhouse gas emission intensity values reported in Fig. 2 are also influenced by differences in edible yield, which is high for cephalopods (~70–80% from liveweight), but low for bivalves (~15–25% from live weight), with finfish species falling between these extremes. Yields can be maximized through technical or temporal regulations or through improved processing technology, but each species has biological limits. Analysing these parameters on a species group level28 misses out on this variability, which can be larger than the between-group variability26.
Half of seafood species perform better than terrestrial animal protein sources
When compared to land-based animal source foods, 22 of the 41 seafood species whose nutrient density scores could be assessed (54%) and 17 of the 34 seafood species whose production-related GHG emissions could be quantified (50%) performed better than beef, pork and chicken (Fig. 2). Pork performs just below the average of seafood in both dimensions assessed, whereas chicken has a much lower nutrient density, comparable to the poorest performing seafood groups (Fig. 1). Beef scores just under the average seafood nutrient density but results in higher GHG emissions than any other food analysed here (Figs. 1, 2). Nutrient density of seafoods is only lower than land-based animal products for a few species (e.g. some whitefish and Japanese carpet shell; Fig. 2).
Nutrient density of seafood is driven by various nutrients
Our results indicate that salmonids, both wild-caught and farmed, and small pelagics are the most nutrient dense seafoods assessed (Fig. 2, Table 1), which is consistent with previously reported research26,27. Some tuna species and common carp also rank high for NDS21 (tertile 1, Table 1). Most bivalves and cephalopods showed intermediate nutrient density scores (tertile 2, Table 1), whilst most crustaceans and whitefish species ranked low (tertile 3, Table 1).
Vitamin B12, niacin and vitamin D are the nutrients that, on average, made the greatest contribution to nutrient density scores across the seafood species assessed (on average 20, 12 and 9% of NDS21, respectively; Fig. 3). Vitamin B12, in particular, had the greatest impact on nutrient density in 35 of the 41 species assessed for NDS21 (see Supplementary Table 1). Analysing the nutritional value at an overall taxonomic level risks missing important within-group differences and average values may not represent individual species or the group well. For example, our data showed that oysters had the highest calcium content of all species included and that all bivalves were over average in calcium content, whereas a recent study28 concludes that bivalves contain less than average calcium across species groups. Despite differences among individual species, patterns can be identified between seafood groups in terms of nutrient composition (Fig. 3). For example, n-3 fatty acids are the key contributor to NDS21 amongst the nutrient dense salmonids (especially when farmed) and small pelagics (Fig. 3). Farmed whitefish and wild salmonids are particularly rich in vitamin D, whereas cephalopods, mussels and crustaceans (both farmed and fished), have a high copper content (Fig. 3). Partly due to the higher vitamin D content, farmed whitefish performed better than most of fished whitefish species assessed. Due to the role whitefish occupies in current consumption, this aspect would probably deserve further investigation by broadening the analysis to include a larger number of farmed whitefish species (e.g. more carp species).

Nutrient profiles of seafood groups expressed as percentage contribution to the nutrient density score NDS21, calculated as weighted averages of included species within groups based on edible production volumes. Desirable nutrients are visualized if contributing ≥10% of nutrient density for at least one group. Other includes all remaining desirable nutrients not reaching the cutoff value.
Nutrient density is a composite index to which desirable and undesirable nutrients contribute positively and negatively to the final score. Our analysis confirms that seafood is a source of animal protein that delivers minimal quantities of sodium and saturated fat, nutrients that are indeed considered ‘undesirable’ from a public health perspective. Bivalves are the only group considered in which sodium can be regarded as non-negligible (Fig. 3).
The importance of individual nutrients for adequate nutritional intake and overall health differs between populations and population groups. To account for such differences it has been suggested that nutrient density scores should be tailored to the targeted population assessed23. Here we intentionally avoid making such local adjustments in order to be able to describe overall patterns of climate impact in relation to nutrient content across species. Results therefore should necessarily be considered within this context and goal in mind. Any future applications of the approach to specific populations should attempt to account for the dietary needs locally or of specific population sub-groups (defined by age, gender or socioeconomic parameters), as well as local availability of seafood products and their source.
The nutrient profiles of many species analysed revealed very high concentrations of a few nutrients, often well above their DRI. As described, capping was applied in the NDS calculation even though this flattened the nutritional variability among species. If not capped to 100% of DRI, disproportionately high scores would be seen even in products providing low levels of most nutrients and high levels of one or a few nutrients (e.g. Japanese carpet shell). Vitamin B12 was the nutrient that most often exceeded the DRI, in 26 of the 41 seafood species assessed, with contents up to almost 25 times the DRI in the flesh of some species (see Supplementary Table 2 for data and Supplementary Discussion 1).
