
Abstract
The theories for substrate recognition in enzyme catalysis have evolved from lock-key to induced fit, then conformational selection, and conformational selection followed by induced fit. However, the prevalence and consensus of these theories require further examination. Here we use cryogenic electron microscopy and African swine fever virus type 2 topoisomerase (AsfvTop2) to demonstrate substrate binding theories in a joint and ordered manner: catalytic selection by the enzyme, conformational selection by the substrates, then induced fit. The apo-AsfvTop2 pre-exists in six conformers that comply with the two-gate mechanism directing DNA passage and release in the Top2 catalytic cycle. The structures of AsfvTop2-DNA-inhibitor complexes show that substantial induced-fit changes occur locally from the closed apo-conformer that however is too far-fetched for the open apo-conformer. Furthermore, the ATPase domain of AsfvTop2 in the MgAMP-PNP-bound crystal structures coexist in reduced and oxidized forms involving a disulfide bond, which can regulate the AsfvTop2 function.
Similar content being viewed by others
Introduction
The past half century has seen major advancements in the theories for substrate recognition in enzyme catalysis from lock-key1 to induced fit2, then conformational selection3,4,5, and recently conformational selection followed by induced fit4,6,7. Furthermore, the apo-enzyme (ligand-free) was suggested to pre-exist as a subset of catalytically relevant conformers (termed catalytic selection here) for facilitating the conformational selection7,8,9. The concept of conformational selection started from the early report of multiple conformational states of myoglobin10 to the recent emphasis of conformational landscapes and conformational selection4,6,7,11,12,13,14,15,16. Of particular interest is the finding by Kern and coworkers that, even in the absence of ligands, adenylate kinase (Adk) in crystals pre-exists in three conformers with varying degrees of lid openness along the catalytic trajectory, suggesting that the structure of the apo enzyme has evolved to adopt conformations ready to bind its substrates and reaction intermediates8, a property termed catalytic selection herein. A recent study showed that this property is coupled with the conformational selection and induced fit mechanisms upon ligand binding, based on an advanced nuclear magnetic resonance (NMR) relaxation approach that enabled the structure determination of Adk at high-energy and minor populated states7. Related properties were also found in several other proteins17, though Adk has been most extensively investigated. In fact, the catalytic selection mechanism could be widespread, but only one structure is solved a time. For example, in the apo state of the four members of the X-family DNA polymerases, Pol β exists in an open form18 while Pol λ19, Pol μ20 and TdT21 pre-exist in the closed form.
On the other hand, the relationship between protein dynamics and functions was often examined for relatively small monomeric proteins by NMR primarily, as in the signaling protein NtrC22, dihydrofolate reductase (DHFR)11 and tyrosine phosphatases12. The Adk study used X-ray crystallography, NMR, and MD simulation7,8,23, which well encapsulated the subtle yet significant local differences between the conformational sub-states. However, for these theories to be broadly applicable, they need to be demonstrated for large and multimeric enzymes that catalyze complicated reactions. The enzyme used in this study is a type II DNA topoisomerase (Top2) from African swine fever virus (Asfv), a dimeric multi-subdomain enzyme (271 kDa) that catalyzes multistep reactions to modulate DNA topology (Fig. 1).

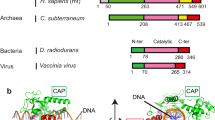
a Schematic illustration for the subdomains of AsfvTop2, Top2 and gyrase. b Cartoon model illustration of the proposed Top2 conformational states (numbered in black circles)40,41,44,47,48,49,50,51,58,59,64 with corresponding subdomains colored as indicated in the scheme above. The cycle starts with the apo-Top2 existing in an open dimeric form (state 1), which closes upon Gate (G) DNA binding to the DNA cleavage core (state 2). Next, the N-terminal ATPase (N-gate) is closed in the presence of ATP to capture the Transport (T) DNA segment (state 3). Then a double-strand G-DNA break occurs transiently to enable the T-DNA passage through the open DNA-gate (state 4), with one ATP molecule concurrently hydrolyzed. Next, re-ligation of the cleaved G-DNA segment triggers opening of the C-gate, leading to the release of T-DNA (state 5). Finally, the enzyme returns to the state 2 by closing the C-gate and releasing ADP. These conformational states or their variants are often referred to as the open form (state 1) or closed form (states 2–3) based on the status of the DNA-gate, while states 4 and 5 are respectively described as DNA-gate open and C-gate open forms here. The PDB codes for the previously reported structures and those from this study (Table 1) are listed, with the latter underlined.
It is now common to identify multiple structures from a single dataset in cryogenic-electron microscopy (cryo-EM)24,25. A vast majority of such conformers involve different ligand binding states or reaction intermediates. Only limited examples were found to address conformational variations in an apo enzyme26,27,28,29,30. In this work, we use cryo-EM to evaluate the current catalytic theories with AsfvTop2 and examine its induced fit conformational changes upon binding of ligands. The results allow us to propose a three-stage mechanism that encapsulates the substrate binding theories in a joint and order fashion. Furthermore, unique structural features and drug binding pocket of AsfvTop2 are unraveled for future antiviral innovation. The characterization of the disulfide bond formation in AsfvTop2 ATPase domain addresses a redox regulatory mechanism unique to topoisomerases.
Results
AsfvTop2 with complex gating mechanisms
Asfv is the only member of the Asfarviridae family, which belongs to the genetically and structurally complex eukaryotic large DNA viruses31. Albeit it poses a threat to worldwide agriculture with very high socioeconomic impact32,33, there is currently no effective vaccine or therapeutics. AsfvTop2 has been found to be involved in the early stage of viral gene replication34,35,36,37,38, and also detected at the intermediate and late stages of infection34. Although AsfvTop2 has low sequence identity to its eukaryotic and bacterial counterparts38,39, it harbors the general Top2 domain composition (Fig. 1a).
