
Abstract
Rhodopila globiformis is the most acidophilic of anaerobic purple phototrophs, growing optimally in culture at pH 5. Here we present a cryo-EM structure of the light-harvesting 1–reaction center (LH1–RC) complex from Rhodopila globiformis at 2.24 Å resolution. All purple bacterial cytochrome (Cyt, encoded by the gene pufC) subunit-associated RCs with known structures have their N-termini truncated. By contrast, the Rhodopila globiformis RC contains a full-length tetra-heme Cyt with its N-terminus embedded in the membrane forming an α-helix as the membrane anchor. Comparison of the N-terminal regions of the Cyt with PufX polypeptides widely distributed in Rhodobacter species reveals significant structural similarities, supporting a longstanding hypothesis that PufX is phylogenetically related to the N-terminus of the RC-bound Cyt subunit and that a common ancestor of phototrophic Proteobacteria contained a full-length tetra-heme Cyt subunit that evolved independently through partial deletions of its pufC gene. Eleven copies of a novel γ-like polypeptide were also identified in the bacteriochlorophyll a-containing Rhodopila globiformis LH1 complex; γ-polypeptides have previously been found only in the LH1 of bacteriochlorophyll b-containing species. These features are discussed in relation to their predicted functions of stabilizing the LH1 structure and regulating quinone transport under the warm acidic conditions.
Similar content being viewed by others

Introduction
The photooxidized bacteriochlorophyll (BChl) dimer (special pair) in the photosynthetic reaction center (RC) of anoxygenic phototrophic bacteria is reduced either directly by a soluble redox protein or indirectly through a membrane-associated heme protein interpositioned between the soluble components and the photooxidized special pair, or by both in some species. Soluble electron donors are typically a small heme protein or a high-potential iron-sulfur protein, while the electron mediating membrane proteins include the RC-bound multi-heme cytochrome (Cyt) in many purple bacteria1 and in green filamentous bacteria2,3, or the multiple copies of a membrane-anchored mono-heme Cyt found in some purple bacteria4 and also in green sulfur bacteria5,6 and the heliobacteria5,7. Electron mediation through a membrane-associated Cyt is most common1 and is thought to be a more ancient form of photosynthesis1,5,8.
In purple phototrophs, the RC-bound multi-heme Cyt subunits are classified into two groups based on whether or not a lipoprotein signal peptide followed by a cysteine (Cys) residue exists in its N-terminus9,10. The Cys-containing Cyt subunits with known structures all have their N-termini posttranslationally truncated (about 20 residues removed) at this Cys residue and are then modified by a covalently bound tri- or diacylglycerol lipid; such is characteristic of Blastochloris (Blc.) viridis11,12, Thermochromatium (Tch.) tepidum13, Thiorhodovibrio (Trv.) strain 97014, and Allochromatium (Alc.) tepidum15. Since the N-terminus-truncated Cyt subunits have no transmembrane domain, they are anchored by the covalently bound lipid in the membrane-embedded RC. By contrast, a Cys-lacking Cyt subunit from the aerobic purple bacterium Roseobacter (Rsb.) denitrificans has a full N-terminus unmodified in the purified RC10, and based on its sequence, presumably forms an α-helix inserted in the membrane as an anchor instead of lipid16. Predictions based on sequence analyses also suggest that this may be the case in Rhodovulum (Rdv.) sulfidophilum, Acidiphilium (Acp.) rubrum, and Rhodospirillum (Rsp.) molischianum10.
In contrast to these, Rhodobacter (Rba.) species lack an RC-bound Cyt subunit and the gene that encodes it (pufC) in their puf operon. Instead, these species use both a soluble Cyt c217 and a membrane-anchored mono-heme Cyt cy4 to facilitate electron transfer to the RC. In all Rhodobacter species, there is a unique protein in the light-harvesting 1–reaction center (LH1–RC) complex called PufX that interacts with the RC-L subunit and LH1 polypeptides18,19,20,21,22,23. PufX (encoded by the gene pufX24,25) is a transmembrane protein composed of about 80 amino acids, and pufX is located at the same position as pufC—immediately following pufM (the gene encoding the RC-M subunit) in the puf operon. PufX plays important roles in the regulation of cyclic electron transfer and photosynthetic membrane morphology and core complex organization26, and its transmembrane portion has structural motifs similar to the N-terminal (and likely transmembrane) helix of the Rsb. denitrificans Cyt subunit10,16. These findings have led to a hypothesis that PufX has phylogenetic roots in the N-terminal domain of the RC-bound Cyt subunit10.
