
Abstract
Monotremata is a clade of egg-lying mammals, represented by the living platypus and echidnas, which is endemic to Australia, and adjacent islands. Occurrence of basal monotremes in the Early Cretaceous of Australia has led to the consensus that this clade originated on that continent, arriving later to South America. Here we report on the discovery of a Late Cretaceous monotreme from southern Argentina, demonstrating that monotremes were present in circumpolar regions by the end of the Mesozoic, and that their distinctive anatomical features were probably present in these ancient forms as well.
Similar content being viewed by others


Introduction
The fossil record and extant distribution of monotremes is almost restricted to Australasia, with the single exception of a fossil ornithorhynchid from the earliest Cenozoic in Patagonia1. In this context, occurrence of a monotreme in Patagonia was interpreted as the result of a single dispersal from Australia to South America, before or during the Late Cretaceous or early Paleocene2,3,4,5,6.
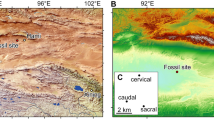
The aim of present contribution is to report on the discovery of a new monotreme, Patagorhynchus pascuali n.gen. et sp., which represents the first Cretaceous toothed monotreme from Gondwana. The material here reported consists of a fragmentary right jaw preserving the lower molar 2 showing the dilambdodon pattern characteristic of monotremes. The molar was collected from levels of the Chorrillo Formation (Upper Cretaceous, Early Maastrichtian7), cropping out in SW Santa Cruz province, Patagonia, Argentina. It was found in association with both terrestrial and aquatic mollusks, calyptocephalellid anurans, chelid turtles, snakes, ornithopod, sauropod, and non-avian and avian theropod remains7,8. As far as mammals is concerned, the same fossil spot yielded a molar of the gondwanatherian Magallanodon baikashkenke9 and isolated caudal vertebrae regarded as Mammalia incertae sedis8. From almost equivalents levels belonging to the Dorotea Formation (Valle del Río de las Chinas, southern Chile), remains of Magallanodon and the meridiolestidan Orretherium have been reported10,11.
Results
Mammalia, Linnaeus 1758
Monotremata, Bonaparte, 1837
Patagorhynchus gen. nov. (monotypic genus)
Etymology
Patago, from Patagonia, and rhynchus, nose.
Diagnosis
Patagorhynchus differs from basal monotremaformes (including Steropodon) in having a dilambdodont crown morphology and a labial cingulid;12 dilambdodont disposition of cusps and crests on molar crown is shared with Teinolophos and ornithorhynchids; Patagorhynchus and ornithorhynchids differ from the basal monotremaformes Teinolophos in having notably low and mesiodistally expanded teeth with the anterior lobe (equivalent to trigonid) positioned lower than the posterior one (equivalent to talonid), in having talonid composed of two (rather than one) transverse lophids, and lacking a labial cingulid. The anterior cingulid of Patagorhynchus is wider than that in Teinolophos but narrower than that in Obdurodon. Patagorhynchus shares with the toothed monotremes Obdurodon and Monotrematum both lingual and buccal extremes of the V-shaped lobe (equivalent to trigonid) with one buccal and two lingual cusps, with the first being more elevated than the latter two, and a complete mid-valley. Patagorhynchus bears two roots on m2 (as also probably in Monotrematum) and differs from Obdurodon and Ornithorhynchus, in which more than 5 roots are present. The lobes of Patagorhynchus and Obdurodon show hypsodonty, in contrast with the much more brachyodont molariforms of Monotrematum. Patagorhynchus exhibits the following features that are lacking in other monotremes, and may be considered autapomorphic among monotremes: mid-valley labially diverges (i.e., the length of the labial edge of this valley represents two times its lingual length) and anterior cingulid labially narrow and does not reach the labial margin of the protoconid.
Type and the only species
Patagorhynchus pascuali sp. nov. (Figs. 1, 2a and Supplementary Fig. 3).

Lower molar 2 and part of the right jaw, in a, occlusal view; b, medial/lingual view; c, lateral/labial view; d, posterior view; e, anterior view. Scale bar: length 2 mm. Abbreviations, ac, anterior cingulid; alv, alveolus; ant, anterior; ar, anterior root; hy, hypoconid; hl, hypoconulid; lapcc, labial posterior cingular cusp; liacc, lingual anterior cingular cusp; me, metaconid; mv, mid-valley; NC1, neomorphic cusp 1; pa, paraconid; pc, posterior cingulid; pr, protoconid; prt, posterior root.

