
Abstract
Amplification of MYCN is the driving oncogenic change in a subset of high-risk neuroblastomas. The MYCN protein and the Aurora-A kinase form a complex during the S phase that stabilizes MYCN. Here we show that MYCN activates Aurora-A on chromatin, which phosphorylates histone H3 at serine 10 in the S phase, promotes the deposition of histone H3.3 and suppresses R-loop formation. Inhibition of Aurora-A induces transcription–replication conflicts and activates ataxia telangiectasia and Rad3-related (ATR) kinase, which limits double-strand break accumulation upon Aurora-A inhibition. Combined inhibition of Aurora-A and ATR kinases induces rampant tumor-specific apoptosis and tumor regression in mouse models of neuroblastoma, leading to permanent eradication in a subset of mice. The therapeutic efficacy is due to both tumor cell-intrinsic and immune cell-mediated mechanisms. We propose that targeting the ability of Aurora-A to resolve transcription–replication conflicts is an effective therapy for MYCN-driven neuroblastoma.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
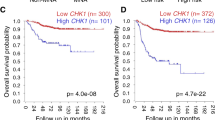

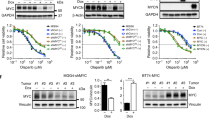
Data availability
ChiP-seq and RNA-seq data are available from the Gene Expression Omnibus under accession number GSE144288. Previously published sequencing data that we re-analyzed here are available under accession codes GSM1838910 and GSM1838911 (ref. 74). Source data are provided with this paper. All other data supporting the findings of the study are available from the corresponding authors upon request.
References
Dang, C. V. MYC on the path to cancer. Cell 149, 22–35 (2012).
Soucek, L. et al. Modelling Myc inhibition as a cancer therapy. Nature 455, 679–683 (2008).
Rickman, D. S., Schulte, J. H. & Eilers, M. The expanding world of N-MYC-driven tumors. Cancer Discov. 8, 150–163 (2018).
Walz, S. et al. Activation and repression by oncogenic Myc shape tumour-specific gene expression profiles. Nature 511, 483–487 (2014).
Sabo, A. et al. Selective transcriptional regulation by Myc in cellular growth control and lymphomagenesis. Nature 511, 488–492 (2014).
Nie, Z. et al. c-Myc is a universal amplifier of expressed genes in lymphocytes and embryonic stem cells. Cell 151, 68–79 (2012).
Lin, C. Y. et al. Transcriptional amplification in tumor cells with elevated c-Myc. Cell 151, 56–67 (2012).
Herold, S. et al. Recruitment of BRCA1 limits MYCN-driven accumulation of stalled RNA polymerase. Nature 567, 545–549 (2019).
Zeid, R. et al. Enhancer invasion shapes MYCN-dependent transcriptional amplification in neuroblastoma. Nat. Genet. 50, 515–523 (2018).
Baluapuri, A., Wolf, E. & Eilers, M. Target gene-independent functions of MYC oncoproteins. Nat. Rev. 21, 255–267 (2020).
Otto, T. et al. Stabilization of N-Myc is a critical function of Aurora A in human neuroblastoma. Cancer Cell 15, 67–78 (2009).
Brockmann, M. et al. Small molecule inhibitors of Aurora-A induce proteasomal degradation of N-myc in childhood neuroblastoma. Cancer Cell 24, 75–89 (2013).
Gustafson, W. C. et al. Drugging MYCN through an allosteric transition in Aurora kinase A. Cancer Cell 26, 414–427 (2014).
Richards, M. W. et al. Structural basis of N-Myc binding by Aurora-A and its destabilization by kinase inhibitors. Proc. Natl Acad. Sci. USA 113, 13726–13731 (2016).
Buchel, G. et al. Association with Aurora-A controls N-MYC-dependent promoter escape and pause release of RNA polymerase II during the cell cycle. Cell Rep. 21, 3483–3497 (2017).
Kufer, T. A. et al. Human TPX2 is required for targeting Aurora-A kinase to the spindle. J. Cell Biol. 158, 617–623 (2002).
Wang, H. et al. Improved low molecular weight Myc-Max inhibitors. Mol. Cancer Ther. 6, 2399–2408 (2007).