Here, we lend experimental support for this sequence-based hypothesis with structural data from a cryo-EM analysis of the LH1–RC complex from the phylogenetically and physiologically unique purple phototrophic bacterium Rhodopila (Rpi.) globiformis27. This phototroph inhabits warm acidic springs at pH values as low as 3 and is the most acidophilic of all known anaerobic purple phototrophs28. The Rpi. globiformis RC complex contains a full-length Cyt subunit whose structure confirms that its N-terminal region forms a transmembrane helix within the LH1–RC complex. The N-terminal portion of the Cyt subunit also displays a marked similarity to PufX, providing crucial new insights into the relationship between the two proteins. Moreover, the structure of the bacteriochlorophyll (BChl) a-containing Rpi. globiformis LH1 complex revealed multiple copies of a polypeptide absent from the LH1 complexes of other BChl a-containing species but related to the γ-polypeptides in the LH1 complexes of BChl b-containing purple bacteria. Collectively, these data show the Rpi. globiformis LH1–RC complex to be structurally unique among purple bacteria and provide experimental evidence of a phylogenetic link between PufX and the RC Cyt subunit.
Results
Structural overview of the Rpi. globiformis LH1–RC
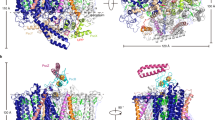
Purified LH1–RC from cells of Rpi. globiformis DSM161T exhibited an LH1 absorption maximum at 879 nm (Supplementary Fig. 1). The cryo-EM structure of the core complex was determined at 2.24 Å resolution (Table 1 and Supplementary Figs. 2–4). The LH1 complex forms a closed ring structure (Fig. 1a, b) with an arrangement of the α- and β-polypeptides similar to that of purple bacteria such as Tch. tepidum13, Trv. strain 97014, Rhodospirillum rubrum29,30 and Alc. tepidum15. In addition to the 16 pairs of αβ-polypeptides, several fragments of electron potential densities with similar length and α-helical characteristic were aligned in the transmembrane region between β-polypeptides in the refined map. These were assumed to be the same protein but could not be modeled by any annotated protein sequences. However, because of the high quality of our density map, we were able to trace most residues of this unknown protein. The derived amino acid sequence allowed us to search the Rpi. globiformis DSM 161T genome31 using TBLASTN32, and in so doing, a 69 bp nucleotide sequence encoding 22 amino acids (NCBI Reference Sequence: NZ_NHRY01000272.1) (Supplementary Fig. 5) emerged whose translated amino acid sequence precisely fit the density map (Supplementary Fig. 4b). Hereafter, we refer to this previously unannotated protein as “γ-like polypeptide”, a structural analogue to the LH1 γ-polypeptides found thus far only in BChl b-containing LH1 complexes33,34,35,36; a total of 11 γ-like polypeptides were present per Rpi. globiformis LH1 complex (Fig. 1c, d).

a Side view of the core complex parallel to the membrane plane. b Top view from the periplasmic side of the membrane. Dotted ovals indicate the positions that lack the γ-like polypeptide. c Side view of surface representation of the complex. Dotted red ovals indicate the positions that lack the γ-like polypeptide. d Tilted view showing distributions of the γ-like polypeptides (magenta cartoons) and cardiolipins (red sticks) in the LH1 ring. e Side view of the Cyt subunit (cyan) in the LH1–RC showing its N-terminal transmembrane domain. Heme groups are shown by paleyellow sticks. Color scheme: LH1-α, green; LH1-β, slate blue; LH1-γ, magenta; RC-C, cyan; RC-L, wheat; RC-M, blue; BChl a, red sticks; carotenoids, yellow sticks; BPhe a, magenta sticks; lipids or detergents, gray.