In dietary guidelines, vitamin D, n-3 fatty acids, selenium and iodine are often identified as nutrients from seafood of special importance in human diets2. Nutrient profiles are highly variable between species (Fig. 4) and two types of seafood with the same nutrient density score can make markedly different contributions to the intake of specific nutrients. By assessing the role of individual nutrients beyond their contribution to the nutrient scores, potentially important sources of specific nutrients can be identified despite their relatively small concentrations (Fig. 4). Largehead hairtail, for example, only displays an intermediate nutrient density score (Table 1) but is the richest source of n-3 fatty acids among the analysed species (Fig. 4, Supplementary Table 2). Common carp and Nile tilapia are farmed whitefish species with markedly different nutrient densities (Fig. 2, Table 1, Supplementary Table 1), but both containing relatively high concentrations of vitamin D (Fig. 4, Supplementary Table 2). Composition data on selenium and iodine were only available for a subset of the studied species (36 out of 41). Amongst these, tunas were all excellent sources of selenium, but even some crustaceans like Gazami crab exceed the DRI for this element (Fig. 4). Iodine is a nutrient that many humans are deficient in globally33. Some species, like American lobster, Atlantic cod and haddock, are good sources of this micronutrient (Fig. 4), despite not scoring well in terms of NDS21 (Fig. 2, Table 1). With the exception of these species, the addition of selenium and iodine to an NDS23 did not markedly affect the overall patterns of nutritional performance of the species considered (see NDS23 in Fig. 2 and Supplementary Table 3). In order to capture the full potential of seafood species in the human diet, it is, however, advisable to include these minerals in nutrition evaluations when their concentrations are available. Additionally, it is important to be aware that the NDS only relies on the nutrient content as a measure of nutritional quality. Other aspects related to the potential health effect of consuming seafood, such as nutrient bioavailability, food matrix effects, content of other bioactive or toxic compounds are not captured by this method.

Nutrient profiles of six seafood species as percentage contribution of 100 g raw edible flesh to the dietary reference intake: Atlantic cod (A), Common carp (B), Albacore (C), Largehead hairtail (D), American lobster (E), and Gazami crab (F). The twenty-one nutrients included in the NDS21 formula are represented here, plus selenium and iodine (not available for Common carp or Largehead hairtail; iodine not available for Gazami crab). Nutrients are grouped by proteins and fats (orange), minerals (blue), and vitamins (green). Values are displayed as relative area of the pie slice, with a maximum value (full slice) representing a 100% contribution. All nutrients with the exception of n-3 are capped to 100% when exceeding the DRI.
Our results are broadly consistent with those of similar analyses undertaken for seafood consumed in Sweden26 despite using distinct data sources, with pelagic and salmonid species performing best. Where differences exist (e.g. the ranking of oysters or lobster), they result from differences in methodological choices made for calculating the nutrient density (e.g. capping of nutrients and a different selection of nutrients), and reliance on a wider suite of nutrient composition data sources. Koehn et al.27 also identified small pelagics, and salmon as best-performers, despite large differences in the modelling of the nutrient index, e.g. excluding the nutrients that were most important in this analysis (Vitamins B12, D and niacin) as well as content of undesirable nutrients, calculating the average instead of the sum of nutrient content per DRI ratios and letting content higher than DRI influence the index for all nutrients.
Nutrient content assessment methods and reporting for seafood species varies widely both within and between nutrition databases and likely affected the resulting nutrient density scoring and combined nutrition-climate impact assessment. Relatedly, nutrient densities for each species are calculated based on single observations, rather than averages from multiple databases29. Additionally, many globally important species (e.g., carp) could not be included due to lack of detailed nutrient composition which points to important data gaps. This suggests the need to include more seafood products in methodologically harmonised, public food composition databases.
Pelagics have lowest greenhouse gas emissions, crustaceans highest
Of the 41 seafood species for which an NDS21 value could be assessed, we were able to quantify production mode specific (e.g. fished or farmed) GHG emissions for 34 species (Supplementary Table 4). Emission intensities of individual species and major patterns of relative emissions associated with seafood groups (Fig. 2) are broadly consistent with previous findings in the study of Swedish consumption26. Differences occur when globally important production technologies, modelled here, differ markedly from the specific sources known to supply Swedish consumption. Moreover, emission data that were not available at the time of the previous analysis were used for a few species (e.g. oyster and Atlantic salmon).
Wild-caught crustaceans and some farmed whitefish, tunas, farmed salmonids and cephalopods had the highest GHG emission intensities. Both farmed bivalves assessed (mussels and Pacific cupped oysters) together with all eight small pelagic species assessed, pink and sockeye salmon and Alaska pollock, all had emission intensities far below that of chicken (Fig. 2), while no seafood species approached the scale of GHG emissions from beef.
Five of the seven species for which a GHG emission value could not be identified or characterised directly are farmed bivalve species (Jackknife clam, Japanese carpet shell, farmed scallop, green and Chilean mussels) which points to a major gap in the LCA and related GHG emission accounting literature. This is unfortunate given the promising performance of bivalve species described previously in terms of emissions11,32 and nutrition26,28. As GHG emission values were available for blue mussel culture, it was used to characterise cultured green mussels and Chilean mussels, as a reasonable first approximation of their actual emission intensities. The five species (four farmed and one fished) that could not be assigned a GHG emission value are all produced primarily in China (Fig. 2).
Fuel and feed dominate seafood emissions
Fuel combustion during fishing is the primary source of GHG emissions from capture fisheries, with fuel use intensity (FUI) rates strongly influenced by the fishing gear employed and the relative abundance and catchability of stocks30,31,34,35. Consequently, amongst the fished species assessed here, those with higher emission intensities were typically landed using more fuel-intensive fishing methods or targeted species that are less abundant (Supplementary Table 5). For example, amongst the four tuna species assessed, those for which a larger proportion of total landings are caught using hook and line gears (bigeye, albacore), have higher emission intensities than those species primarily caught using purse seines (yellowfin, skipjack). The case of Alaska pollock presents an interesting example of a species in the whitefish group that performs very well due to a relatively fuel-efficient fishing method, pelagic trawling, resulting in a high catch rate and a remarkably low emission intensity, confirmed by very recent data36.