Patagorhynchus pascuali sp. nov.
Etymology
Species name honors the Argentine paleomammalogist Rosendo Pascual (1923–2012), who described the first Cenozoic monotreme remains from Patagonia, thus demonstrating the presence of this clade outside Australia.
Holotype
MPM-PV-23087, Museo Padre Molina (Rio Gallegos, Santa Cruz, Argentina), a right lower m2 attached to a fragment of dentary. Collected by N. R. Chimento during a joint Argentine-Japanese field trip in March 2022.
Diagnosis
The same as for genus by monotypy.
Type locality and age
La Anita farm, Santa Cruz Province, Patagonia, Argentina. The tooth was collected from the “Puma Cave” fossil site (S 50 30.639 W 72 33.617), Chorrillo Formation, early Maastrichtian7,8. This new discovery expands the list of Late Cretaceous mammaliaforms recorded in the Chorrillo Formation and equivalent Dorotea Formation in southern Chile, previously known to include gondwanatherians (Magallanodon) and dryolestoids (Orretherium)9,10,11,13.
Description
Despite the occlusal surface being somewhat damaged, the morphology of the main cusps and anatomical details can be clearly discerned. The tooth is identified as a second lower molar based on the similarities with the m2 of Obdurodon, including a subrectangular-shaped outline in occlusal view, the presence of two lobes each bearing three cusps, a mid-valley lacking cusps, and prominent anterior and posterior cingulids (Fig. 1). Immediately anterior to m2, the fragmentary mandible shows a partially preserved and relatively small alveolus on the labial margin, which presumably corresponds to one of the roots of m1.
The Patagorhynchus m2 exhibits a distinct morphology that easily identifies it as a monotreme. This includes a unique lophid and cusp structure resulting in the presence of two mesiodistally compressed lobes that are sub-equally in shaped and size each consisting of three cusps, twinned paraconid and metaconid, wrapping cingulid, hypsodont lobes, and un-basined talonid12,14,15.
The m2 is 5.8 mm in mesiodistal length (see Supplementary Results 3), indicating that this tooth of Patagorhynchus was possibly intermediate in size between Monotrematum and some species of Obdurodon. The m2 is mesiodistally longer than transversely wide, and narrows mesially. Six large cusps are present: protoconid, paraconid, metaconid, hypoconid, hypoconulid, and NC1 (neomorphic cuspid 1)16. These cusps are relatively low and mound-like and connected by lophids, which form two main lobes or triakididrepanids (Fig. 1, Supplementary Fig. 2).
The anterior lobe (equivalent to trigonid) is labiolingually narrower and apicobasally taller than the posterior lobe (equivalent to a talonid), a condition shared with Obdurodon12. In Patagorhynchus, the anterior lobe is heart-shaped, with the anterior and posterior lophids being slightly convex posteriorly. This results in the paraconids being located anteriorly at the same level as the protoconid. In Obdurodon, by contrast, the anterior lophid is anteriorly convex and the posterior one is straight, resulting in metaconid and protoconid being located at the same level. In Patagorhynchus, the paraconid is larger than the metaconid, and its base is ventrally positioned relative to the base of both the metaconid and protoconid, suggesting that the paraconid was more ventrally located than the other cusps.
The posterior (talonid) lobe is similar in shape to the anterior (trigonid) lobe, but much wider labiolingually. The lingual cusps are notably taller than the labial one (hypoconid). The preserved bases of the NC1 and hypoconulid are subequal in size and position. The hypoconid is mesiodistally narrower than the protoconid.
Between the lingual cusps of paired lobes there is a narrow, eye-shaped enamel invagination, reminiscent of a flexid. Such a condition is also present in Monotrematum17 and some specimens of Obdurodon18. Patagorhynchus resembles Monotrematum in that the invaginations are delimited by a narrow enamel layer (Fig. 2, Supplementary Fig. 2), in contrast to Obdurodon in which the invaginations are labiolingually extended19.