The Rpi. globiformis RC complex also contained a highly unusual Cyt subunit with its full N-terminal domain present and embedded in the transmembrane region forming an α-helix as the membrane anchor (Fig. 1e). This arrangement differs from that in all other purple bacterial RC-bound Cyt subunits with known structures; in the latter, the N-termini are truncated and modified by acyl lipids that function as membrane anchors. The intact (untruncated) Cyt structure in Rpi. globiformis may therefore be an ancestral form of the RC-bound Cyt from which more compact (truncated) versions evolved.
The Rpi. globiformis LH1–RC contains 36 molecules of BChls a (32 in the LH1 and 4 in the RC) (Supplementary Fig. 15, Supplementary Table 1) and 17 keto-carotenoids (16 trans-carotenoids in LH1 and 1 cis-carotenoid in RC), the latter pigments are unique to this species27,37. Four types of keto-carotenoids were reported in the Rpi. globiformis membranes37, and were confirmed in our work (Supplementary Fig. 6). Because the R.g. keto-II type carotenoid is dominant in the purified LH1–RC (Supplementary Fig. 6), it was modeled in the LH1 structure with its keto group located on the periplasmic side. Three types of quinones, ubiquinone (UQ), menaquinone (MQ) and rhodoquinone, are present in Rpi. globiformis membranes, with UQ predominating (Supplementary Fig. 7). Based on the density map, one MQ and one UQ were identified at the QA and QB sites in the RC (Supplementary Fig. 4b, Supplementary Fig. 15), respectively. Electron potential density corresponding to a disulfide bond (Cys170–Cys172) in the RC-M subunit on the periplasmic surface is clearly visible in Supplementary Fig. 4b, indicating that our cryo-EM map was free from electron damage38.
The Cyt subunit in the Rpi. globiformis RC
The Rpi. globiformis Cyt subunit contains fewer charged residues (30 and 24 of basic and acidic residues, respectively) than that of the neutrophilic Blc. viridis (39 and 28 of basic and acidic residues, respectively), a tendency also displayed by the extremely acidophilic aerobic purple bacterium Acp. rubrum39. Moreover, the surface of the membrane-extruded portion of the Rpi. globiformis Cyt subunit is slightly more basic than the Cyt subunits of Blc. viridis and Tch. tepidum (Fig. 2a, Supplementary Fig. 8). This is likely a function of the acidophilic nature of Rpi. globiformis27 and is consistent with the observation that lowering pH results in a relatively more basic surface of the RC-bound Cyt subunit40. The purified Rpi. globiformis LH1–RC also showed some degree of thermostability over a pH range of 5.0–7.5 and a slightly lower stability at higher pH (Supplementary Fig. 8c); in nature, Rpi. globiformis inhabits warm acidic springs near 40 °C27,28. The overall structure of the Rpi. globiformis Cyt subunit including the four heme groups is similar to that of the Blc. viridis12,41 except, and importantly, for the N-terminal domains (Fig. 2b). The N-terminus of the Rpi. globiformis Cyt subunit is extended into the transmembrane region and forms an α-helix anchor in the membrane; by contrast, the Blc. viridis Cyt subunit is truncated at Cys21 and contains a diacylglycerol as the membrane anchor11. The latter structure is also found in the RC-bound Cyt subunits from Tch. tepidum13, Trv. strain 97014 and Alc. tepidum15. The transmembrane domain of the Rpi. globiformis Cyt subunit is surrounded by phospholipids and detergents on the periplasmic surface (Fig. 2d), implying a propensity of this portion for lipid molecules. The N-terminus of the Cyt subunit from Asn2 to Ala8 interacts with the RC-M and RC-H proteins and a nearby LH1 α-polypeptide on the cytoplasmic surface (Fig. 2e), mainly through charge/polar and hydrogen bond interactions. In summary, the extensive interactions of the Rpi. globiformis Cyt subunit transmembrane N-terminal domain with surrounding proteins and lipids not only make the subunit structurally unique among anaerobic purple bacteria but firmly anchor the Cyt subunit and thus stabilize the entire LH1–RC complex.