Sources of GHG emissions from aquaculture production are far more diverse37,38 though tend to be higher when species in culture are fed, especially if also substantial energy inputs are required to maintain culture water quality (e.g. aeration, waste removal, chilling, etc). Amongst the farmed species assessed, Amur catfish had the highest GHG emission intensity while farmed mussels and oysters, both unfed in culture, had the lowest (Supplementary Table 4). Relatively low edible yield rates are a secondary explanatory factor behind the relatively high emissions from crustacean and catfish production.
Nutrition a more relevant basis for comparison than liveweight
Communicating the environmental performance of seafood products based on their nutrient density more completely captures the function of these products relative to the performance of the systems providing them. It improves upon comparisons made on the basis of edible weight, which in turn, is a substantial improvement over comparisons made on liveweight. Importantly, this nutritional lens not only facilitates comparisons between species using a more product-relevant basis, but substantially changes the result of those comparisons. In fact, when scaling up results to annual global production volumes of the species and species groups that we have analysed here, the most important groups in terms of total liveweight mass produced are, in descending order: bivalves (oysters and mussels), farmed whitefish species, small pelagics and wild-caught whitefish species (Fig. 5). Wild-caught salmonids, crustaceans and cephalopods represent the smallest liveweight tonnage species groups produced (Fig. 5). On the basis of mass of edible product available, small pelagics and farmed whitefish dominate. This is because small pelagic species have higher edible yield rates (53–62%, Supplementary Table 6) than most other groups but in particular compared to farmed whitefish species (37–45%) and bivalves with the lowest edible yields of all groups (15–24%). Multiplying edible mass of each species group, calculated using species-specific edible yield factors, by the weighted average NDS21 score of species within each group, the importance of small pelagics in terms of potential human nutrition increases further (Fig. 5). Other groups whose relative nutritional importance to humans increases when moving from volumes to nutrition density are large pelagics, farmed whitefish and salmon, while the relative importance of fished whitefish, cephalopods, farmed and wild-caught crustaceans, and bivalve groups are all reduced from their contributions to total edible volume. When translated into GHG emissions, the differences are even more pronounced with the three top-performing species groups, small pelagics, wild salmonids and bivalves, together representing 35% of the available nutrition density while only contributing 6% of production-related GHG emissions across all species assessed. In contrast, farmed and fished crustacean species represent 8% of total seafood nutrition density and produce 17% of total emissions estimated across all species assessed. All data used to produce graphics is presented in Supplementary Data 1 and Supplementary Material.

Contribution of seafood groups to global production volumes in 2015 expressed on a live and edible weight basis (for species-specific edible yields see Supplementary Table 6), nutrient density (NDS21) and GHG emissions, all weighted by species within each seafood group. Solid groups consist of wild-caught, striped ones of farmed species.
In addition to strategies to improve the nutritional output from individual seafood systems noted above (i.e. changes in feed, timing of harvest), larger-scale opportunities exist to increase the nutritional performance of seafood systems more broadly. Policies and technological innovation that increase direct consumption of landings from small pelagic fisheries could result in dramatic improvements in the nutritional output of global fisheries while limiting emissions. Although the proportion of global seafood production destined for non-food purposes is declining1, in many settings utilisation of small pelagic species for feeds is still incentivised. For example, policies aiming to concentrate fisheries to fewer, larger vessels landing larger volumes result in reduced catch quality, and a larger proportion of catches ending up as feed, also because of limited capacity to process these landings before quality deteriorates further.
Policy changes that facilitate greater utilisation of landings for food could take many forms (e.g. quota reallocations, distributing harvest opportunities in time or space, improving on-board and in storage product conservation, differential resource rents based on product destiny, etc.), but will need to take into account the unique characteristics of individual fisheries and their settings. Moreover, for any substantial change to be successful, many actors need to be involved beyond fish harvesters, including the food industry and retailers who are going to need to develop, produce and sell new products. Efforts will be needed to understand consumer attitudes towards these species whose top performance in both dimensions, nutrition and climate, would call for using a much higher proportion directly as food than is the case today. In addition, food product innovation designed to increase the utilization of fish or by-products from fish in supplements could contribute to making seafood more accessible to consumers in both high- and low-income country settings1,39. Separately, policies that facilitate the expansion of mussel farming, together with efforts to increase mussel consumption (e.g. dietary advice recommending mussels, sponsored cultural events featuring mussels, developing convenient and affordable mussel-based food products etc.) would also improve the combined nutritional and climate impacts of seafood consumption in general. While macroalgae species were not included here, a previous analysis of environmental stressors did32 and found that seaweed was a promising low-impact group of species. There are big knowledge gaps related to the content and bioavailability of nutrients from seaweeds, as well as about their content of undesirable substances, but research is ongoing on these topics. Species from unfed, low-trophic aquaculture have been identified to have a large potential as future foods10,29,40.
Conversely, just as dietary advice in many countries recommends against intake of red and processed meat, advice related to seafood consumption could indicate types to avoid based on lowest nutritional value at highest emissions. For example, the European Commission is, as part of its Green Deal policy, developing nutrition and sustainability labelling for food products in the coming years and as more data become available this type of analysis will become easier, more robust and informative in that kind of effort. From a global perspective, it may even be wise to promote the most nutritious forms of seafood in nutrient-deficient populations and communities, even when production results in relatively higher emissions, while in populations not at risk for nutrient deficiencies, consumers could give more attention to the emissions than to nutritional content when choosing seafood products for their diets. In fact, it is in nutrient-deficient population groups that any increase in seafood consumption would have the most positive effects for human nutrition. Our results show that nutrition-based functional units can be a valuable complementary tool when comparing the environmental performance across seafood species and other foods.