Zirath, H. et al. MYC inhibition induces metabolic changes leading to accumulation of lipid droplets in tumor cells. Proc. Natl Acad. Sci. USA 110, 10258–10263 (2013).
Mathsyaraja, H. et al. Max deletion destabilizes MYC protein and abrogates Eµ-Myc lymphomagenesis. Genes Dev. 33, 1252–1264 (2019).
Zeng, K., Bastos, R. N., Barr, F. A. & Gruneberg, U. Protein phosphatase 6 regulates mitotic spindle formation by controlling the T-loop phosphorylation state of Aurora A bound to its activator TPX2. J. Cell Biol. 191, 1315–1332 (2010).
Kim, S. R., Kim, K. B., Chae, Y. C., Park, J. W. & Seo, S. B. H3S10 phosphorylation-mediated transcriptional regulation by Aurora kinase A. Biochem. Biophys. Res. Commun. 469, 22–28 (2016).
Klaeger, S. et al. The target landscape of clinical kinase drugs. Science https://doi.org/10.1126/science.aan4368 (2017).
Manfredi, M. G. et al. Characterization of alisertib (MLN8237), an investigational small-molecule inhibitor of Aurora A kinase using novel in vivo pharmacodynamic assays. Clin. Cancer Res. 17, 7614–7624 (2011).
Shimomura, T. et al. MK-5108, a highly selective Aurora-A kinase inhibitor, shows antitumor activity alone and in combination with docetaxel. Mol. Cancer Ther. 9, 157–166 (2010).
Shannon, K. B. & Salmon, E. D. Chromosome dynamics: new light on Aurora B kinase function. Curr. Biol. 12, R458–R460 (2002).
Wilkinson, R. W. et al. AZD1152, a selective inhibitor of Aurora B kinase, inhibits human tumor xenograft growth by inducing apoptosis. Clin. Cancer Res. 13, 3682–3688 (2007).
Wang, F. et al. Histone H3 Thr-3 phosphorylation by haspin positions Aurora B at centromeres in mitosis. Science 330, 231–235 (2010).
Zeineldin, M. et al. MYCN amplification and ATRX mutations are incompatible in neuroblastoma. Nat. Commun. 11, 913 (2020).
Nakatani, Y., Ray-Gallet, D., Quivy, J. P., Tagami, H. & Almouzni, G. Two distinct nucleosome assembly pathways: dependent or independent of DNA synthesis promoted by histone H3.1 and H3.3 complexes. Cold Spring Harb. Symp. Quant. Biol. 69, 273–280 (2004).
Ivaldi, M. S., Karam, C. S. & Corces, V. G. Phosphorylation of histone H3 at Ser10 facilitates RNA polymerase II release from promoter-proximal pausing in Drosophila. Genes Dev. 21, 2818–2831 (2007).
Moore, M. J., Wang, Q., Kennedy, C. J. & Silver, P. A. An alternative splicing network links cell-cycle control to apoptosis. Cell 142, 625–636 (2010).
Ho, Y. S. et al. Griseofulvin potentiates antitumorigenesis effects of nocodazole through induction of apoptosis and G2/M cell cycle arrest in human colorectal cancer cells. Int. J. Cancer 91, 393–401 (2001).
Girard, C. et al. Post-transcriptional spliceosomes are retained in nuclear speckles until splicing completion. Nat. Commun. 3, 994 (2012).
Castellano-Pozo, M. et al. R loops are linked to histone H3 S10 phosphorylation and chromatin condensation. Mol. Cell 52, 583–590 (2013).
Hamperl, S., Bocek, M. J., Saldivar, J. C., Swigut, T. & Cimprich, K. A. Transcription-replication conflict orientation modulates R-loop levels and activates distinct DNA damage responses. Cell 170, 774–786.e19 (2017).
Cimprich, K. A. & Cortez, D. ATR: an essential regulator of genome integrity. Nat. Rev. 9, 616–627 (2008).
Shiotani, B. et al. Two distinct modes of ATR activation orchestrated by Rad17 and Nbs1. Cell Rep. 3, 1651–1662 (2013).