Both lobes are separated by a wide, deep mid-valley, which extends from the labial through the lingual edges of the tooth. The margins of the valley widen slightly towards the labial edge of the tooth. The valley lacks well-defined cusps or fossettes, and becomes deeper towards its labial margin.
Posterior and anterior cingulids are prominent, being wider than those in Teinolophos but narrower than those in Obdurodon16,20. The posterior cingulum is eroded on its lingual end, but the preserved segment maintains a constant width along its length, being similar in this morphology to that in Obdurodon. In contrast, the cingulids become lingually wider in Teinolophos20. The anterior cingulid hosts a small cusp on its lingual end, whereas in the posterior cingulid the labial end shows a cusp (the lingual end of this cingulid is eroded, precluding the recognition of cusps), similar to the morphology in Monotrematum, but differing from that in Obdurodon (Fig. 2, Supplementary Fig. 2).
The tooth bears two roots that are broad labiolingually and constricted at mid-height; they are obliquely oriented with respect to the main axis of the tooth. Regarding the root number, Patagorhynchus retains the ancestral condition in m2 shared with Teinolophos (and probably Monotrematum), differing from the multiple roots present in Obdurodon and Ornithorhynchus16,20.
Discussion
As indicated above, the crown shape of Patagorhynchus unambiguously indicates that this taxon belongs to monotremes. With the aim to test the phylogenetic position of Patagorhynchus, we scored this tooth into a previously published data matrix composed by 558 characters and 128 taxa21 (see Supplementary Methods 1 and 2). We concentrated on the characters available in this new tooth, a total of 54 characters can be scored for Patagorhynchus (Supplementary Results 1 and 2). The results of the phylogenetic analysis consistently place Patagorhynchus as nested within monotremes, together with the genera Ornithorhynchus, Tachyglossus, Monotrematum and Obdurodon (Fig. 3, Supplementary Fig. 1).

Basal Monotremaformes44 are indicated in red and Monotremata in green. The Late Cretaceous (Maastrichtian) palaeogeographical map (based in Scotese35) indicates the fossiliferous sites that yielded fossil toothed monotremes and distribution of the extant platypus Ornithorhynchus anatinus shaded in light brown. [1], occurrence of Patagorhynchus pascuali, La Anita farm, Chorrillo Formation (Maastrichtian, Late Cretaceous); [2], occurrence of Monotrematum sudamericanum, Punta Peligro locality, Salamanca Formation (Danian, lower Paleocene); [3], occurrence of Obdurodon spp., different localities from South Australia, Queensland, and New South Wales Oligocene-Pliocene); [4], Pleistocene occurrences and geographic distribution of extant Ornithorhynchus anatinus.
Australia has yielded the most complete fossil record of monotremes2, including an array of Barremian through Cenomanian taxa, as well as several species of the Oligocene-Pliocene monotreme Obdurodon. In this context, the presence of the toothed monotreme Monotrematum in the early Paleocene of Patagonia1,22 was interpreted as the result of a single dispersal of monotremes from Australia to South America, before or during the Late Cretaceous or early Paleocene2,3,4,5,6. Discovery of Patagorhynchus clearly demonstrates that the monotremes had already attained a wide paleogeographic distribution, stretching across southern South America, Australia, and Antarctica, the later one as a connecting pathway (but fossil monotremes are still unknown from this landmass), constituting a clade characteristic of the Weddelian Paleobiogeographical Province23,24,25,26,27,28.
The new discovery expands the list of mammals documented in the Chorrillo and equivalent Dorotea formations of southern South America, adding the Monotremata to the assemblage of non-therian mammals (i.e., gondwanatherians and meridiolestidan dryolestoids9,10,11,13). Remarkably, monotremes are absent from the extensively sampled Late Cretaceous localities of northern and central Patagonia2,29,30. Such a difference among mammalian assemblages characteristic of Patagonia is consistent with the uneven distribution of non-avian dinosaurs in this region. For example, megaraptorid theropods, colossosaurian titanosaurs, and elasmarian iguanodontians are numerically dominant in the Chorrillo Formation8,31 whereas abelisaurid theropods and saltasaurine titanosaurs are prevalent in coeval units in northern Patagonia. Similar differences are documented in terrestrial and marine biotas between southern and northern Patagonia32,33,34. Thus, evidence at hand suggests that the Maastrichtian vertebrate fauna in southern Patagonia was different from that in northern Patagonia. It is noteworthy that the former had, instead, several taxa in common with Australia (e.g., Monotremata, Megaraptoridae). It is likely that a latitudinal zonation of environmental conditions (i.e., dry and warm in northern Patagonia versus humid and cold in southern Patagonia) controlled the distribution and partial abundance of the above-mentioned vertebrate clades.