Seafood statistics and research often defaults to a production perspective, with even consumption being measured in liveweight mass1. If human nutrition is the ultimate objective of fisheries and aquaculture, it is important that outputs are understood and evaluated on a nutritionally relevant basis particularly given the diversity of species involved4 and maximising the nutritional output while minimising environmental costs of seafood provisioning should be a guiding principle for policy-making in these areas41. As both fisheries and aquaculture face many environmental challenges—in terms of sustainable utilisation of stocks, reduction of by-catch and impacts on local ecosystem structure and function, nutrient enrichment, and disease amplification—restricting the analysis of sustainability to GHG emissions may seem very limited. However, it is not uncommon to see biotic impacts aligning with climate impacts as carbon-intensive fishing methods often also result in larger ecosystem impacts30,31,42,43,44. In such cases, relative rates of GHG emissions can serve as a rough indicator of broader environmental sustainability, although there are important exceptions when GHG emissions and broader environmental impacts do not align, e.g. when comparing open and closed aquaculture systems45. Ideally, nutrition data and key emissions drivers, fuel use in fisheries and feed use and composition would be collected and made available in a standardised way to facilitate and increase the robustness of this type of synthetic analysis and comparisons across species, species groups and production technologies. This would also enable monitoring of performance over time, which could help guide us into a future of nutritious foods at low environmental costs.
Conclusions
Results confirm that seafood is a relatively low climate impact source of highly nutritious food. This suggests substantial emission reduction gains are possible by shifting protein sources while simultaneously achieving nutritional benefits. In addition, amongst seafood species, even within the same species group or species, there are substantial differences in the climate performance, depending on production methods. Increased consumption of small pelagic species, wild-caught salmonids and bivalves would reduce greenhouse gas emission of seafood consumption considerably, while improving nutritional benefits, in particular if replacing red meat. While many hurdles need to be overcome, we have the potential to reshape seafood production and consumption towards species that optimise nutrition while minimising climate emissions both in terms of the suite of which species are produced and how. As a next step, such recommendations could be designed for specific population groups to meet their nutritional needs and emission reduction goals.
Methods
Fish and shellfish species selection
We extracted liveweight mass production data for 2015 for over 200 wild-caught and farmed seafood species from the global fisheries and aquaculture production data compiled by the FAO Fisheries and Aquaculture Department and made available through the software FishStatJ. From that initial list, we sequentially excluded those records that:
-
i.
could not be categorised as one of five major animal groups of interest (i.e. freshwater fish, diadromous fish, marine fish, crustaceans, and molluscs);
-
ii.
reported landings only at the Family or Genus level or in any other undifferentiated grouping;
-
iii.
were not among the top fifty remaining species globally by liveweight production after previous filtering; or
-
iv.
could not be linked to sufficiently detailed nutritional composition data for the specific species or a closely related proxy species.
Several species, despite substantial 2015 production volumes, could not be included due to lack of nutrient composition data. Examples include a number of carp species (grass carp, bighead carp, etc.) though sufficient data were available to retain common carp (Cyprinus carpio).
We allowed some exceptions from these criteria. A few additional seafood species were included despite not ranking amongst the top species by 2015 production volume. These were either species belonging to a species group of particular interest from a nutritional perspective (e.g. blue mussel, Mytilus edulis), or species analysed previously26 (e.g. haddock, Melanogrammus aeglefinus) for the purpose of comparing nutrition data for these species across databases. Two higher taxonomic groups, the class Cephalopoda (squids and octopodes) and the family Pectinidae (scallops) were also included as aggregated groups as these important food sources would otherwise have been absent from our analysis. The nutritional profile of both these two larger groups were characterized using the nutritional composition data of a major member species. The species assessed, the top-producing countries for each species and the reason for their inclusion is reported in Supplementary Table 6.
Overall, a total of 41 seafood species or groups were included in the study of which 14 were farmed and 27 fished, together accounting for 27% of global fish and shellfish production volume (including seaweeds) in 2015.
Assessment of nutrition density
We identified relevant food composition databases containing detailed nutritional data by species using the World Nutrient Databases for Dietary Studies on-line search tool (https://foodsystems.org/resources/wndds/). We selected the uFishJ food composition database as the preferred source of nutrient composition data based on data availability and quality. When information on individual nutrient content was lacking in uFishJ, data were supplemented from the Canada Nutrient File, the Japan Standard Tables of Food Composition or the Swedish Food Composition Database. All nutrient composition data assembled were based on analyses of uncooked edible products (after the exclusion of inedible parts e.g. shells, bones and skin), see Supplementary Table 7 for further detail and references.
A composition profile including 21 nutrients, of which 19 were considered desirable (protein, n-3 fatty acids, vitamins A, D, E, B6, B12, thiamin, riboflavin, niacin, panthotenic acid, folate, calcium, copper, iron, potassium, magnesium, phosphorous, zinc) and two non-desirable (saturated fatty acids and sodium) was considered sufficiently inclusive of the major nutritional attributes of seafood for the purpose of comparing different seafood products, as well as covering most of the nutrients for which a DRI value exists. The choice of nutrients for inclusion was based on those selected in an earlier study26 with the following exceptions. Dietary fibre and vitamin C were excluded as they are not important for comparison between seafood products and retaining them would have further restricted the number of species with complete data availability. Pantothenic acid (vitamin B5) was, however, included in this analysis, based on data availability in uFishJ and amounts that were judged noteworthy for some of the studied products. Selenium and iodine are two nutrients that are important in human diets for which seafood is an important source. However, as selenium and iodine content were only available for a subset of the species included, we excluded them from the main analysis to avoid limiting the number of species further. We subsequently undertook a second nutrient density analysis for the smaller number of species for which selenium and iodine data were available bringing the total number of nutrients considered in this second analysis to 23. Details on nutrients included in the analysis and the choice of food composition databases appear in Supplementary Tables 7, 8. The nutrition data extracted from the databases for the species included is provided in Supplementary Table 2.