The domain composition of AsfvTop2 resembles that of the canonical Type IIA DNA topoisomerases (Top2), including the well-studied eukaryotic counterparts40,41,42, which form homodimers (Fig. 1a). Some other Top2 members, such as Topo IV and gyrase, are divided into two subunits and form heterotetramers43,44. Top2s change DNA topology by employing a complex catalytic cycle. Figure 1b outlines the major conformational states (1–5) of the ATP-fueled catalytic process, involving the proposed mechanisms based on a wealth of biochemical and structural studies for the truncated forms or subunits of Top2 from different species40,41,45,46,47,48,49,50,51,52. The catalytic process not only involves well-coordinated changes in protein quaternary structures to allow timely opening and closing of the N-, DNA-, and C-gates, but also ATP binding and sequential hydrolysis that relate the dimerization and catalytic activity of the ATPase domain to the mechanistic steps of the cycle49,51,53,54,55,56,57,58,59. However, structural and regulatory studies concerning how AsfvTop2 functions in the catalytic cycle and its drug binding modes have never been reported. Thus, it is an ideal system to test the recent theories of substrate recognition in catalysis, as well as to examine the catalytic mechanism in comparison with other Top2 enzymes. The structures we have obtained and their main properties are summarized in Table 1. Detailed structural data are listed in Tables 2–4. The detailed flow charts for solving the cryo-EM structures are shown in Supplementary Figs. 1–4.
Cryo-EM structures of apo-AsfvTop2
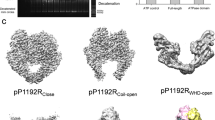
We first performed the cryo-EM reconstruction of the full-length apo-AsfvTop2 without DNA or nucleotide. As shown in the flowchart of data processing (Supplementary Fig. 1), one single dataset produced three globally distinct conformational states I, II, and III, instead of two states reported recently60 (Fig. 2a and Table 2). These three conformational states resemble the open (1), closed (2-3), and C-gate open (5) conformational states, respectively, in Fig. 1b. The initial 3D volumes of apo-AsfvTop2 conformers II and III are contributed by more than half of the total selected particle population (Fig. 2a and Supplementary Fig. 1). Furthermore, these primary conformational states are each sub-classified into two conformers, a and b, exhibiting local differences. The superimposed pairs show subtle global conformational changes and dynamic motions (Supplementary Movie 1: 3D variability of apo-AsfvTop2). These six structures are resolved at resolutions of 2.31–3.49 Å, and their structural models are shown in superposed pairs in Fig. 2b with the variations between a and b conformers highlighted.

a Cryo-EM density maps for the six conformers of the full-length AsfvTop2. The selected particle percentages of conformer I, II and III are labeled. The densities for the subdomains were clearly defined at a resolution of 2.31–3.49 Å, except for the ATPase domain. Sparse densities for ATPase of conformer IIb can be seen, which indicates that these domains were not cleaved in the process of obtaining structures, but were flexible instead. The TOPRIM subdomain is not modeled in the conformer I. b High resolution cryo-EM reconstructions of the apo-AsfvTop2 cleavage core domain for the three states (I-III), with the two sub-conformers of each state superposed with each other. The distance between the Cα atoms of the Y744/Y’744 from the two α-helices 13 on the WHD subdomains of the two subunits is shown for Ia (Ia: 30.3 Å; Ib: 27.6 Å). The detailed differences between sub-conformers are shown in insets. Annotation of the helices is based on the structure-based alignment (Supplementary Figs. 5, 6). c Upper panel: Superposed structures showing major conformational changes between conformers Ia and IIa (left); Ia and IIIa (center), and IIa and IIIa (right). The TOPRIM subdomain is omitted for clarity. Lower panel: The hinge connecting helix α27 and α28 contains two non-conserved proline residues. The residues and contacts that contribute to the structural differences are labeled. d Superposition of Ia, Ib, IIa, and IIIa with the comparable crystal structures from the yeast Top2 cleavage core: pdb 1bgw (apo, TOPRIM omitted)40, 1bjt (apo, TOPRIM omitted)46, 3l4k (crossed-linked with dsDNA, closed state)64, and 2rgr (complexed with dsDNA, closed state with C-gate open)48, respectively.
The distribution of the three primary co-existing apo-conformers is approximately 1:4:2 (Supplementary Fig. 1b). Even though such differences are modest energetically, they do suggest that the closed state II is the most stable state, followed by the C-gate open state III and then the open state I. These thermodynamic properties are reflected in their structural properties described in the following sections.
Catalytic selection of apo-AsfvTop2
Global conformational changes between different conformational states are illustrated by overlaying the conformers Ia/IIa, Ia/IIIa, and IIa/IIIa (Fig. 2c). The changes in the Ia/IIa pair are mainly in the closing of the DNA-gate via the WHD subdomain. The major difference between IIa and IIIa is hinge-like movements of the coiled-coils (α22 and α27) with approximately 13.6° and 18.3° tilts, respectively, governing the mobile states of the C-gate (Fig. 2c). The key determinants of the movements likely involve the flexible turn (residues 1158-1167) connecting the helices α27 and α28. Of which, two proline residues (P1163/P1165) provide certain degree of flexibility61 to this joint region, which was previously defined as an elbow46. Additionally, the indole ring side chain of W1170 from apo-IIIa turns clockwise with the nitrogen atom of its pyrrole ring shifted away from the hydroxyl group of Y1043. The interaction between the two aromatic residues is weakened in the apo-IIIa, therefore the helix α22 is less engaged with the helix α28 and attains flexibility to introduce the swing motion at the coiled-coil region. The continuum of structural conversion from conformers Ia to IIIb is shown in Supplementary Movie 2 (Structure conversion of apo-AsfvTop2 conformers), which not only highlights the inherent dynamic nature of apo-Top2s, but also illustrates the unexpected yet functionally relevant conformational changes reminiscent of the process for the unidirectional strand passage in the Top2 catalytic cycle (Fig. 1b)46,59,62,63.
To further provide the structural basis and functional relevance, we compared the conformers from the apo form of AsfvTop2 with individual structures of the cleavage core domain of yeast Top2, where each structure was solved separately under different conditions. As shown in Fig. 2d, conformers Ia/Ib were aligned globally with two apo-yeast Top2 structures45,46. Subsequently, the TOPRIM subdomain is posited downwards for constituting the DNA binding groove, as shown by aligning conformer IIa and the DNA bound structure of yeast Top2 (pdb 3l4k64, closed form). Furthermore, conformer IIIa resembles the C-gate open structure of yeast Top2 (pdb 2rgr48), one of the key conformations for DNA strand transport48,59. The parallel conformational changes between Asfv IIa-to-IIIa in the absence of DNA and yeast 3l4k64-to-2rgr48 in the presence of DNA provide strong support for the catalytic selection mechanism of AsfvTop2.