Foote, K. M. et al. Discovery and characterization of AZD6738, a potent inhibitor of ataxia telangiectasia mutated and rad3 related (ATR) kinase with application as an anticancer agent. J. Med. Chem. 61, 9889–9907 (2018).
Lecona, E. & Fernandez-Capetillo, O. Targeting ATR in cancer. Nat. Rev. Cancer 18, 586–595 (2018).
Maya-Mendoza, A. et al. High speed of fork progression induces DNA replication stress and genomic instability. Nature 559, 279–284 (2018).
Hamperl, S. & Cimprich, K. A. Conflict resolution in the genome: how transcription and replication make it work. Cell 167, 1455–1467 (2016).
Sollier, J. et al. Transcription-coupled nucleotide excision repair factors promote R-loop-induced genome instability. Mol. Cell 56, 777–785 (2014).
Sloane, D. A. et al. Drug-resistant Aurora A mutants for cellular target validation of the small molecule kinase inhibitors MLN8054 and MLN8237. ACS Chem. Biol. 5, 563–576 (2010).
Dodson, C. A. et al. Crystal structure of an Aurora-A mutant that mimics Aurora-B bound to MLN8054: insights into selectivity and drug design. Biochem. J. 427, 19–28 (2010).
Tse, A. N. et al. CHIR-124, a novel potent inhibitor of Chk1, potentiates the cytotoxicity of topoisomerase I poisons in vitro and in vivo. Clin. Cancer Res. 13, 591–602 (2007).
Brannan, K. et al. mRNA decapping factors and the exonuclease Xrn2 function in widespread premature termination of RNA polymerase II transcription. Mol. Cell 46, 311–324 (2012).
Weiss, W. A., Aldape, K., Mohapatra, G., Feuerstein, B. G. & Bishop, J. M. Targeted expression of MYCN causes neuroblastoma in transgenic mice. EMBO J. 16, 2985–2995 (1997).
DuBois, S. G. et al. Phase II trial of alisertib in combination with irinotecan and temozolomide for patients with relapsed or refractory neuroblastoma. Clin. Cancer Res. 24, 6142–6149 (2018).
Beltran, H. et al. A phase II trial of the Aurora kinase A inhibitor alisertib for patients with castration-resistant and neuroendocrine prostate cancer: efficacy and biomarkers. Clin. Cancer Res. 25, 43–51 (2019).
Mosse, Y. P. et al. A phase II study of alisertib in children with recurrent/refractory solid tumors or leukemia: Children’s Oncology Group Phase I and Pilot Consortium (ADVL0921). Clin. Cancer Res. 25, 3229–3238 (2019).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Kortlever, R. M. et al. Myc cooperates with Ras by programming inflammation and immune suppression. Cell 171, 1301–1315.e14 (2017).
Barber, G. N. STING: infection, inflammation and cancer. Nat. Rev. Immunol. 15, 760–770 (2015).
Chiu, A. C. et al. Transcriptional pause sites delineate stable nucleosome-associated premature polyadenylation suppressed by U1 snRNP. Mol. Cell 69, 648–663.e7 (2018).
Guarino Almeida, E., Renaudin, X. & Venkitaraman, A. R. A kinase-independent function for AURORA-A in replisome assembly during DNA replication initiation. Nucleic Acids Res. 48, 7844–7855 (2020).
Merigliano, C., Mascolo, E., Cesta, A., Saggio, I. & Verni, F. A new role for Drosophila Aurora-A in maintaining chromosome integrity. Chromosoma 128, 41–52 (2019).
Debruyne, D. N. et al. BORIS promotes chromatin regulatory interactions in treatment-resistant cancer cells. Nature 572, 676–680 (2019).
Kotake, Y. et al. Splicing factor SF3b as a target of the antitumor natural product pladienolide. Nat. Chem. Biol. 3, 570–575 (2007).
Olson, C. M. et al. Pharmacological perturbation of CDK9 using selective CDK9 inhibition or degradation. Nat. Chem. Biol. 14, 163–170 (2018).
Chao, S. H. et al. Flavopiridol inhibits P-TEFb and blocks HIV-1 replication. J. Biol. Chem. 275, 28345–28348 (2000).