As nutrients play different roles in human nutrition and occur in very different concentrations, it is necessary to construct a composite metric or score for products being compared. Following Hallström et al.26 whose analysis was based on the Nutrient Rich Food model24, we calculated a nutrient density score (NDS) for each seafood product as the sum of the fractional contribution to the DRI values of desirable nutrients minus the sum of the fractional contribution to MRI values of non-desirable nutrients per 100 g of a food product (Eq. 1).
Where x is the number of desirable nutrients, y is the number of non-desirable nutrients, Nutrient i and j is the content of nutrient i (desirable nutrients) or j (undesirable nutrients) per 100 g of raw edible seafood product.
In line with the global perspective of the present study and recommendations23, reference values for DRI and MRI were taken from the international Codex Alimentarius46, except for the reference value for n-3 fatty acids which was derived from an expert consultation report47 (see Supplementary Table 8).
The NDS algorithm in Eq. 1 was chosen among seven methods assessed by Hallström et al.26, as it balanced the influence of desirable and non-desirable nutrients and avoided generating negative values which can be difficult to interpret and compare. Calculating an average ratio across nutrients, as done by Koehn et al.27 and explored but dismissed by Hallström et al.26 is more useful in cases when only desirable nutrients are included, as in Koehn et al.27. Additionally, the use of an NDS assessed per 100 g of seafood product, rather than 100 kcal, was considered appropriate when comparing products within a food category (i.e. seafood products), which do not differ markedly in water content and are usually consumed in similar portions (but may differ considerably in fat content and, consequently, calories—a difference partly captured by the nutrient score itself). An important methodological choice when constructing the NDS for each product is whether to limit or ‘cap’ the fractional contribution value of individual desirable nutrients to 100% of DRI for those nutrients whose concentrations exceed the DRI (i.e. vitamin B12 and vitamin D in this study). Alternatively, an NDS can be constructed of all fractional contribution values regardless of whether one or more exceed 100% of DRI (as e.g. done in Hallström et al.26). Results analysed with capping are presented here, but a comparison with uncapped NDS values and ranking of products is available in Supplementary Tables 9, 10. Capping was applied to desirable nutrients, with the only exception being n-3 fatty acids, where the DRI is given in terms of minimum energy % of the total dietary calories. Due to the global focus of this study, all nutrients contributed equally to the nutrient density score and no weighting was applied in the NDS calculation, a possibility in follow up studies with a focus on specific population groups (e.g. defined by age, gender or socioeconomic parameters).
We calculated the NDS based on 21 nutrients (NDS21) for all 41 species and groups in the larger dataset (Supplementary Table 1) and additionally based on 23 nutrients (NDS23) for the 34 species for which selenium and iodine content values were available. For each seafood product, we calculated the percentage contribution of individual nutrients to the final NDS and identified the top contributing nutrients (where individual nutrients represent ≥10% of the NDS of the product).
In order to compare the performance of seafood to other animal protein sources, we additionally calculated NDS21 for representative products from beef, chicken and pork. For this comparative analysis, we obtained nutritional profiles for raw meat from the Canada Nutrient File database (Supplementary Table 7) and referred to an average of cuts for beef and pork and to an average of skin-on fillet for chicken.
Production-related greenhouse gas emissions in aquaculture
Greenhouse gas emission intensities, as kg CO2 equivalents (CO2e) per kg liveweight, of farmed species were selected from a review of LCA studies48, which was updated with more recent research. To determine the most representative method of production for each farmed species, we undertook internet searches, reviewed the FAO species sheets, and consulted with aquaculture experts in major producing regions. We then reviewed a total of 50 peer-reviewed aquaculture LCA studies, representing a combined 104 case studies, from which the most relevant and representative matches were selected based on species, production method, country of origin, and methodological consistency in underlying studies. In a recent study, we discuss suitable methods for food LCA aggregation18 and conclude that methodological consistency between studies is key for meaningful outcomes. It also recommends that key drivers of the issue studied, here greenhouse gas emissions, should be taken into account when defining groups rather than attributes like taxonomy that do not drive impacts. Here, emission drivers are factored into the data behind each species by weighting each species based on relative contribution of production method, but we present the results using a taxonomic grouping (mainly species), because of the second focus on nutrition, where species is a relevant differentiator. Also, an approach was taken where the most representative study was selected for each combination of species and production method rather than aggregating all available data to avoid common errors made when food LCA data are aggregated18. Still there are methodological inconsistencies amongst the LCA studies we used and efforts were made to minimize the potential impact of combining results of studies employing different allocation strategies through selection of studies and adjusting to a common functional unit. Ultimately, nine published aquaculture LCAs were identified as representative of dominant commercial production practices and whose methods were consistent for analysis (Supplementary Table 4).