The presence of monotremes in the southern La Anita fossil site (which occupied a paleolatitude of approximately 60° S during the Maastrichtian, roughly the same as that of southern Australia35) is congruent with the interpretation by Flannery et al.2 that monotremes evolved under humid, cool and densely forested environments in circumpolar Gondwana. Some authors already proposed that certain anatomical and physiological characteristics of living monotremes (e.g., low metabolism, a mechanoreceptive and electroreceptive beak for probe feeding, and relatively large body size) may have evolved in the context of polar environments2,18,36.
The crown morphology of the only available molar of Patagorhynchus is closely similar to that of the Paleogene Monotrematum and the Neogene Obdurodon, revealing a highly conservative dental morphology for toothed monotremes15. Remarkably, this molar pattern underwent only minor changes for approximately 60 million years from the Late Cretaceous through to Miocene times. This duration of stasis in dental morphology considerably exceeds that seen in other mammalian groups (e.g., therians and dryolestoids37,38,39,40).
The labiolingually broad segment of the molar of Patagorhynchus and the reduction in the number of teeth (eventually restricted to only two upper molars inferred for Monotrematum2) may be congruent with the duck-billed morphology of the snout documented in more derived ornithorhynchids. In addition, the presence of a hypertrophied mandibular canal in Teinolophos suggests the development of electroreception occurred early in the evolutionary history of Monotremata and that the acquisition of a specialized duckbill for high-resolution aquatic electroreception is unique to the clade39. Based on such evidence, we hypothesize that a highly sensitive duck-billed snout is likely to have already been present in Late Cretaceous monotremes, such as Patagorhynchus. Apparently, a similar anatomical inference could be made for the rest of the body, as suggested by the morphology of the distal femur of Monotrematum41 being almost identical to that of the living platypus. As in Ornithorhynchus, extinct monotremes may have had a sprawling posture of their hind limbs, and eventually adapted for swimming42. The possibility that Patagorhynchus had already acquired ecological and behavioral characteristics similar to those of the living platypus, which inhabits ponds and lakes, is congruent with sedimentological evidence suggesting that such environments were prevalent during deposition of the Chorrillo Formation7, as well as with occurrences of Nymphaeaceae aquatic plants, freshwater snails and abundant larvae of chironomid insects, with the latter two invertebrates constituting part of the food for the living platypuses8,36,43.
Discovery of Patagorhynchus gives an insight into the degree of continuity between the terrestrial vertebrate faunas of western and eastern Gondwana during the Late Cretaceous, suggesting the lack of paleobiogeographic barriers to their dispersal prior to the deep-water opening of the Drake Passage and the Tasman Gateway. The diversification of monotremes towards the end of the Mesozoic suggested by the present discovery implies that an extensive and still unknown history of this clade of peculiar mammals awaits to be documented in Mesozoic beds in southern South America.
Methods
The material reported in this publication was collected from the Chorrillo Formation (Upper Cretaceous, lower Maastrichtian7) cropping out in La Anita fossil site, SW Santa Cruz Province, Patagonia, Argentina. The specimen was found in association with both terrestrial and aquatic mollusks, calyptocephalellid anurans, chelid turtles, snakes, ornithopods, sauropods, and non-avian and avian theropod remains7,8. With regard to mammals, the same outcrop yielded remains of the gondwanatherian Magallanodon baikashkenke and isolated caudal vertebrae of yet unidentified mammals8,9,13.
Cusp nomenclature of molariforms used in the description and codifications of Monotrematum and Patagorhynchus follows the terminology applied by Kielan-Jaworowska et al.45, Rich et al.46, and Woodburne16.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “http://zoobank.org/”. The LSID for this publication is: 01EF7079-F4F8-4996-ABD3-D61BBD04A2BA.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The datasets analyzed during the current study are included in this published article (and its Supplementary Information file).