Michelena, J. et al. Analysis of PARP inhibitor toxicity by multidimensional fluorescence microscopy reveals mechanisms of sensitivity and resistance. Nat. Commun. 9, 2678 (2018).
Ginno, P. A., Lott, P. L., Christensen, H. C., Korf, I. & Chedin, F. R-loop formation is a distinctive characteristic of unmethylated human CpG island promoters. Mol. Cell 45, 814–825 (2012).
Boguslawski, S. J. et al. Characterization of monoclonal antibody to DNA·RNA and its application to immunodetection of hybrids. J. Immunol. Methods 89, 123–130 (1986).
Chen, X. et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell 133, 1106–1117 (2008).
Jaenicke, L. A. et al. Ubiquitin-dependent turnover of MYC antagonizes MYC/PAF1C complex accumulation to drive transcriptional elongation. Mol. Cell 61, 54–67 (2016).
Kopper, F. et al. Damage-induced DNA replication stalling relies on MAPK-activated protein kinase 2 activity. Proc. Natl Acad. Sci. USA 110, 16856–16861 (2013).
Henssen, A. G. et al. Therapeutic targeting of PGBD5-induced DNA repair dependency in pediatric solid tumors. Sci. Transl. Med. https://doi.org/10.1126/scitranslmed.aam9078 (2017).
Langmead, B., Trapnell, C., Pop, M. & Salzberg, S. L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, R25 (2009).
Li, H. et al. The sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Quinlan, A. R. BEDTools: the Swiss-Army tool for genome feature analysis. Curr. Protoc. Bioinformatics 47, 11.12.1–11.12.34 (2014).
Ramirez, F., Dundar, F., Diehl, S., Gruning, B. A. & Manke, T. deepTools: a flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 42, W187–W191 (2014).
Shen, L., Shao, N., Liu, X. & Nestler, E. ngs.plot: Quick mining and visualization of next-generation sequencing data by integrating genomic databases. BMC Genomics 15, 284 (2014).
Devaiah, B. N. et al. BRD4 is a histone acetyltransferase that evicts nucleosomes from chromatin. Nat. Struct. Mol. Biol. 23, 540–548 (2016).
Hao, Z. et al. RIdeogram: drawing SVG graphics to visualize and map genome-wide data on the idiograms. PeerJ Comput. Sci. 6, e251 (2020).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Liberzon, A. et al. Molecular signatures database (MSigDB) 3.0. Bioinformatics 27, 1739–1740 (2011).
Acknowledgements
This work was supported by grants from the European Research council (AuroMYC), German Cancer Aid (Enable), the Federal Ministry of Education and Research (DKTK), the German Research Foundation (via DFG Research Group 2314 to M.E.) and German Cancer Aid (via the Mildred Scheel Early Career Center to G.B.). Cancer Research UK supports the Cancer Imaging Centre at ICR in association with the MRC and Department of Health (England) (C1060/A16464) and a Children with Cancer UK Research Fellowship (to Y.J.). A.G.H. is supported by the German Research Foundation and Wilhelm Sander Stiftung and participates in the BIH-Charité Clinical Scientist Program. E.P. and L.C. received a Children with Cancer UK Project Grant (2014/174). L.C. received Cancer Research UK Program Grants (C34648/A18339 and C34648/A14610). R.B. received a Cancer Research UK Programme Award (C24461/A23302). We thank B. Bauer for technical help with the immunohistochemistry, A. Schlosser for proteomics analyses, W. Schmitz for mass spectrometry, B. Martins da Costa and J. Rolff for help with the animal experiments, S. Heinzlmeier and B. Küster for providing the graphics on Aurora-A inhibitors, and members of the Eilers laboratory for evaluating the immunohistochemical images.