GHG emission intensities derived from selected studies were inclusive of life cycle emissions from feeds originating in fisheries and agriculture and on-farm activities but did not include emissions associated with land use change (LUC), for which data were inconsistently available from published LCA studies and results varied widely with methods and models. While inclusion of LUC emissions would have rendered many studies incomparable due to using different LUC- modelling approaches, it is important to note that its exclusion may substantially underestimate the climate impact of some systems, notably those associated with deforestation for feed production or farm siting49. To harmonize methods in data used, GHG emissions from LUC were subtracted from total emission when these were accounted for. Production volumes of all green and blue mussels belonging to the genera Mytilus and Perna were grouped in the modelling of GHG emissions due to lack of studies of Chilean and New Zealand Green mussels, although nutritional data were available for the three species individually. Neither carbon sequestration nor release in the process of mussel shell formation was accounted for. When several commercially relevant (i.e. non-experimental or niche) production systems for a species were included in a reviewed study and selected as a best match to inform our characterisation of production-related GHG emissions, a weighted average farmgate GHG emission intensity value was calculated using country-level, method-specific liveweight mass of production as a weighting factor. In most cases, however, we were able to select the production system representing the dominant production volumes globally.
Production-related greenhouse gas emissions in fisheries
While a growing number of LCAs are also available for wild capture fisheries, far more species- and gear-specific data are available from assessments of fishing vessel fuel consumption, and fuel use has typically been identified as the primary—often well in excess of 75%—source of emissions up to point of landing in detailed LCA studies of fisheries31,50. We therefore followed the approach established by35,50 and previously implemented26 to base wild capture fishery emissions estimates on rates of fuel use per tonne of round weight landings.
Species-specific primary fishing methods together with aggregate tonnages landed for the wild-caught species amongst the 41 candidate species and groups were taken from an analysis characterising species-specific landings by gear type globally in 201451 (Supplementary Table 5). Each species was associated with one or more of several major gear types (e.g. bottom and pelagic trawls, gillnets, longlines, purse seines, etc.) and weighted according to relative share of landings by each gear type. We then cross-referenced the resulting list of species and gear combinations with species- and gear-specific records of fuel use intensity, measured in liters per tonne, in the Fisheries Energy Use Database (FEUD)34. Similar to Gephart et al.32, we calculated the median value of all records matching species and gear, truncating records to only include those since 1990. Where species-specific FUI values were not available, values for related species for which effort, environment, and behaviour were expected to be similar, were selected instead. Landings reported for other gear types, including unknown and ‘small-scale’ were all summed and accounted for as having the weighted average emission intensity of the specified methods used for that species, as it was impossible to match them with sufficiently gear-specific fuel use data. The weighted average FUI for each species (Supplementary Table 5) was converted to life cycle GHG emissions (i.e. including both production and combustion of fuel) at a rate of 3.3 kg CO2e per liter, after which a correction was made to account for the roughly 25% of production-related emissions assumed to come from non-fuel sources such as vessel and gear-provision related emissions and refrigerant loss35.
Combined GHG emissions per nutrient density score
Lastly, we calculated an integrated indicator (GHG divided by NDS21) as the ratio between the climate impact, expressed as kg CO2e per kg edible seafood, and NDS21. The combined index ranks seafood products according to their climate impact per kg related to their nutrient density, hence allowing the identification of the species with the lowest climate cost delivering most nutrition.
In order to better estimate the global impact of seafood production to human nutrition and climate, a weighted climate emissions per nutrition ratio was obtained by multiplying NDS21 values for the 2015 edible production volumes.
Comparison of performance between species and with terrestrial animal proteins
Of the 41 species included in the nutritional analysis, we ultimately derived production-related GHG emissions estimates for 34. Excluded species included one wild-caught species (Gazami crab) and four farmed species for which, to date, no appropriate farmgate GHG emission data from Chinese production were identified in the literature (scallops, red swamp crawfish, Japanese carpet shell and jackknife clam/Constricted tagelus).
To provide a nutrition-relevant common basis of comparison, we transformed all production-related GHG emissions on a liveweight basis to emissions per unit of edible product by applying species-specific edible yield factors52 complemented with yield factors from additional sources such as food composition databases for species where data were lacking or (in the case of Atlantic salmon) for coherence with the GHG data source used (see Supplementary Table 6 for yield data used per species). All production-related emissions were allocated to the edible portions of each species, i.e. no co-product allocation was applied and the by-products are hence considered as waste, which in reality is not the case as an increasing proportion of processing by-products are further utilized. Only production-related emissions were modelled, as these generally dominate supply chain emissions. Seafood is traded around the world in complex patterns and transport emissions will depend heavily on the location of the consumer in relation to the producer, and transport modes used and any general data on transport distances and modes per species are not available. Hence, neither processing- or distribution-related emissions were included as these are highly idiosyncratic to specific post-production supply chains. Details of GHG emission characterisation steps and results are available in Supplementary Tables 4, 5.
To put seafood in perspective, GHG emissions estimates for terrestrial animal source foods (beef, pork and poultry) were also included. We based our estimates for these products on the data compiled by Poore and Nemecek19, following their weighed mean calculation from their assembled food LCA studies. To maintain consistency with our analysis of farmed seafood production, only feed- and production-related emissions were included and we subtracted the emissions associated with LUC from the terrestrial protein GHG emission dataset19. The methodological alignment undertaken by the authors included limiting reported emissions where co-product allocation was conducted using economic values of outputs (e.g. milk or meat) or inputs to feeds, similarly to some of the studies used to characterize emissions from farmed seafood species. The conversion to edible meat was made in the same way as for seafood, assuming no use of processing by-products.