Conformational selection by ligands
In the conventional concept, the structural change between the apo form (yeast Top2 structure 1bgw45 or 1bjt46) and the DNA complex (2rgr48 or 3l4k64) can be attributed to induced fit, as has been addressed for 1bgw45 and 2rgr48 previously. However, it has not been addressed how DNA is able to bind a wide-open structure and induce large conformational changes, both globally and locally. Since the apo-state can pre-exist in multiple conformational states, the ligands are expected to select a conformer that is competent for ligand binding, according to the conformational selection theory3,4,5. Comparison with yeast Top2 structures suggested that the DNA complexes resemble apo-AsfvTop2 conformers II or III (apo-II or apo-III, respectively, Fig. 2d).
To obtain direct evidence for conformational selection, we performed structural analyses of AsfvTop2 complexes with DNA and an inhibitor. The sequences of DNA for the structural study are shown in Fig. 3a (Cut02a and Cut02b), which were chosen based on the DNA cleavage specificity of the active full-length AsfvTop2 (Supplementary Fig. 7) and described in Supplementary Note 1, Supplementary Fig. 8, and Supplementary Tables 1–3. Then we solved the cryo-EM structures of the full-length AsfvTop2 complexed with MgAMP-PNP (adenylyl-imidodiphosphate, a non-hydrolysable ATP analogue), DNA, and the Top2-targeting drug etoposide (a podophyllotoxin, with both Cut02a and Cut02b) (enzyme-DNA-inhibitor complexes EDI-1 and EDI-2, respectively) or m-AMSA (an aminoacridine, with Cut02a) (EDI-3) (2.70–3.0 Å, Tables 1 and 3) to dissect the specific binding of the DNA segments to the enzyme. Both etoposide and m-AMSA have been approved as anti-cancer drugs for human65, and have been shown to decrease the DNA decatenation activity of AsfvTop2 in vitro35.

a The DNA sequences of the two G-DNA segments (Cut02a and Cut02b). The cleavage sites are indicated with arrows. b Left: The representative global cryo-EM density of the full-length complexes, AsfvTop2 Cut02a/inhibitor (EDI-1, resolution 2.70 Å, contoured at 0.034 σ). Center: The global map fitted with the modeled cartoon structure, contoured at 0.198 σ. Right: Maps of bound DNA and inhibitor molecules. c Left: The EDI-1 complex is shown in cartoon cylinders and colored based on the color scheme in Fig. 1a. Center/right: The structure is slabbed through to see the two α-helices 13 clearly with their distance labeled. The DNA and inhibitor are colored in cyan with cartoon and sphere presentations, respectively. d Enlarged view of the overlaid ß-HP1 region (colored in cyan, residues 846–861). The conserved isoleucine in yeast (orange)/human (blue) Top2s64,66 and AsfvTop2 P852 (shown in stick) emanate towards DNA to different extents. e The intercalation of P852 and its distances (Å) to adjacent DNA bases are labeled. f Left: the spatial location of the ß-HP1, adjacent to ß-HP2 (residues 822-834) and ß-HP3 (residues 1012-1025), and the elbow region. Right: ß-HP1 is linked to the catalytic Y800 through ß-HP3. The structural moieties mentioned are highlighted in cyan. g, h Structural superimposition between AsfvTop2 and human Top2 showing the differences in drug binding in stereo views. The corresponding human Top2 residues (pdb 3qx3, etoposide bound66, pdb 4g0u, m-AMSA bound68) are shown in spheres and labeled in bold letters in parentheses.
The global structures of all three complexes resemble that of apo-II. As an example, the 3D maps of the complex with etoposide and Cut02a DNA (EDI-1) and those of the bound DNA and inhibitor are shown in Fig. 3b. The corresponding maps for EDI-2 and EDI-3 are shown in Supplementary Figs. 3 and 4, respectively. In all three structures, the DNA segment adopts a U-shaped conformation positioned on the DNA binding surface constituted of the TOPRIM, WHD, and Tower subdomains from both subunits (Fig. 3c). Importantly, despite the global similarity to apo-II by comparison Figs. 3c and Fig. 2b, there are substantial local changes as a result of induced fit. In the next two sections we first address the unique features of the AsfvTop2-DNA-inhibitor complexes relative to Top2s from other organisms, and then the induced fit conformational changes in the AsfvTop2.
Uniqueness of AsfvTop2-DNA-drug complex
Specific enzyme-DNA interactions in the three complexes are shown by schematic diagrams (Supplementary Fig. 9). In comparison with the corresponding diagram for eukaryotic Top248,66, the conserved DNA-intercalating residue (I872 in human or I833 in yeast Top2) is replaced by the P852 in AsfvTop2 (Supplementary Fig. 5b). The well-conserved isoleucine across eukaryotic and bacterial Top2s is involved in the DNA bending, and DNA-stimulated ATPase activity48,67. The alteration of the isoleucine to a proline by site-specific mutagenesis has been shown to dampen the E. coli Top2 (Top IV) functions in DNA bending and thus reduce ATPase activity47. Figure 3d shows that P852 and the conserved isoleucine from other Top2s residing at the ß-hairpin loop (annotated as ß-HP1 hereafter), protrude into DNA to different extents. The five-membered ring structure of P852 is intercalated between the DNA bases by an average distance of 4.0 Å (Fig. 3e). The ß-HP1 is buttressed by another two ß-HPs, and linked to the catalytic Y800 at the DNA-gate and helix α17 adjacent to the elbow (Fig. 3f). The potential functional relevance of this structural organization is elaborated in the following section. Additionally, AsfvTop2 lacks the essential conserved basic residue (K700 in yeast Top2) from the WHD subdomain (Supplementary Fig. 5b) for stabilizing the bent DNA conformation48. These distinct structural features likely differentiate the DNA modulation functions of AsfvTop2 from other Top2s.
Binding of the two Top2-targeting drugs etoposide and m-AMSA to AsfvTop2 is described in detail in Supplementary Note 2 and Supplementary Fig. 10. Structural comparison with human Top2-DNA-drug complexes66,68 highlights the distinctive residues from the two species facilitating the drug binding in different modes. For instances, AsfvTop2 lacks the conserved segment (PLR503GKXL) and polar residues that are characterized in its eukaryotic counterparts for the major interactions with the etoposide and m-AMSA, respectively (Fig. 3g, h). Based on the structural information, general guidelines for drug design targeting AsfvTop2 are summarized here: (1) a polycyclic aromatic core to assist drug intercalation into the DNA cleavage site as suggested for human Top2 targeting drug development68; (2) attaching a polar and bulky minor groove protruding moiety to the aromatic core that enhances the drug’s binding and specificity towards the minor groove-binding pocket of AsfvTop2.