References
Pascual, R. et al. First discovery of monotremes in South America. Nature 356, 704–706 (1992).
Flannery, T. F. et al. A review of monotreme (Monotremata) evolution. Alcheringa 46, 1–18 (2022).
Rich, T. H. & Vickers-Rich, P. Palaeobiogeographic implications of Early Cretaceous Australian placental mammals. Proc. Indian Nat. Sci. Acad., A 65, 315–327 (1999).
Rich, T. H. et al. Early Cretaceous mammals from Flat Rocks, Victoria, Australia. Rec. Queen Vic. Mus. 106, 1–35 (1999).
Beck, R. M. D. The biogeographical history of non-marine mammaliaforms in the Sahul region in Handbook of Australasian biogeography (ed. Ebach, M. C.) 329–366 (Boca Raton, CRC Press, 2017).
Musser, A. M. Review of the monotreme fossil record and comparison of palaeontological and molecular data. Comp. Biochem Physiol. A: Mol. Integr. Physiol. 136, 927–942 (2003).
Moyano-Paz, D. et al. The uppermost Cretaceous continental deposits at the southern end of Patagonia, the Chorrillo Formation case study (Austral-Magallanes Basin): Sedimentology, fossil content and regional implications. Cretac. Res 130, 105059 (2022).
Novas, F. E. et al. Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina. Rev Mus Argentino. Cienc. Nat., n. s. 21, 217–293 (2019).
Chimento, N. R., Agnolin, F. L., Tsuihiji, T., Manabe, M. & Novas, F. E. New record of a Mesozoic gondwanatherian mammaliaform from Southern Patagonia. Sci. Nat. 107, 1–7 (2020).
Goin, F. J. et al. First Mesozoic mammal from Chile: The southermost record of a Late Cretaceous gondwanatherian. Bol. Mus. Nac. Hist. Nat. Chile 69, 5–31 (2020).
Martinelli, A. G. et al. New cladotherian mammal from southern Chile and the evolution of mesungulatid meridiolestidans at the dusk of the Mesozoic Era. Sci. Rep. 11, 1–18 (2021).
Archer, M., Flannery, T. F., Ritchie, A. & Molnar, R. E. First Mesozoic mammal from Australia-an early Cretaceous monotreme. Nature 318, 363–366 (1985).
Chimento, N. R., Agnolin, F. L., Tsuihiji, T., Manabe, M. & Novas, F. E. New gondwanatherian (Mammaliaformes) remains from the Chorrillo Formation (Upper Cretaceous) of southern Patagonia, Argentina. Cretac. Res 127, 104947 (2021).
Luo, Z. X., Cifelli, R. L. & Kielan-Jaworowska, Z. Dual origin of tribosphenic mammals. Nature 409, 53–57 (2001).
Pian, R., Archer, M. & Hand, S. J. A new, giant platypus, Obdurodon tharalkooschild, sp. nov (Monotremata, Ornithorhynchidae), from the Riversleigh World Heritage Area, Australia. J. Vertebr. Paleontol. 33, 1255–1259 (2013).
Woodburne, M. O. Monotremes as pretribosphenic mammals. J. Mamm. Evol. 10, 195–248 (2003).
Pascual, R., Goin, F. J., Balarino, M. L. & Udrizar Sauthier, D. E. New specimens of Monotrematum sudamericanum, and the convergent evolution of the triangulate molar. Acta Palaeontol. Pol. 47, 487–492 (2002).
Archer, M., Murray, P., Hand, S. & Godthelp, H. Reconsideration of monotreme relationships based on the skull and dentition of the Miocene Obdurodon dicksoni in Mammal Phylogeny (eds. Szalay, F. S., Novacek, M. J. & McKenna, M. C.) 75–94 (New York, Springer, 1993).
Woodburne, M. O. & Tedford, R. H. The first Tertiary monotreme from Australia. Am. Mus. Nov. 2588, 1–11 (1975).
Rich, T. H. et al. The mandible and dentition of the Early Cretaceous monotreme Teinolophos trusleri. Alcheringa 40, 475–501 (2016).