Data availability
Nutrition data from four publicly available food composition databases was used, see Supplementary References: [16] (FAO, (2016). FAO/INFOODS Global food composition database for fish and shellfish – version 1.0 (uFiSh1.0) FAO/INFOODS Global Food Composition Database for Fish and Shellfish : Data for policy | AIMS) [17] Ministry of Education, Culture, Sports, Science and Technoloy (MEXT), Office for Resources, Policy Division Science and Technology Policy Bureau, Japan. Standard Tables of Food Composition in Japan. Seventh Revised edn. (2015) http://www.mext.go.jp/en/policy/science_technology/policy/title01/detail01/1374030.htm [18] Health Canada, (2015). Canadian Nutrient File. https://food-nutrition.canada.ca/cnf-fce/index-eng.jsp [21] SFA, (2022). Livsmedelsdatabasen (“The Food Database”). Swedish Food Agency. https://www7.slv.se/SokNaringsinnehall. Other key data, originating in scientific publications, is provided in Supplementary Tables 1–10 as well as Supplementary Data 1.
References
Food and Agricultural Organization of the United Nations. The State of the Worlds Fisheries and Aquaculture 2020—Sustainability in Action (FAO, 2020).
EFSA (European Food Safety Authority Scientific opinion on health benefits of seafood (fish and shellfish) consumption in relation to health risks associated with exposure to methyl mercury. EFSA J. 12, 3761 (2014).
Food and Agricultural Organization & World Health Organization of the United Nations. Report of the joint FAO/WHO Expert Consultation on the Risks and Benefits of Fish Consumption, Rome, 25–29 January 2010. (FAO, 2011).
Bernhardt, J. R. & O’Connor, M. I. Aquatic biodiversity enhances multiple nutritional benefits to humans. Proc Natl. Acad. Sci. USA 118, e1917487118 (2021).
Hicks, C. C. et al. Harnessing global fisheries to tackle micronutrient deficiencies. Nature 574, 95–98 (2019).
Kawarazuka, N. The Contribution of Fish Intake, Aquaculture, and Small-Scale Fisheries to Improving Nutrition: A Literature Review. (The WorldFish Center, 2010).
Khalili Tilami, S. & Samples, S. Nutritional value of fish: lipids, proteins vitamins and minerals. Rev. Fish Sci. Aquac 26, 243–253 (2018).
Lund, E. K. Health benefits of seafood: is it just the fatty acids? Food Chem. 140, 413–420 (2013).
Vaitla, B. et al. Predicting nutrition content of ray-finned fishes using phylogenetic information. Nat. Commun. 9, 3742 (2018).
Costello, C. et al. The future of food from the sea. Nature 588, 95–100 (2020).
Hilborn, R., Banobi, J., Hall, S. J., Pucylowski, T. & Walsworth, T. E. The environmental cost of animal source foods. Front. Ecol. Environ. 16, 329–335 (2018).
Scarborough, P. et al. Dietary greenhouse gas emissions of meat-eaters, fish-eaters, vegetarians and vegans in the UK. Climate Change 125, 179–192 (2014).
Avadí, A. & Freon, P. A set of sustainability performance indicators for seafood: direct human consumption products from Peruvian anchoveta fisheries and freshwater aquaculture. Ecol. Indic. 48, 518–532 (2015).
Bogard, J. R., Farmery, A. K., Little, D. C., Fulton, E. A. & Cook, M. Will fish be part of future healthy and sustainable diets? Lancet Planet. Health 3, e159–e160 (2019).
Farmery, A. K., Hendrie, G. A., O’Kane, G., McManus, A. & Green, B. S. Sociodemographic variation in consumption patterns of sustainable and nutritious seafood in Australia. Front Nutr 5, 118 (2018).
Seves, S. M. et al. Sustainability aspects and nutritional composition of fish: evaluation of wild and cultivated fish species consumed in the Netherlands. Climate Change 135, 597–610 (2016).
Tlusty, M. F. et al. Reframing the sustainable seafood narrative. Global Environ. Change 59, 101991 (2019).
Ziegler, F., Parker, R. W. R. & Tyedmers, P. Methods matter: Improved practices for environmental evaluation of dietary patterns. Global Environ. Change 73, 102482 (2022).
Poore, J. & Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 360, 987–992 (2018).
Bianchi, M. et al. Systematic evaluation of nutrition indicators for use within food LCA studies. Sustainability 12, 8992 (2020).
Grigoriadis, V., Nugent, A. & Brereton, P. Working towards a combined measure for describing environmental impact and nutritive value of foods: A review. Trends in Food Sci. Technol. 112, 298–311 (2021).
Hallström, E., Davis, J., Woodhouse, A. & Sonesson, U. Using dietary quality scores to assess sustainability of food products and human diets: a systematic review. Ecol. Ind. 93, 219–230 (2018).
McLaren, S. et al. Integration of Environment and Nutrition in Life Cycle Assessment of FoodItems: Opportunities and Challenges (FAO, 2021).
Drewnowski, A. Defining nutrient density: development and validation of the nutrient rich food index. J. Am. College. Nutr. 28, 421–426 (2009).
WHO. Nutrient profiling: Report of a WHO/IASO Technical Meeting, London, United Kingdom, 4–6 October 2010 (World Health Organization of the United Nations, 2011).
Hallström et al. Combined climate and nutritional performance of seafoods. J. Clean Prod. 230, 402–411 (2019).
Koehn, J. Z., Allison, E. A., Golden, C. E. & Hilborn, R. The role of seafood in sustainable diets. Environ. Res. Lett. 17(2022), 035003 (2022).
Gephart, J. A., Golden, C. D. Environmental and nutritional double bottom lines in aquaculture. OneEarth 5, 324–328 (2022).