a Surface charge distributions of the Cyt subunit with color codes according to the electrostatic potential from –5.0 kBT (red, negative charge) to +5.0 kBT (blue, positive charge) in the scale bars. Heme-1 is shown by green sticks. b Superposition of the Cα carbons of the Cyt subunits between Rpi. globiformis (cyan) and Blc. viridis (PDB: 3T6E, gray). The diacylglycerol group attached to the N-terminal Cys21 in Blc. viridis is shown by red sticks. Heme groups in the Rpi. globiformis Cyt subunit is shown by pale yellow sticks. c Superposition of the Cα carbons of the Cyt subunits of Rpi. globiformis (cyan) and Rof. castenholzii (PDB: 5YQ7, black). d The Cyt subunit (cyan cartoon) in the Rpi. globiformis RC showing its N-terminal domain surrounded by phosphatidylglycerol (magenta sticks) and detergent (DDM, green sticks) molecules on the periplasmic surface. e Expanded view of the N-terminus of the Cyt subunit marked in d showing interactions with the surrounding M subunit (blue), H-subunit (salmon) and LH1 α-polypeptide (green). Dashed lines indicate several representative distances shorter than 4.0 Å.
Comparison of the Rpi. globiformis Cyt subunit with PufX in Rhodobacter species
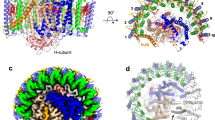
Because the structure of the Rpi. globiformis LH1–RC determined here is the first in an anaerobic purple bacterium containing an intact RC-bound Cyt with a full N-terminus, we were able to test a hypothesis surrounding the relationship between the N-terminal portion of the intact Cyt and the PufX polypeptide in the LH1–RC from Rhodobacter species (Supplementary Fig. 9)10. Superposition of the RC-M proteins from Rpi. globiformis and Rba. sphaeroides revealed that the C-terminus (Leu55–Gly69) of PufX is indeed closely located and parallel to a loop region (Phe27–Arg40) of the Cyt subunit immediately following its N-terminal transmembrane domain (Fig. 3a, b, f). The positional and conformational overlaps between the two fragments along with a similar topology of their transmembrane helices are structurally remarkable features and support the contention that PufX from Rhodobacter species may have phylogenetic roots in the intact N-terminus of the Cyt subunit.

a Superposition of the Cα carbons of the RC-M subunits between the Rba. sphaeroides monomeric LH1–RC (PDB: 7F0L) and Rpi. globiformis RC (this work). PufX is shown as orange cartoon. The transmembrane helix and the following loop region of the Cyt subunit are shown by cyan cartoon and green sticks (main chain only), respectively. The other portion of the Cyt subunit is shown by a black ribbon and heme groups are shown by pale yellow sticks. Protein-U in Rba. sphaeroides LH1–RC is shown by magenta ribbon. b Expanded view marked in a showing that the C-terminus (Leu55–Gly69) of PufX overlaps positionally and conformationally with the N-terminal loop region (Phe27–Arg40, colored green) of the Rpi. globiformis Cyt subunit. c Similar comparison between the C-terminus of Rba. sphaeroides PufX and the N-terminus (Cys21–Phe34, colored green) of Blc. viridis Cyt subunit (PDB: 3T6E). d Similar comparison between the C-terminus of Rba. sphaeroides PufX and the N-terminus (Cys23–Tyr35, colored green) of Tch. tepidum Cyt subunit (PDB: 5Y5S). e Comparison of the Rba. sphaeroides PufX with the N-terminal transmembrane domain (Arg17–Ala49, red cartoon) of Rof. castenholzii Cyt subunit in an LH–RC structure (PDB: 5YQ7). A hypothetical subunit X is shown by magenta cartoon. f Sequence alignments of Rba. sphaeroides PufX (from strain IL106) with the N-termini of the Cyt subunits from other phototrophs based on the residues in the loop regions marked by dashed box or the transmembrane domains. Residues in the C-terminus (Leu55–Gly69) of PufX are shown by orange fonts. Residues truncated by posttranslational modification are shown by gray fonts. The Cys residues that bind a triacyl or diacyl group are shown by red fonts. Residues with green fonts correspond to the N-terminal portions shown in a–d. Transmembrane regions are underlined.