Induced fit from apo-IIa to EDI complex
By overlaying the apo-conformer IIa and the DNA/inhibitor bound AsfvTop2 structure, Fig. 4a shows that the protein conformations of the two structures are very similar except that binding of DNA/inhibitor causes small subdomain reorientations to constitute the protein-DNA interface and re-organize the dimer interfaces at DNA- and C-gates (Fig. 4a and Supplementary Movie 3: DNA-induced subdomain reorientation of AsfvTop2), which can be considered the induced fit by ligand binding to the pre-selected conformer IIa. Presumably, conformer IIb should work equally well due to its similarity to IIa.

a The subdomain reorganization of AsfvTop2 is illustrated by aligning EDI-1 (gray) with apo-IIa (pink). b Half model of a, with a 90° rotation around the x-axis. The black rectangle highlights the extended loop that appears upon DNA binding. c The extended loop (residues 481-493, colored in green) shown inside the black square in panel b. The corresponding region in yeast/human Top264,66 is colored in orange/blue, respectively. d The close-up stereo view of the specific interactions between AsfvTop2 and DNA in the square box in (c). The interactions are governed by the K479 and K480 of AsfvTop2, which are replaced with Glu and Ala (residue numbers in parentheses), respectively, in yeast/human Top2. e Upper panels: A global view of the local movements upon DNA binding with focus on the ß-HP1 region (left) and the catalytic Y800 (right). Lower panels: The enlarged view of the squared regions in the upper panel. Left: The three ß-HP loops from apo-IIa concurrently shift downward in the EDI-1 complex. Middle: The hydrophobic network that coordinates the concurrent movements of the three ß-HPs and α22 in the EDI-1 complex. Right: Shifting of the catalytic Y800 toward DNA in the complex. f Stereo view for the spatial locations of the three ß-HPs and Y800 in apo-Ia (colored in green with italic label), apo-IIa (colored in pink with underlined label), and the EDI-1 complex (gray).
Furthermore, comparison of the DNA-bound complex with the apo-AsfvTop2 conformers unravels a flexible extended loop (from residue 481–493) protruding from the TOPRIM subdomain that becomes visible upon DNA binding (Fig. 4b). Sequence alignment (Supplementary Fig. 5b) indicates that this long-extended loop is unique for AsfvTop2. As it resides at the vicinity of the DNA binding groove, and located opposite to the ß-HP1 across the DNA (Fig. 4c), it could be involved in DNA modulation, analogous to the potential role of the human Top2 CTD (missing in AsfvTop2) in facilitating strand passage process69. Figure 4d highlights the differences in the protein-DNA specific interactions in that region between the viral and eukaryotic Top2-DNA complexes. The key positively charged residues involved in the DNA binding are well conserved amongst Asfv orthologues (Supplementary Fig. 11), which likely illustrates the conserved functionality of the DNA responsive loop in AsfvTop2.
Another induced fit feature involves re-positioning of ß-HP1 and subsequent movements of the neighboring structural elements that likely harbor functional relevance. Aligning EDI-1 and conformer IIa (Fig. 4e left panels) indicates that ß-HP1 shifts downward in EDI-1, resulting in relocation of P852 by 8.3 Å to accommodate DNA binding. Simultaneously, several hydrophobic contacts are formed between ß-HP1 and ß-HP2, shifting the ß-HP2 downward to form hydrophobic interactions with ß-HP3 and helix α22 (Fig. 4e lower panels left and center), part of the coiled-coil C-gate region. The ß-HP3 residing beneath the ß-HP2 moves downward by 5.5 Å. Since the N-terminal end of ß-HP3 is linked to the catalytic residue Y800 through a long β-strand linker and two small α-helices (Figs. 3f and 4e lower right), the spatial location of the Y800 in EDI-1 is shifted from apo-IIa by a distance of 7.3 Å and moves upward toward DNA. These DNA-responsive conformational changes occurring in the DNA binding groove are relayed to the active site and potentially the C-gate region, which likely collectively coordinate the dynamic gating operation.
Incompetence of apo-I for ligand binding
The results from the preceding section suggest that, even though the global conformation of apo-IIa is very similar to that of EDI-1, binding of DNA/inhibitor still induces small subdomain reorientations (Supplementary Movie 3: DNA-induced subdomain reorientation of AsfvTop2), yet coupled with substantial catalytically relevant conformational changes locally. On the other hand, as shown in Fig. 4f, the three ß-HPs and the catalytic Y800 in apo-Ia are farther away, relative to the corresponding moieties in apo-IIa, from the catalytically relevant positions in EDI-1, suggesting that it is less feasible for apo-I to bind the DNA substrate and undergo functionally relevant conformational changes. Taken together, our results suggest that it is advantageous for apo-II to coexist with apo-I through the catalytic selection process in order to be selected by the ligands, and to undergo further induced changes after ligand binding.
Functional selection by redox regulation
The ATPase domain regulates the Top2 catalytic cycle by binding ATP to induce closure of the N-gate, followed by hydrolyzing ATP to stimulate cleavage and passage of DNA40,41,51,53,55,56,58,70 (Fig. 1b), though the specific structural basis remains elusive. Here we report a different type of regulation by the ATPase domain of AsfvTop2. Using X-ray crystallography, we first found that the ATPase domain of AsfvTop2 seemed to coexist in two conformers in the MgAMP-PNP-bound crystal structure, and suspected that it could a mixture of reduced and oxidized forms. Then we resolved the crystal structure of AsfvTop2 ATPase domain complexed with MgAMP-PNP in the presence of the reducing agent β-mercaptoethanol (β-ME) (Fig. 5a, with density maps in Supplementary Fig. 12a). This structure showed unique features relative to the corresponding ATPase domain from E. coli and yeast Top2 (Supplementary Note 3 and Supplementary Fig. 13).