Parodi, A. et al. Future foods: towards a sustainable and healthy diet for a growing population. Nat. Sustain. 1, 782–789 (2018).
Driscoll, J. & Tyedmers, P. Fuel use and greenhouse gas emission implications of fisheries management: the case of the New England Atlantic herring fishery. Mar. Pol. 34, 353–359 (2010).
Ziegler, F. et al. Expanding the concept of sustainable seafood using Life Cycle Assessment. Fish Fish 17, 1073–1093 (2016).
Gephart, J. A. et al. Environmental impacts of blue foods. Nature 597, 360–365 (2021).
Bailey, R. L., West, K. P. Jr & Black, R. E. The epidemiology of global micronutrient deficiencies. Ann. Nutr. Met. 66, 22–33 (2015).
Parker, R. W. R. & Tyedmers, P. H. Fuel consumption of global fishing fleets: current understanding and knowledge gaps. Fish Fish 16, 684–696 (2015).
Parker, R. W. R. et al. Fuel use and greenhouse gas emissions of world fisheries. Nat. Clim. Change 8, 333–337 (2018).
Zhang, X., Kotin, A., Zgola, M. Life Cycle Assessment of wild Alaska pollock. Final ISO Report. (Quantis US, 2021)
Bohnes, F. A., Hauschild, M. Z., Schlundt, J. & Laurent, A. Life Cycle Assessments of aquaculture systems: a critical review reported findings with recommendations for policy and system development. Rev. Aquac. 11, 1061–1079 (2019).
Henriksson, P. J. G., Guinée, J. B. & de Snoo, G. R. Life Cycle Assessment of aquaculture systems—a review of methodologies. Int J LCA 17(3), 304–313 (2012).
Egbi, G. et al. Impact of cowpea-based food containing fish meal served with vitamin C-rich drink on iron stores and hemoglobin concentrations in Ghanian schoolchildren in a malaria endemic area. Food Nutr. Bull. 36, 264–275 (2015).
Krause, G. et al. Prospects of low trophic marine aquaculture to achieve food security in a zero-carbon world. Front. Sustain. Food Syst. https://doi.org/10.3389/fsufs.2022.875509 (2022).
Robinson, J. P. W., et al. (2022) Managing fisheries for maximum nutrient yield. Fish. Fish. https://doi.org/10.3389/fsufs.2022.875509 (2022).
Hospido, A. & Tyedmers, P. Life cycle environmental impacts of Spanish tuna fisheries. Fish. Res. 76, 174–186 (2005).
Winther, U., Skontorp Hognes, E., Jafarzadeh, S., Ziegler, F. Greenhouse Gas Emissions of Norwegian Seafood Products in 2017 (SINTEF, 2020).
Ziegler, F. & Hornborg, S. Stock size matters more than vessel size: the fuel efficiency of Swedish demersal trawl fisheries 2002–2010. Mar. Pol. 2014, 72–81 (2014).
Philis, G. et al. Comparing Life Cycle Assessment (LCA) of salmonid aquaculture production systems: status and perspectives. Sustainability 11, 2517 (2019).
Food and Agricultural Organization & World Health Organization of the United Nations. Guidelines for Nutrition labelling. Codex Alimentarius, International Food Standards. (FAO, 2017).
Food and Agricultural Organization of the United Nations. Fats and Fatty Acids in Human Consumption. Report of an Expert Consultation Group. (FAO, 2010).
Parker, R. W. R. Review of Life Cycle Assessment Research on Products Derived from Fisheries and Aquaculture. (Sea Fish Industry Authority, 2012).
Jonell, M. & Henriksson, P. Mangrove-shrimp farms in Vietnam—comparing organic and conventional systems using life cycle assessment. Aquaculture 447, 66–75 (2015).
Tyedmers, P. H., Watson, R. & Pauly, P. Fueling global fishing fleets. Ambio 34, 619–622 (2005).
Cashion, T. et al. Reconstructing global marine fishing gear use: catches and landed values by gear type and sector. Fish. Res. 206, 57–64 (2018).
Food and Agricultural Organization of the United Nations. Yield and Nutritional Value of the Commercially More Important Species. (FAO, 1989).
Acknowledgements
This research was funded by the Swedish Research Council Formas (Grant 2017-00842).
Author information
Authors and Affiliations
Contributions
M.B.: conceptualization, investigation, writing—original draft, writing—editing. E.H.: writing—editing. R.W.R.P.: conceptualization, investigation, visualization, writing—editing. K.M.: investigation, writing—editing. P.T.: conceptualization, writing—editing. F.Z.: conceptualization, investigation, writing–original draft, writing—editing, project administration, funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Heike Langenberg and Clare Davis. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bianchi, M., Hallström, E., Parker, R.W.R. et al. Assessing seafood nutritional diversity together with climate impacts informs more comprehensive dietary advice. Commun Earth Environ 3, 188 (2022). https://doi.org/10.1038/s43247-022-00516-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-022-00516-4
This article is cited by
-
Offshore aquaculture greenhouse gas emissions based on ocean net primary productivity
Nature Food (2024)
-
A traits-based approach to assess aquaculture’s contributions to food, climate change, and biodiversity goals
npj Ocean Sustainability (2024)
-
Nutrient yields from global capture fisheries could be sustainably doubled through improved utilization and management
Communications Earth & Environment (2023)
-
Extending the product-group-specific approach in nutritional life cycle assessment
The International Journal of Life Cycle Assessment (2023)
-
Eat more fish: when switching to seafood helps — and when it doesn’t
Nature (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