Author information
Authors and Affiliations
Contributions
I.R. and G.B. performed most of the experiments. E.P. and Y.J. performed all of the in vivo experiments and MRI measurements in TH-MYCN mice. H.D.G. analyzed the in vivo experiments in PDX models. M.G. performed the high-content microscopy experiments. C.G. performed the fiber assays. C.S.-V. analyzed the immunofluorescence experiments. M.R. performed the in vitro kinase assays. P.G. analyzed the ChIP and RNA-seq data. M.R. evaluated the pathology. C.P.A. measured the high-throughput data. P.B. analyzed the proteomics data. G.B., E.P., A.G.H., M.A., M.D., R.B., A.E., J.A., L.C. and M.E. devised and supervised the experiments. G.B., L.C. and M.E. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Cancer thanks the anonymous reviewers for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Aurora-A phosphorylates H3S10 in S phase.
a, Propidium-Iodide stained FACS profiles of IMR-5 cells synchronized at the G1/S boundary by a double-thymidine block (T0). Asynchronous cells are shown as controls. Cells were released from the block for 4 h into S phase, for 8 h into G2/M phase and for 14 h into G1 phase Data representative of 3 independent experiments with similar results. b, Fractionation of synchronized MYCN-amplified IMR-5 neuroblastoma cells. (Left): Immunoblots of equal aliquots of each fraction were probed for the indicated proteins. Data representative of 3 independent experiments with similar results. (Right): Quantitation of Aurora-A levels in each cell cycle phase. Shown is mean ± S.D. (n = 3 independent experiments). c, MYCN ChIP at indicated loci in asynchronous IMR-5 cells treated for 4 h with 10058-F4 (100 µM). IgG control was used as control for antibody specificity. Data are presented as mean of technical triplicates. Data representative of 3 independent experiments with similar results. d, Immunoblots of indicated proteins of IMR-5 cells either asynchronous, synchronized by double thymidine block into S or G2 phase or synchronized into mitosis by incubation with nocodazole for 16 h. Data representative of 2 independent experiments with similar results. e, Radar blots documenting specificity of Aurora kinase inhibitors: MLN8237, MK5108, AZD1152; data are from22. f, Quantification of pH3S10 staining in IMR-5 cells treated for 8 h with MLN8237 (1 µM) relative to control (DMSO) cells; each grey dot represents a cell. In S and G2 phase number of spots and in mitotic cells intensity of pH3S10 signal relative to DMSO is shown. Data are presented as mean ± S.D. (n≥614 cells per condition examined over 3 independent experiments). g, Quantification of pH3S10 staining of IMR-5 cells in S phase treated for 8 h with 100 nM MLN8237, 1 µM MK5108 or DMSO as control. Each dot represents one cell. Data are presented as mean ± S.D. (n≥147 cells per condition examined over 3 independent experiments). h, Quantification of pH3T3 intensity in IMR-5 cells treated for 8 h with 100 nM MLN8237, 100 nM AZD1152 or DMSO as control; each grey dot represents a cell. Shown is the mean ± S.D. (n≥47 cells per condition examined over 3 independent experiments). i, Representative example pictures of 3 independent experiments with similar results. White line indicates 15 µm.
Extended Data Fig. 2 Effects of Aurora-A inhibition on H3.3 and pH3S10.
a, (Top): Ideogram illustrating the 50 bins with the strongest reduction in pH3S10 MLN8237 as compared to pH3S10 DMSO (blue rectangles). (Bottom): Levels of pH3S10 relative to total H3 along all chromosomes of S phase-synchronized IMR-5 cells treated for 4 h with 1 µM MLN8237 (red line) and DMSO (blue line) (n = 2 independent experiments). b, pH3S10 ChIP at indicated loci in S phase synchronized IMR-5 cells treated for 4 h with 1 µM MLN8237. IgG control was used as control for antibody specificity. Data are presented as mean of technical triplicates Data representative of 3 independent experiments with similar results c, Metagene plot of ChIP-Rx signal for histone H3.3 and pH3S10 in S phase-synchronized IMR-5 cells treated for 4 h with MLN8237 (1 µM) or DMSO. The graph is centered on the first nucleosome (‘+1 dyad’) downstream of the TSS for N = 14,340 genes. (n = 2 independent replicates). d, Browser tracks of MYCN, total RNAPII, H3.3 and pH3S10 of ChIP-Rx at a MYCN high (left) and low (right) bound locus. Nucleosome-free zone is indicated in grey. e, Metagene plot of ChIP-Rx signal for histone H3 and histone H3.3 in S phase-synchronized IMR-5 cells treated for 4 h with 1 µM MLN8237 or DMSO. The signal is centered on the first nucleosome (‘+1 dyad’) located downstream of the TSS for N = 14,340 genes (n = 3 independent experiments). f, Immunoblot of asynchronous SH-EP and SH-EP MYCN cells. Vinculin was used as loading control. Data representative of 3 independent experiments with similar results.