a, b The close-up view for the relative positions of MgAMP-PNP and the essential residues (C72, C138, H68, H73, and D137, all in stick presentation) involved in the potential regulatory mechanism. The reduced (a) and oxidized (b) forms are shown in cyan and pale cyan cartoon presentations, respectively. The disulfide bond between C72 and C138 is shown in brown stick. D137 and C138 are located at a long-extended surface loop (colored in blue) that harbors the active site residues interacting with the triphosphate group of the AMP-PNP. The segment E69-H73 that moves to form the disulfide bond is shown in red. c Analysis of ATP hydrolysis for the AsfvTop2 ATPase domain in the presence of 1 mM ATP and varying concentrations of the enzyme. The y-axis shows the specific activity from each measurement. Raw data are provided in Supplementary Data 5. d C72A mutation of the full-length AsfvTop2 resulted in significant reduction of DNA decatenation activity in comparison to the wild type over different time points. Detailed conditions are described in Supplementary Fig. 13c, d. Raw data are provided in Supplementary Data 6. Each data point in panels c, d includes the mean ± SE value from three independent reactions (n = 3) from the same batch of sample. Similar assay methods for ATPase and decatenation activities have been used previously for other Top2 proteins35,83. e A graphical summary illustrating that the enzyme-substrate recognition is mediated by the joint application of the substrate binding theories in order: (1) catalytic selection, (2) conformational selection, and (3) induced fit. The catalytic selection should be replaced by functional selection when referring to other functions instead of catalysis.
Next we address a new regulatory mechanism unique to AsfvTop2. The crystal structure of AsfvTop2 ATPase domain obtained from a non-reducing condition was solved at 1.14 Å resolution (Supplementary Note 3, with density maps in Supplementary Fig. 12). Structural analysis indicates that the structures from reducing and non-reducing conditions are without and with a C72-C138 disulfide bond, respectively (Fig. 5a, b). Interestingly, the disulfide bond formation involves a large movement of C72 to reach C138, along with relocation of H73 to pair with D137. Both residues and C138 are located at the solvent-exposed loops, in favor of the local motions. Additionally, this significant movement led to partial deformation of helix-α2 (Fig. 5b). The formation of disulfide bond between C72-C138 and the involvement of H73 are supported by mass spectrometry and NMR, respectively (Supplementary Fig. 14a, b).
Kinetic assays further indicated that the ATP hydrolysis activity was significantly dampened upon oxidation (Fig. 5c). Furthermore, under non-oxidizing condition, mutation of the residues involved in the disulfide bond formation, including H68A, C72A, H73A, and C138A, also led to different extent of inactivation, suggesting that these residues play additional roles in the non-oxidizing environment. H68 and C72 appeared to be crucial in holding adenosine moiety of AMP-PNP in position and thus both alanine mutations significantly attenuated the ATP hydrolysis activity. C138 resides at the long extended loop (residues F125-G147), encompassing the conserved ATP-lid that completely encloses the bound MgAMP-PNP at the active site (Fig. 5a, b, Supplementary Fig. 13b), therefore C138A mutation could affect the flexibility of the loop and subsequently the ATP hydrolysis. This effect appeared independent of the redox condition since the activity of C138A remained low in the oxidized form (Supplementary Fig. 14c). Thus, formation of the C72-C138 disulfide bond is most likely a naturally occurring way, in place of mutation, to enable the regulation of the ATPase activity and presumably the topoisomerase function. Importantly, this disulfide bond-mediated regulation of ATPase activity has not been shown in other Top2 enzymes. Furthermore, we showed that C72A significantly affected DNA decatenation activity of the full-length AsfvTop2 (Fig. 5d, Supplementary Fig. 14d, e), which demonstrates that the oxidation-mediated ATPase inactivation shown in Fig. 5c can regulate the primary function of the AsfvTop2.
These structural and functional analyses further support that the conformational states of the ATPase domain are coupled to the conformational and functional states of the cleavage core. Although the full-length structure of AsfvTop2 is not attainable due to conformational flexibility of the linker region that bridges two major functional domains, we have obtained a modeled structure based on the cryo-EM structure of the EDI-1 complex and the crystal structure of the ATPase domain, as described in Supplementary Note 4 and Supplementary Fig. 15.
Our result is in line with the redox-regulation of AsfvPolX71,72 and AsfvAP73, which have been shown to exist in both reduced and oxidized forms involving C81-C86 and C16-C20, respectively. The oxidized form of AsfvPolX has been shown to be specific to GG mismatch in addition to the four Watson-Crick base pairs74, whereas the reduced form has higher fidelity75. The structural basis of the low fidelity of the oxidized AsfvPolX has been elucidated by NMR76. AsfvPolX and AsfvAP, along with a low-fidelity Asfv DNA ligase77 are the key enzymes in the Asfv base excision repair system78,79. In line with the redox regulation of AsfvPolX and AsfvAP, formation of the disulfide bond by the cysteine pair (C72-C138) of AsfvTop2 can lower the activity of the topoisomerase, thus also save the ATP consumption and slow down DNA replication and cell growth, all of which may favor survival of the virus in under oxidative stress. Taken together, we propose that the redox mediated conformational transition of AsfvTop2 ATPase domain from reduced to oxidized state via the cysteine pair C72-C138 is a part of the functional selection, along with the redox regulations of AsfvPolX and AsfvAP, for the virus to adapt to the oxidizing environment of macrophage cytoplasm, analogous to catalytic selection in catalysis.
Conclusion
As a conclusion, we quote a statement by D. E. Koshland from his 1994 review entitled “The Key-Lock Theory and the Induced Fit Theory”80 with relevant terms from recent literature inserted in parentheses: “A new theory (conformational selection3,4,5, or conformational selection followed by induced fit4,6,7) must explain all the existing facts that pertain to it at the time of its enunciation. Gradually the new theory becomes accepted and then acquires anomalies due to the new facts uncovered after its enunciation. That in turn generates a newer theory (catalytic selection8,9) which elicits new techniques (cryo-EM) to test it and its predictions. These new techniques then uncover facts which eventually require further new theories and so on. The new theories are built on components of the old principles.” In this study, the newest theory is the combination of the old components in order: (1) catalytic selection by the enzyme, (2) conformational selection by the substrates, and induced fit together, as illustrated in Fig. 5e. For signaling or other function, the first step should be replaced by “functional selection by the protein”. We propose that such a three-stage mechanism is universal in enzyme catalysis or protein-ligand recognitions, and expect that it will be demonstrated in more examples in the future.