Further evidence for a structural relationship between the C-terminus of PufX and the N-terminus of the RC-bound Cyt subunit can be observed for the Cyt subunits with truncated N-termini (Fig. 3, Supplementary Fig. 10). The N-terminal stretches of Cyt subunits containing about 14 residues after posttranslational truncation from Blc. viridis, Tch. tepidum, Trv. strain 970 and Alc. tepidum all closely overlap with the C-terminal domain (Leu55–Gly69) of PufX on the periplasmic surface, indicating that these portions are conserved in both proteins (Fig. 3c, d, f, Supplementary Fig. 10). It is of interest to note that an RC-bound Cyt subunit from the phylogenetically distinct ancient filamentous anoxygenic phototroph Roseiflexus (Rof.) castenholzii also has an intact N-terminal domain3 (Fig. 2c), but its N-terminal portion (transmembrane helix and the following loop region) is located at a slightly different position from that of PufX (Fig. 3e). An additional hypothetical subunit X (63 residues) in Rof. castenholzii was found near to where PufX is positioned in Rhodobacter species3.
Anchoring mechanism of the Rpi. globiformis Cyt subunit to the LH1–RC
In addition to anchoring the Rpi. globiformis Cyt subunit by its N-terminal transmembrane domain, four LH1 α-polypeptides tightly associate the soluble portion of the Cyt subunit with their C-termini in a manner similar to that seen with the α3-polypeptide (the longest chain) in the Alc. tepidum LH115 and the α2- and α4-polypeptides (also the longest chains) in the Trv. strain 970 LH114 (Fig. 4a). In fact, one Rpi. globiformis α-polypeptide (chain ID:3) structurally overlaps with the Alc. tepidum α3-polypeptide and the Trv. strain 970 α2-polypeptide at their C-termini, and another (chain ID: S) overlaps with the Trv. strain 970 α4-polypeptide. Although overall sequence comparison of the Rpi. globiformis α-polypeptide with these polypeptides shows relatively low similarities for their C-termini, local alignment reveals a key motif [A/S]-[P/A]-[L/M/I]-P (Fig. 4b, Supplementary Fig. 11) at their distal C-terminal ends that is likely responsible for the extensive interactions with their corresponding Cyt subunits. The Rpi. globiformis Cyt subunit also has a larger surface contact area (332.1 Å2) with its LH1–RC than do the other LH1–RC (159.6 Å2 for Trv. strain 970 and 138.4 Å2 for Alc. tepidum). In addition, despite low sequence similarities, the RC-M subunits interact extensively with the Cyt subunits at their C-termini with similar structural features for all Cyt subunit-containing species (Supplementary Fig. 12). These data strongly suggest that the interacting residues between the two subunits evolved simultaneously to form suitable surface recognition contacts.

a Left panel shows a top view of interactions from the periplasmic side between the Rpi. globiformis α-polypeptides (chain 3, orange; chain S, red; chain U, yellow; chain W, turquoise blue) and the Cyt subunit (transparent cyan); arrow heads indicate the overlapping C-terminal regions between the Rpi. globiformis α-polypeptides and the Alc. tepidum α3-polypeptide (PDB: 7VRJ, gray) and the α2- or α4-polypeptide of Trv. strain 970 (PDB: 7C9R, black). Other color scheme: LH1-α, green; LH1-β, slate blue. Right panel is a side view showing the four Rpi. globiformis α-polypeptides (colored) interacting with the Cyt subunit. For comparison, the Alc. tepidum α3-polypeptide and the α2- or α4-polypeptide of Trv. strain 970 are shown by gray and black cartoons, respectively. b Alignment of the C-terminal amino acid sequences of the α-polypeptides between Rpi. globiformis, Alc. tepidum and Trv. strain 970. Bold letters indicate residues in the overlapping regions of the structures. Symbol scheme: identical (*), conservative (:), semi-conservative (.). Upper colored bars represent regions of the Rpi. globiformis α-polypeptides that interact with the Cyt subunit. The color code is the same as in a.