Extended Data Fig. 3 Aurora-A impacts RNAPII, splicing and R loop formation in S phase.
a, Metagene plot of all expressed genes (N = 17,533) illustrating distribution of ChIP-Rx signal of total RNAPII within transcribed regions in S phase-synchronized IMR-5 cells treated for 4 h with 1 µM MLN8237, 1 µM MK5108 or solvent control (DMSO) (n = 2 independent experiments). b, 2D Kernel density plot showing RNAPII traveling ratio in S phase-synchronized cells treated for 4 h with MLN8237 (1 µM) and in control (DMSO) cells (n = 2 independent experiments). c, Metagene plot of ChIP-Rx signal for RNAPII pSer2 in S phase-synchronized IMR-5 cells treated for 2 h with MLN8237 (1 µM), MK5108 (1 µM) or DMSO. The signal is centered on the first nucleosome (‘+1 dyad’) located within 300 nt downstream of the TSS for n = 14,340 genes (n = 2 independent replicates). d, Browser tracks of MYCN, total RNAPII and RNAPII pSer2 of ChIP-Rx at FASN gene. e, Immunoblots of IMR-5 cells that were synchronized for the indicated cell cycle phase and treated with Pladienolide B (PlaB;1 µM), MLN8237 (1 µM), MK5108 (1 µM) or DMSO (4 h). Actin was used as loading control. Data representative of 3 independent experiments with similar results. f, Definition of read categories; blue lines show where reads were mapped to; grey categories (exonic, spliced) reads represent mature mRNA, blue reads categories (exon-intron, intronic) represents non-spliced pre-mRNA, red categories (TSS, TES, TES-RT, intergenic) represents RNA without coding sequence. TSS: Transcription start site, TES: transcription end site. g, Diagram showing the setup of the 4sU-sequencing experiments in S phase-synchronized IMR-5 cells. h, Mean percentage of reads recovered in each category described in (f) (n = 3 independent experiments). All treatments (PlaB, MK, MLN) significantly reduce the percentage of spliced reads relative to the ‘DMSO’ control (p<1.0e-15 using paired two-tailed t test and Wilcoxon matched-pairs signed rank test). i, DRIP using S9.6 antibody after 8 h of 1 µM MLN8237 or 1 µM PlaB treatment. Incubation with RNaseH1 and IgG were used as controls. Shown is the mean of technical triplicates. Data representative of 2 independent experiments with similar results. j, (Left) DRIP using S9.6 antibody after 48 h of siAurora-A treatment. Shown is the mean of technical triplicates. (Right) Immunoblot of siRNA-transfected IMR-5 cells. Vinculin was used as loading control. Data representative of 2 independent experiments with similar results.
Extended Data Fig. 4 Characterization of DNA replication and R-loop formation.