Methods
A brief summary is provided here. The functionally active full-length AsfvTop2 was over-expressed and purified from the yeast protein expression system (Supplementary Fig. 7) for cryo-EM structure determination of apo-AsfvTop2 and its DNA-inhibitor bound complexes (Supplementary Figs. 1–4). The functional studies performed included DNA decatenation, DNA relaxation, and cleavage assays (Fig. 5d, Supplementary Figs. 7, 8, 14). The active AsfvTop2 ATPase domain and its mutants were over-expressed and purified from E. coli for crystal structure determination (Supplementary Figs. 12, 13), redox studies, ATP hydrolysis activity assays (Figs. 5), 1D NMR, and mass spectrometry (Supplementary Fig. 14). The raw data for the functional and biophysical analyses are provided in Supplementary Data 1–6. A detailed explanation of all the procedures can be found in the Supplementary Methods section.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The coordinates and structure factors for AsfvTop2 ATPase domain in the reduced and oxidized states have been deposited in the Worldwide Protein Data Bank (wwPDB) under the accession codes: 8JA2 and 8JA1. Cryo-EM maps and the corresponding coordinate files for the apo full length AsfvTop2 have been deposited in the wwPDB under accession codes: 8J87, EMD-36062 (conformer Ia); 8J88, EMD-36063 (conformer Ib); 8J89, EMD-36064 (conformer IIa); 8J8A, EMD-36065 (conformer IIb); 8J8B, EMD-36066 (conformer IIIa); and 8J8C, EMD-36067 (conformer IIIb). Accession codes for the complexes are: 8J9V, EMD-36116 (Cut02aDNA/etoposide-bound, EDI-1); 8J9W, EMD-36117 (Cut02bDNA/etoposide-bound, EDI-2); and 8J9X, EMD-36118 (Cut02aDNA/m-AMSA-bound, EDI-3). The 3.68 Å cryo-EM map of full-length DNA/etoposide/AMP-PNP-bound AsfvTop2 can be accessed in the EMD-36473. The raw data for the functional and biophysical analyses are provided in Supplementary Data 1-6. The formal PDB validation reports of all deposited structures and maps are provided in Supplementary Data 7–18. The files of Supplementary Data 1, 3, 4, 6 have been stored in Figshare with https://doi.org/10.6084/m9.figshare.25123829.
References
Fischer, E. Einfluss der configuration auf die wirkung der enzyme. Ber. Dtsch. Chem. Ges. 27, 2985–1993 (1894).
Koshland, D. E. Application of a theory of enzyme specificity to protein synthesis. Proc. Natl Acad. Sci. USA 44, 98–104 (1958).
Monod, J., Wyman, J. & Changeux, J. P. On the nature of allosteric transitions: a plausible model. J. Mol. Biol. 12, 88–118 (1965).
Ma, B. & Nussinov, R. Enzyme dynamics point to stepwise conformational selection in catalysis. Curr. Opin. Chem. Biol. 14, 652–659 (2010).
Weikl, T. R. & Paul, F. Conformational selection in protein binding and function. Protein Sci. 23, 1508–1518 (2014).
Boehr, D. D., Nussinov, R. & Wright, P. E. The role of dynamic conformational ensembles in biomolecular recognition. Nat. Chem. Biol. 5, 789–796 (2009).
Stiller, J. B. et al. Structure determination of high-energy states in a dynamic protein ensemble. Nature 603, 528–535 (2022).
Henzler-Wildman, K. A. et al. Intrinsic motions along an enzymatic reaction trajectory. Nature 450, 838–844 (2007).
Henzler-Wildman, K. & Kern, D. Dynamic personalities of proteins. Nature 450, 964–972 (2007).
Austin, R. H., Beeson, K. W., Eisenstein, L., Frauenfelder, H. & Gunsalus, I. C. Dynamics of ligand binding to myoglobin. Biochemistry 14, 5355–5373 (1975).
Bhabha, G. et al. A dynamic knockout reveals that conformational fluctuations influence the chemical step of enzyme catalysis. Science 332, 234–238 (2011).
Whittier, S. K., Hengge, A. C. & Loria, J. P. Conformational motions regulate phosphoryl transfer in related protein tyrosine phosphatases. Science 341, 899–903 (2013).
Kern, D. From structure to mechanism: skiing the energy landscape. Nat. Methods 18, 435–436 (2021).
Nam, K. & Wolf-Watz, M. Protein dynamics: the future is bright and complicated! Struct. Dyn. 10, 014301 (2023).
Petrovic, D., Risso, V. A., Kamerlin, S. C. L. & Sanchez-Ruiz, J. M. Conformational dynamics and enzyme evolution. J. R. Soc. Interface 15, 20180330 (2018).
Ramanathan, A., Savol, A., Burger, V., Chennubhotla, C. S. & Agarwal, P. K. Protein conformational populations and functionally relevant substates. Acc. Chem. Res. 47, 149–156 (2014).
Eisenmesser, E. Z. et al. Intrinsic dynamics of an enzyme underlies catalysis. Nature 438, 117–121 (2005).
Sawaya, M. R., Pelletier, H., Kumar, A., Wilson, S. H. & Kraut, J. Crystal structure of rat DNA polymerase beta: evidence for a common polymerase mechanism. Science 264, 1930–1935 (1994).
Liu, M. S. et al. Structural mechanism for the fidelity modulation of DNA polymerase lambda. J. Am. Chem. Soc. 138, 2389–2398 (2016).
Moon, A. F. et al. Sustained active site rigidity during synthesis by human DNA polymerase mu. Nat. Struct. Mol. Biol. 21, 253–260 (2014).
Delarue, M. et al. Crystal structures of a template-independent DNA polymerase: murine terminal deoxynucleotidyltransferase. EMBO J. 21, 427–439 (2002).
Volkman, B. F., Lipson, D., Wemmer, D. E. & Kern, D. Two-state allosteric behavior in a single-domain signaling protein. Science 291, 2429–2433 (2001).
Kerns, S. J. et al. The energy landscape of adenylate kinase during catalysis. Nat. Struct. Mol. Biol. 22, 124–131 (2015).
Murata, K. & Wolf, M. Cryo-electron microscopy for structural analysis of dynamic biological macromolecules. Biochim Biophys. Acta Gen. Subj. 1862, 324–334 (2018).
Tsai, M. D., Wu, W. J. & Ho, M. C. Enzymology and dynamics by cryogenic electron microscopy. Annu Rev. Biophys. 51, 19–38 (2022).