The novel γ-like polypeptides in the Rpi. globiformis LH1 complex
The newly identified γ-like polypeptide from our cryo-EM density map of the purified Rpi. globiformis LH1–RC was confirmed by mass spectroscopy; the novel polypeptide contained 22 residues with its N-terminal Met modified by a formyl group (Supplementary Fig. 13). The amino acid sequence exhibited high hydrophobicity, in good agreement with prediction by a membrane protein topology analysis42 that the γ-like polypeptide is an integral membrane protein (Supplementary Fig. 5c). Eleven γ-like polypeptides are located between β-polypeptides in the LH1–RC with an opposite orientation (N-terminus on the periplasmic side) to the α-and β-polypeptides (Fig. 5a); the five remaining sites were occupied by cardiolipin molecules (Fig. 1d, Fig. 5b). Inspection of the asymmetrically distributed binding sites of the γ-like polypeptides revealed a high structural homogeneity in their binding environments with nearby β-polypeptides (Supplementary Fig. 14b). By contrast, the cardiolipin-binding sites exhibited relatively large structural inhomogeneities (Supplementary Fig. 14b), indicating specific bindings of the γ-like polypeptide and cardiolipin to these sites.

a Density map of a typical γ-like polypeptide with its nearby LH1 β-polypeptides showing the opposite orientations. The density map is shown at a contour level of 4.0 σ. b Density map of a cardiolipin with its nearby LH1 β-polypeptides at the location that lacks a γ-like polypeptide. c Interactions between a γ-like polypeptide (magenta) with its neighboring LH1 β-polypeptides (slate blue) with the dashed lines representing typical close contacts (<4.0 Å) between the two polypeptides. A BChl a molecule is shown by red sticks. d Side view of a surface representation showing a channel (white dashed circle) in the LH1 ring which lacks a γ-like polypeptide, other channels are sealed by the γ-like polypeptides (magenta). Color scheme: LH1-α, green; LH1-β, slate blue; BChl a, red; carotenoids, yellow. e Superposition of the LH1 polypeptides between Rpi. globiformis (colored) and Blc. viridis (PDB: 6ET5, gray) showing the structural similarity between the γ-polypeptides (cartoon). f Alignment of the γ-polypeptides between Rpi. globiformis and BChl b-containing species with identical amino acids (*), conservative (:) and semi-conservative (.). The N-terminal fragments in Blastochloris species shown by gray fonts were not identified in the purified protein. Hlr: Halorhodospira.
The γ-like polypeptides interact over the entire range with neighboring β-polypeptides, mainly through hydrophobic interactions (Fig. 5c), enhancing stability of the LH1 ring. Moreover, the γ-like polypeptides block 11 of the 16 pores formed between LH1-αβ pairs that presumably function as quinone transport channels (Fig. 5d); one other channel is blocked by an additional UQ and a carotenoid (Supplementary Fig. 15b, c) leaving four channels in an open state. This incomplete blockage of the circular LH1 “fence” may be a mechanism for regulating the rate of quinone transport (Supplementary Fig. 16). Also, because the one channel blocked by UQ and a carotenoid is near the QA site, the route for quinone access is limited to the QB site. Such a partial blockage arrangement implies a mechanism for quinone exchange only through the QB site.
The γ-like polypeptide in the Rpi. globiformis LH1 complex is the first (and thus far only) example of such a protein in BChl a-containing purple phototrophs. The polypeptides are positioned and show a similar conformation and orientation as do the LH1 γ-polypeptides in the BChl b-containing bacterium Blc. viridis35 (Fig. 5e). However, differing from Blc. viridis, the Rpi. globiformis γ-like polypeptides do not interact with LH1 α-polypeptides because the Rpi. globiformis α-polypeptide has a much shorter N-terminus than that of Blc. viridis. Although comparison of amino acid sequences over the transmembrane domain revealed that the full-length Rpi. globiformis γ-polypeptide has relatively low sequence similarities to those of the expressed γ-polypeptides from BChl b-containing species33,34,35,36,43 (Fig. 5f), this could indicate that the Rpi. globiformis γ-like polypeptide is a new type of LH1 γ-polypeptide, perhaps one functionally best suited to the warm acidic conditions in which this unusual phototroph thrives.