a, Quantification of fork progression of SH-EP and SH-EP MYCN cells pretreated for 3 h with 100 nM or 1 µM MLN8237, 1 µM of AZD6738 or a combination (100 nM MLN8237 and 1 µM AZD6738). P-values were calculated using unpaired two-tailed t-test comparing two conditions (n≥168 fibers per condition were examined over 2 independent experiments). b, Annexin-V/PI FACS of IMR-5 or IMR-5 cells with inducible Aurora-A WT and T217D. After pre-treatment for 24 h with doxycycline to induce Aurora-A WT and T217D, cells were treated for 48 h with MLN8237 (100 nM), AZD6738 (1 µM) or both. Shown is the mean ± S.D. (n = 3 independent experiments). c, Annexin-V/PI FACS of IMR-5 treated for 48 h with MLN8237 (100 nM), CHIR-124 (1 µM) or both. Shown is the mean ± S.D. (n = 3 independent experiments). d, DNA-RNA-Immunoprecipitation (DRIP) using S9.6 antibody of IMR-5 cells expressing a doxycycline inducible HA-RNaseH1. After 24 h doxycycline treatment, cells were treated for 8 h with 1 µM MLN8237. Incubation with RNaseH1 and IgG were used as controls for non-specific chromatin binding. Shown is the mean of technical triplicates. Data representative of 2 independent experiments with similar results. Insert shows expression of HA-RNaseH1 in asynchronous IMR-5 cells upon doxycycline treatment for 24 h. Vinculin (VCL) was used as loading control. Data representative of 3 independent experiments with similar results. e, Metagene plot of ChIP-Rx signal for total RNAPII and H3.3 in S phase-synchronized RNaseH1-IMR-5 cells treated for 24 h with doxycycline to induce RNaseH1 expression or EtOH (as control). Data show mean for N = 3,731 genes. The signal is centered on the first nucleosome (‘+1 dyad’) located within 300 nt downstream of the TSS (n = 2 independent experiments). f, Browser tracks of MYCN and total RNAPII of ChIP-Rx in (e) at FASN and NCL gene. g, Metagene plot of ChIP-Rx signal for RNAPII pSer2 in S phase-synchronized RNaseH1-IMR-5 cells treated for 24 h with doxycycline (to induce RNaseH1 expression) or EtOH (as control). Data shown for n = 14,340 genes. The signal is centered on the first nucleosome (‘+1 dyad’) located within 300 nt downstream of the TSS for 14,340 genes (n = 2 independent experiments). h, Quantification of PLA signals between RNAPII and PCNA in asynchronous RNaseH1-IMR-5 cells treated for 24 h with doxycycline to induce RNaseH1 expression. Each dot represents mean PLA signal of all cells in one well compared to solvent control. Shown is the mean ± S.D.. P-value was calculated using unpaired two-tailed t-test relative to DMSO control (n = 3 independent experiments). i, Boxplot showing intensity of pKAP1 staining in mitotic cells upon the treatment with the indicated drugs (8 h) and induction of RNaseH1 expression for 24 h. P-value was calculated using two-tailed t-test between EtOH and Dox of each condition (n≥50 cells per condition were examined over 2 independent experiment).
Extended Data Fig. 5 Treatment is not toxic and tumor specific.
a, Boxplot showing the concentration of indicated inhibitors in the tumor tissue of mice treated for 24 h or 5 days. N = 4 mice (control, 24 h treatment), 5 mice (5 days treatment). b, Relative changes in body weight of mice treated with MLN8237 and AZD6738 compared to mice treated with vehicle control. Shown is the mean ± S.D. N = 42 mice (control), 43 mice (treated). c, Staining of tumor tissue with S9.6 incubated with different RNases to document specificity of S9.6. staining for R-loops (n = 1 section for each experimental condition). d, Histology of proliferative gut tissue in untreated (top) and treated (bottom) mice showing H&E, Ki-67 and cleaved caspase 3 staining. N = 2 mice (control), 3 mice treated). e, Representative MRI as well as sections of a tumor treated with a combination of MLN8237 and olaparib (N = 3 animals were evaluated). f, MRI sections of the two long term-surviving mice at day 0 and after 7 days of treatment with the combination of AZD6738 (30 mg/kg) and MLN8237 (15 mg/kg). Dashed white lines indicate tumor circumference. g, P-values calculated using Mantel-Cox log-rank test comparing the survival of different groups shown in Figure 6c.
Extended Data Fig. 6 Therapeutic efficacy in Patient-derived xenograft models.
a, Histology of representative tumor sections of MYCN-amplified PDX models treated as indicated. AZD6738 was administered at 50 mg/kg every day and MLN8237 at 7.5 mg/kg on a 5 days on, 2 days off schedule. Shown are stainings from tumors recovered after treatment for 14 days. Each group comprises three animals. b, Box plot showing quantification of R-loop-, γH2AX- and cleaved caspase 3-positive cells in tumor sections. P-values were calculated using unpaired two-tailed t-test using Welch’s correction (n≥9 sections from animals described in panel (a) were evaluated). c, Relative changes in tumor volume of four MYCN non-amplified PDX models upon treatment with the indicated inhibitors. Shown is the mean ± S.E.M. N indicates the animal number for each experimental cohort. P-values comparing control and combination (indicated with a black dashed line) were calculated using unpaired two-sided t-test.