Dashti, A. et al. Trajectories of the ribosome as a Brownian nanomachine. Proc. Natl. Acad. Sci. USA 111, 17492–17497 (2014).
Frank, J. & Ourmazd, A. Continuous changes in structure mapped by manifold embedding of single-particle data in cryo-EM. Methods 100, 61–67 (2016).
Roh, S. H. et al. Subunit conformational variation within individual GroEL oligomers resolved by Cryo-EM. Proc. Natl. Acad. Sci. USA 114, 8259–8264 (2017).
Haselbach, D. et al. Structure and conformational dynamics of the human spliceosomal B(act) complex. Cell 172, 454–464 e411 (2018).
Murphy, B. J. et al. Rotary substates of mitochondrial ATP synthase reveal the basis of flexible F(1)-F(o) coupling. Science 364, eaaw9128 (2019).
Schulz, F., Abergel, C. & Woyke, T. Giant virus biology and diversity in the era of genome-resolved metagenomics. Nat. Rev. Microbiol 20, 721–736 (2022).
Sanchez-Cordon, P. J., Montoya, M., Reis, A. L. & Dixon, L. K. African swine fever: a re-emerging viral disease threatening the global pig industry. Vet. J. 233, 41–48 (2018).
Karger, A. et al. An update on African swine fever virology. Viruses 11, 864 (2019).
Coelho, J., Martins, C., Ferreira, F. & Leitao, A. African swine fever virus ORF P1192R codes for a functional type II DNA topoisomerase. Virology 474, 82–93 (2015).
Coelho, J., Ferreira, F., Martins, C. & Leitao, A. Functional characterization and inhibition of the type II DNA topoisomerase coded by African swine fever virus. Virology 493, 209–216 (2016).
Freitas, F. B., Frouco, G., Martins, C., Leitao, A. & Ferreira, F. In vitro inhibition of African swine fever virus-topoisomerase II disrupts viral replication. Antivir. Res. 134, 34–41 (2016).
Baylis, S. A., Dixon, L. K., Vydelingum, S. & Smith, G. L. African swine fever virus encodes a gene with extensive homology to type II DNA topoisomerases. J. Mol. Biol. 228, 1003–1010 (1992).
Coelho, J. & Leitao, A. The African swine fever virus (ASFV) topoisomerase II as a target for viral prevention and control. Vaccines (Basel) 8, 312 (2020).
Zhu, Z. & Meng, G. ASFVdb: an integrative resource for genomic and proteomic analyses of African swine fever virus. Database (Oxf.) 2020, baaa023 (2020).
Berger, J. M. & Osheroff, N. Structure and mechanism of eukaryotic type IIA topoisomerases. in DNA Topoisomerases and Cancer (ed Pommier, Y.) 87–101 (Springer, 2012).
Chen, S. H., Chan, N. L. & Hsieh, T. S. New mechanistic and functional insights into DNA topoisomerases. Annu Rev. Biochem 82, 139–170 (2013).
Pommier, Y., Sun, Y., Huang, S. N. & Nitiss, J. L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell Biol. 17, 703–721 (2016).
Sutormin, D. A. et al. Diversity and functions of type II topoisomerases. Acta Nat. 13, 59–75 (2021).
McKie, S. J., Neuman, K. C. & Maxwell, A. DNA topoisomerases: advances in understanding of cellular roles and multi-protein complexes via structure-function analysis. Bioessays 43, e2000286 (2021).
Berger, J. M., Gamblin, S. J., Harrison, S. C. & Wang, J. C. Structure and mechanism of DNA topoisomerase II. Nature 379, 225–232 (1996).
Fass, D., Bogden, C. E. & Berger, J. M. Quaternary changes in topoisomerase II may direct orthogonal movement of two DNA strands. Nat. Struct. Biol. 6, 322–326 (1999).
Corbett, K. D., Schoeffler, A. J., Thomsen, N. D. & Berger, J. M. The structural basis for substrate specificity in DNA topoisomerase IV. J. Mol. Biol. 351, 545–561 (2005).
Dong, K. C. & Berger, J. M. Structural basis for gate-DNA recognition and bending by type IIA topoisomerases. Nature 450, 1201–1205 (2007).
Schmidt, B. H., Osheroff, N. & Berger, J. M. Structure of a topoisomerase II-DNA-nucleotide complex reveals a new control mechanism for ATPase activity. Nat. Struct. Mol. Biol. 19, 1147–1154 (2012).
Wendorff, T. J., Schmidt, B. H., Heslop, P., Austin, C. A. & Berger, J. M. The structure of DNA-bound human topoisomerase II alpha: conformational mechanisms for coordinating inter-subunit interactions with DNA cleavage. J. Mol. Biol. 424, 109–124 (2012).
Laponogov, I. et al. Structure of an ‘open’ clamp type II topoisomerase-DNA complex provides a mechanism for DNA capture and transport. Nucleic Acids Res. 41, 9911–9923 (2013).
Wang, Y. et al. Structure of African swine fever virus and associated molecular mechanisms underlying infection and immunosuppression: a review. Front. Immunol. 12, 715582 (2021).
Brino, L. et al. Dimerization of Escherichia coli DNA-gyrase B provides a structural mechanism for activating the ATPase catalytic center. J. Biol. Chem. 275, 9468–9475 (2000).
Hu, T., Sage, H. & Hsieh, T. S. ATPase domain of eukaryotic DNA topoisomerase II. Inhibition of ATPase activity by the anti-cancer drug bisdioxopiperazine and ATP/ADP-induced dimerization. J. Biol. Chem. 277, 5944–5951 (2002).
Classen, S., Olland, S. & Berger, J. M. Structure of the topoisomerase II ATPase region and its mechanism of inhibition by the chemotherapeutic agent ICRF-187. Proc. Natl. Acad. Sci. USA 100, 10629–10634 (2003).
Bates, A. D. & Maxwell, A. The role of ATP in the reactions of type II DNA topoisomerases. Biochem Soc. Trans. 38, 438–442 (2010).
Blower, T. R. et al. A complex suite of loci and elements in eukaryotic type II topoisomerases determine selective sensitivity to distinct poisoning agents. Nucleic Acids Res. 47, 8163–8179 (2019).
Laponogov, I. et al. Trapping of the transport-segment DNA by the ATPase domains of a type II topoisomerase. Nat. Commun. 9, 2579 (2018).
Chen, S. F. et al. Structural insights into the gating of DNA passage by the topoisomerase II DNA-gate. Nat. Commun. 9, 3085 (2018).