Zhou, C. F., Bhullar, B. A. S., Neander, A. I., Martin, T. & Luo, Z.-X. New Jurassic mammaliaform sheds light on early evolution of mammal-like hyoid bones. Science 365, 276–279 (2019).
Pascual, R. et al. The first non-Australian monotreme: an early paleocene South American platypus (Monotremata, Ornithorhynchidae) in Platypus and Echidnas (ed. Augee, M. L.) 1–14 (Royal Zoological Society of New South Wales, 1992b).
Case, J. A., Woodburne, M. O. & Chaney, D. S. A new genus and species of polydolopid marsupial from the La Meseta Formation, Late Eocene, Seymour Island, Antarctic Peninsula in Geology and paleontology of Seymour Island (eds. Feldmann, R. M. & Woodburne, M. O.) 505–521 (Geological Society of America, Boulder, 1988).
Shen, Y. Cretaceous and Paleogene biogeography in Antarctic Peninsula and its significance in the reconstruction of Gondwanaland in Stratgraphy and palaeontology of Fildes Peninsula, King George Island, Antarctic (ed. Shen, Y.) 246–258 (Beijing: Science Press, 1994).
Case, J. A. et al. The first duckebilled dinosaur (family Hadrosauridae) from Antarctica. J. Vert. Paleontol. 20, 612–614 (2000). [0612:TFDBDF]2.0.CO;2.
Agnolin, F. L., Ezcurra, M. D., Pais, D. F. & Salisbury, W. A reappraisal of the Cretaceous nonavian dinosaur faunas from Australia and New Zealand, evidence for their Gondwanan affinities. J. Syst. Palaeontol. 8, 257–300 (2010).
Rozadilla, S. et al. A new ornithopod (Dinosauria, Ornithischia) from the Upper Cretaceous of Antarctica and its palaeobiogeographical implications. Cretac. Res 57, 311–324 (2016).
Agnolín, F. L., Brisson Egli, F., Chatterjee, S., García-Marsà, J. A. & Novas, F. E. Vegaviidae, a new clade of southern diving birds that survived the K/T boundary. Sc. Nat. 104, 1–9 (2017).
Flannery, T. F., Archer, M., Rich, T. H. & Jones, R. A new family of monotremes from the Cretaceous of Australia. Nature 377, 418–420 (1995).
Rich, T. H. The palaeobiogeography of Mesozoic mammals: A review. Arquivos Mus. Nac. Rio de. Jan. 66, 231–249 (2008).
Novas, F. E., Agnolín, F. L., Ezcurra, M. D., Porfiri, J. & Canale, J. I. Evolution of the carnivorous dinosaurs during the Cretaceous: the evidence from Patagonia. Cretac. Res 45, 174–215 (2013).
Woodburne, M. O. et al. Paleogene land mammal faunas of South America; a response to global climatic changes and indigenous floral diversity. J. Mamm. Evol. 21, 1–73 (2014).
Bogan, S. et al. A new species of the genus Echinorhinus (Chondrichthyes, Echinorhiniformes) from the Upper Cretaceous of southern South America (Argentina-Chile). Cretac. Res 78, 89–94 (2017).
Reguero, M. A. & Goin, F. J. Paleogeography and biogeography of the Gondwanan final breakup and its terrestrial vertebrates: New insights from southern South America and the “double Noah’s Ark” Antarctic Peninsula. J. South Am. Earth Sci. 108, 103358 (2021).
Scotese, C. R. Map Folio 17, Late Cretaceous (Maastrichtian, 68 Ma), PALEOMAP PaleoAtlas for ArcGIS, Volume 2, Cretaceous Paleogeographic, Paleoclimatic and Plate Tectonic Reconstructions, PALEOMAP Project (Evanston, IL, 2013).
Bino, G. et al. The platypus: evolutionary history, biology, and an uncertain future. J. Mammal. 100, 308–327 (2019).
Bonaparte, J. F. New Late Cretaceous mammals from the Los Alamitos Formation, northern Patagonia. Nat. Geogr. Res 6, 63–93 (1990).