Extended Data Fig. 7 Aurora-A/ATR inhibition engages the immune system.
a, GSEA signatures showing response of a hallmark gene set indicating Interferon gamma response in the TH-MYCN (top) and a PDX (bottom) model. b, (Top): Representative pictures by flow cytometry showing cell gating. (Bottom): Populations of CD45+ immune cells in the tumor microenvironment after treatment of TH-MYCN mice with the combination of AZD6738 (25 mg/kg) and MLN8237. Shown is the mean ± S.E.M.. Significance was calculated using unpaired two-tailed t-test (N = 4 animals from each condition were evaluated). c, Histology of representative tumor sections showing NKp46-positive cells and pSTAT1 in tumors of TH-MYCN mice treated with combined Aurora-A/ATR inhibition. N = 4 mice (control, 24 h treatment), 5 mice (5 days treatment). d, Relative changes in tumor volume of subcutaneous xenografts in nude mice after treatment with vehicle or the combination of MLN8237 and AZD6738 (25 mg/kg). Shown is the mean ± S.E.M. (N = 5 animals per group). e, Bar graph showing the survival of allograft mice treated with MLN8237 and AZD6738. Shown is the mean ± S.E.M. (N = 4 animals per group).
Supplementary information
Source data
Source Data Fig. 1
Unprocessed western blots for Fig. 1a.
Source Data Fig. 1
Statistical source data for Fig. 1a–c,f.
Source Data Fig. 2
Statistical source data for Fig. 2d–f.
Source Data Fig. 3
Unprocessed western blots for Fig. 3b.
Source Data Fig. 3
Statistical source data for Fig. 3a,c,e,f.
Source Data Fig. 4
Statistical source data for Fig. 4a,c,d.
Source Data Fig. 5
Statistical source data for Fig. 5b,d.
Source Data Fig. 6
Statistical source data for Fig. 6b,c.
Source Data Fig. 7
Statistical source data for Fig 7a,c,d.
Source Data Extended Data Fig. 1
Unprocessed western blots for Extended Data Fig. 1b,d.
Source Data Extended Data Fig. 1
Statistical source data for Extended Data Fig. 1b,c,f–h.
Source Data Extended Data Fig. 2
Unprocessed western blots for Extended Data Fig. 2f.
Source Data Extended Data Fig. 2
Statistical source data for Extended Data Fig. 2b.
Source Data Extended Data Fig. 3
Unprocessed western blots for Extended Data Fig. 3e,j.
Source Data Extended Data Fig. 3
Statistical source data for Extended Data Fig. 3i,j.
Source Data Extended Data Fig. 4
Unprocessed western blots for Extended Data Fig. 4d.
Source Data Extended Data Fig. 4
Statistical source data for Extended Data Fig. 4a–d,h,i.
Source Data Extended Data Fig. 5
Statistical source data for Extended Data Fig. 5a,b.
Source Data Extended Data Fig. 6
Statistical source data for Extended Data Fig. 6b,c.
Source Data Extended Data Fig. 7
Statistical source data for Extended Data Fig. 7b,d,e.
Rights and permissions
About this article

Cite this article
Roeschert, I., Poon, E., Henssen, A.G. et al. Combined inhibition of Aurora-A and ATR kinases results in regression of MYCN-amplified neuroblastoma. Nat Cancer 2, 312–326 (2021). https://doi.org/10.1038/s43018-020-00171-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s43018-020-00171-8
This article is cited by
-
Chromatin modifiers in human disease: from functional roles to regulatory mechanisms
Molecular Biomedicine (2024)
-
Targeting ATR in patients with cancer
Nature Reviews Clinical Oncology (2024)
-
PAF1c links S-phase progression to immune evasion and MYC function in pancreatic carcinoma
Nature Communications (2024)
-
Protein kinase D2-Aurora kinase A-ERK1/2 signalling axis drives neuroendocrine differentiation of epithelial ovarian cancer
Molecular and Cellular Biochemistry (2024)
-
The chromatin network helps prevent cancer-associated mutagenesis at transcription-replication conflicts
Nature Communications (2023)