Zhao, Y. et al. Cryo-EM structures of African swine fever virus topoisomerase. mBio 14, e0122823 (2023).
Ho, B. K., Coutsias, E. A., Seok, C. & Dill, K. A. The flexibility in the proline ring couples to the protein backbone. Protein Sci. 14, 1011–1018 (2005).
Wang, J. C. Moving one DNA double helix through another by a type II DNA topoisomerase: the story of a simple molecular machine. Q Rev. Biophys. 31, 107–144 (1998).
Soczek, K. M., Grant, T., Rosenthal, P. B. & Mondragon, A. CryoEM structures of open dimers of gyrase A in complex with DNA illuminate mechanism of strand passage. Elife 7, e41215 (2018).
Schmidt, B. H., Burgin, A. B., Deweese, J. E., Osheroff, N. & Berger, J. M. A novel and unified two-metal mechanism for DNA cleavage by type II and IA topoisomerases. Nature 465, 641–644 (2010).
Pommier, Y., Leo, E., Zhang, H. & Marchand, C. DNA topoisomerases and their poisoning by anticancer and antibacterial drugs. Chem. Biol. 17, 421–433 (2010).
Wu, C. C. et al. Structural basis of type II topoisomerase inhibition by the anticancer drug etoposide. Science 333, 459–462 (2011).
Lee, I., Dong, K. C. & Berger, J. M. The role of DNA bending in type IIA topoisomerase function. Nucleic Acids Res. 41, 5444–5456 (2013).
Wu, C. C., Li, Y. C., Wang, Y. R., Li, T. K. & Chan, N. L. On the structural basis and design guidelines for type II topoisomerase-targeting anticancer drugs. Nucleic Acids Res. 41, 10630–10640 (2013).
Vanden Broeck, A. et al. Structural basis for allosteric regulation of Human Topoisomerase IIalpha. Nat. Commun. 12, 2962 (2021).
Baird, C. L., Harkins, T. T., Morris, S. K. & Lindsley, J. E. Topoisomerase II drives DNA transport by hydrolyzing one ATP. Proc. Natl. Acad. Sci. USA 96, 13685–13690 (1999).
Maciejewski, M. W. et al. Solution structure of a viral DNA repair polymerase. Nat. Struct. Biol. 8, 936–941 (2001).
Showalter, A. K., Byeon, I. J., Su, M. I. & Tsai, M. D. Solution structure of a viral DNA polymerase X and evidence for a mutagenic function. Nat. Struct. Biol. 8, 942–946 (2001).
Chen, Y. et al. A unique DNA-binding mode of African swine fever virus AP endonuclease. Cell Discov. 6, 13 (2020).
Showalter, A. K. & Tsai, M. D. A DNA polymerase with specificity for five base pairs. J. Am. Chem. Soc. 123, 1776–1777 (2001).
Voehler, M. W., Eoff, R. L., McDonald, W. H., Guengerich, F. P. & Stone, M. P. Modulation of the structure, catalytic activity, and fidelity of African swine fever virus DNA polymerase X by a reversible disulfide switch. J. Biol. Chem. 284, 18434–18444 (2009).
Wu, W. J. et al. How a low-fidelity DNA polymerase chooses non-Watson-Crick from Watson-Crick incorporation. J. Am. Chem. Soc. 136, 4927–4937 (2014).
Lamarche, B. J., Showalter, A. K. & Tsai, M. D. An error-prone viral DNA ligase. Biochemistry 44, 8408–8417 (2005).
Showalter, A. K. et al. Mechanistic comparison of high-fidelity and error-prone DNA polymerases and ligases involved in DNA repair. Chem. Rev. 106, 340–360 (2006).
Wang, G., Xie, M., Wu, W. & Chen, Z. Structures and functional diversities of ASFV proteins. Viruses 13, 2124 (2021).
Daniel, E. & Koshland, J. The key-lock theory and the induced fit theory. Angew. Chem. Int. Ed. Engl. 33, 2375–2378 (1994).
Baek, M. et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 373, 871–876 (2021).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr D Struct. Biol. 74, 531–544 (2018).
Ling, E. M. et al. A comprehensive structural analysis of the ATPase domain of human DNA topoisomerase II beta bound to AMPPNP, ADP, and the bisdioxopiperazine, ICRF193. Structure 30, 1129–1145 e1123 (2022).
Acknowledgements
The authors thank Dr. Alexandre Leitão for kindly providing the original AsfvTop2 protein expression plasmid, and Dr. Nei-Li Chan and Dr. Lars-Oliver Essen for the comments during manuscript preparation The work was supported by Academia Sinica (AS-KPQ-109-TPP2) and Taiwan Cryo-EM Consortium funded by National Council of Science and Technology, grant no. NSTC 112-2740-B-006-001. Cryo-EM experiments were performed at the Academia Sinica Cryo-EM Facility, which is supported by Academia Sinica (AS-CFII-108-110) and Taiwan Protein Project (AS-KPQ-109-TPP2). The cryo-EM data were processed at the Academia Sinica Grid-computing Center supported by Academia Sinica. The X-ray crystallography data were collected at the Taiwan Photo Source (TPS) beamline 05 A, National Synchrotron Radiation Research Center (NSRRC), Taiwan. NMR data were collected at the Academia Sinica High Field Nuclear Magnetic Resonance Center (Grant No. AS-CFII-108-112). We thank Dr. Shu-Yu Lin for the support on the Mass spectrometry experiments conducted at the Proteomics Mass Spectrometry Common Facility, Institute of Biological Chemistry, Academia Sinica.
Author information
Authors and Affiliations
Contributions
C.-W.C. and M.-D.T. conceived and designed the research and wrote the paper. S.-C.W. characterized AsfvTop2 preferential DNA sequences and designed experiments for protein expression, purification and activity assays. C.-W.C. and C.-H.W. performed cryo-EM experiments. C.-W.C., A.H.P., and Y.-K.C. performed X-ray crystallography experiments. W.-J.W. performed NMR experiments. C.-H.Y. and all other authors analyzed data.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications chemistry thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chang, CW.M., Wang, SC., Wang, CH. et al. A unified view on enzyme catalysis by cryo-EM study of a DNA topoisomerase. Commun Chem 7, 45 (2024). https://doi.org/10.1038/s42004-024-01129-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42004-024-01129-y
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