Bonaparte, J. F. Approach to the significance of the Late Cretaceous mammals of South America. Berl. Geowissenschaftliche Abhandl E 13, 31–44 (1994).
Rowe, T., Rich, T. H., Vickers-Rich, P., Springer, M. & Woodburne, M. O. The oldest platypus and its bearing on divergence timing of the platypus and echidna clades. Proc. Natl Acad. Sci. 105, 1238–1242 (2008).
Luo, Z. X. Transformation and diversification in early mammal evolution. Nature 450, 1011–1019 (2007).
Forasiepi, A. M. & Martinelli, A. G. Femur of a monotreme (Mammalia, Monotremata) from the early Paleocene Salamanca Formation of Patagonia, Argentina. Ameghiniana 40, 625–630 (2003).
Westerman, M. & Edwards, D. DNA hybridization and the phylogeny of monotremes in Platypus and Echidnas (ed. Augee, M. L.) 28–34 (Royal Zoological Society of New South Wales, 1992).
Vera, E. I. et al. Paleobotany of the uppermost Cretaceous Chorrillo Formation, Santa Cruz Province, Argentina: insights in a freshwater floral community. Cretac. Res 138, 105296 (2022).
Luo, Z. X. Developmental patterns in Mesozoic evolution of mammal ears. Annu Rev. Ecol. Evol. Syst. 42, 355–380 (2011).
Kielan-Jaworowska, Z., Crompton, A. W. & Jenkins, F. A. The origin of egg-laying mammals. Nature 326, 871–873 (1987).
Rich, T. H. et al. Monotreme nature of the Australian Early Cretaceous mammal Teinolophos. Acta Palaeontol. Pol. 46, 113–118 (2001).
Acknowledgements
The present paper is the result of a second Argentine-Japanese expedition, carried out in March 2022. We thank Coleman Burke (New York) for his encouragement and financial assistance to carry out the first field explorations to La Anita farm. A special thanks to Dr. Y. Harashi, former General Director of the National Museum of Nature and Science, Japan. Special thanks to Federico Braun for allowing access to his property. Facundo Echeverría and his wife Daphne Fraser (La Anita farm) offered their valuable geographic knowledge of these territories, allowing us an easy access to fossil sites with our 4×4 vehicles. Oscar Canto and Carla Almazán (Secretaría de Cultura) for supporting our projects and explorations in Santa Cruz. Also, we thank Marcelo Isasi (LACEV-MACN, CONICET) for the technical preparation of the specimen. Thanks to geologists Francisco Nullo, Damián Moyano Paz, Maria Sol Raigemborn, Sabrina Lizzoli and Daniel Poiré provided valuable comments on the stratigraphy and regional geology of the Chorrillo Formation. Thanks to Jorge Barrientos for providing the bibliography. Thanks to Mauro Aranciaga Rolando, Gerardo Álvarez Herrero, Ana Moreno Rodríguez, F. Brissón Eglí, G. Muñoz, Santiago Miner, David Piazza, Adrian Misantone (LACEV-MACN) and Mateo Monferran (CECOAL-CONICET) which made part of the field crew to La Anita farm. We thank two anonymous reviewers, Steve Brusatte and Robin Beck, for their enlightening comments that greatly improved the quality of this paper. This work was supported by The Susan and Coleman Burke Foundation (New York), and a grant from the National Museum of Nature and Science, Japan.
Author information
Authors and Affiliations
Contributions
N.R.C. and F.L.A. carried out the fieldwork and discovery of the described specimen. N.R.C., F.L.A. and F.E.N. conceptualized the study. N.R.C., F.L.A., T.H.R. and P.V.R. phylogenetic analyses. F.E.N., M.M. and T.T. acquired fundings. N.R.C. and F.L.A. made the figures. All authors contributed to the writing and general revision of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests
Peer review
Peer review information
Communications Biology thanks Robin Beck and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Luke Grinham.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chimento, N.R., Agnolín, F.L., Manabe, M. et al. First monotreme from the Late Cretaceous of South America. Commun Biol 6, 146 (2023). https://doi.org/10.1038/s42003-023-04498-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-023-04498-7
This article is cited by
-
A Maastrichtian insect assemblage from Patagonia sheds light on arthropod diversity previous to the K/Pg event
Communications Biology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