Discussion
Photosynthetic reaction centers bound with a multi-heme Cyt subunit are common in phototrophic purple bacteria (Proteobacteria)1, and those with known structures all contain a truncated N-terminus and are modified by a lipid. By contrast, the structure of the LH1–RC from Rpi. globiformis (α1-subgroup in the Proteobacteria44,45) solved in the present work shows that its RC Cyt subunit is unique; it has an intact N-terminus that forms an α-helix in the photosynthetic membrane (Fig. 1e). This implies that a different strategy for membrane-anchoring of the Cyt subunit is necessary in this phototroph. Comparison of the N-terminal loop region (following the transmembrane helix) of the Rpi. globiformis Cyt with the C-terminal portion of PufX from Rhodobacter species (α3-subgroup) revealed remarkable structural similarities in terms of location within the complex and conformation including the topology of the transmembrane domain. Hence, our structural work here supports partly the hypothesis10 that the phylogenetic roots of PufX lie in the N-terminus of the RC-bound Cyt subunit, furthermore suggesting that a common ancestor of phototrophic purple bacteria contained a full-length Cyt subunit that evolved independently in certain species through partial deletions of the pufC gene. Such deletions would have had at least two significant outcomes. First, the major portion of the Cyt subunit was cleft, generating an N-terminal fragment (corresponding to PufX) in Rhodobacter species10 that evolved as a distinct protein to structurally organize the core complex. And second, in species lacking PufX, the major portion of the Cyt subunit that contains heme groups remained and was subsequently modified by an acyl lipid at its truncated Cys residue as a membrane anchor. These species include those with structures, such as Blc. viridis12 (α2-subgroup), Tch. tepidum13 (γ-Proteobacteria) and Trv. strain 97014 (γ-Proteobacteria), and probably those with structures predicted from their sequences, such as Rubrivivax (Rvi.) gelatinosus (β-Proteobacteria) and Alc. vinosum (γ-Proteobacteria). Therefore, we hypothesize that the full-length Rpi. globiformis RC Cyt subunit represents a more ancient form than the lipid-anchored forms and more firmly associates with the RC to ensure efficient electron transfer.
The transmembrane domain of the Rpi. globiformis Cyt subunit is located close to the position occupied by protein-U in the Rba. sphaeroides LH1–RC (Fig. 3a), possibly as a consequence of spatial restriction. Because the interior of the LH1–RC is crowded, there is little space to accommodate a transmembrane domain, especially in closed-type LH1–RCs such as that of Rpi. globiformis. On the other hand, PufX is located on one edge of the open-shaped LH1 ring18,19,20,21,22,23. Occupation of this position by PufX prevents LH1 from forming a closed ring. This may be the reason why all known PufX-containing core complexes have an open LH1 ring. As a result, for the closed Rpi. globiformis LH1–RC that lacks protein-U, the position corresponding to protein-U and its immediate surroundings becomes the ideal candidate for accommodating the transmembrane domain of the Rpi. globiformis Cyt subunit. This is also applicable for the closed core complex of the filamentous green bacterium Rof. castenholzii. Similar to the Rpi. globiformis, Rof. castenholzii also lacks protein-U and its position is occupied by the N-terminal transmembrane domain of its Cyt subunit (Fig. 3e).
Inspection of the truncated N-terminal fragments of the Cyt subunits from Trv. strain 970, Alc. tepidum (Supplementary Fig. 10), Blc. viridis and Tch. tepidum (Supplementary Fig. 17a, b) showed hydrophobic signatures over a length of the membrane thickness, revealing their nature as transmembrane components. Modification of the Cys residues of these species by a lipid molecule at the truncated site coupled with the fact that the transmembrane domain of the Rpi. globiformis Cyt subunit is surrounded by phospholipid molecules at a similar position on the periplasmic surface (Fig. 2d) reflect a strong propensity of the truncation site to associate with lipid molecules. The lipid-anchored Cyt subunits likely have a weaker association with the RC than that of Rpi. globiformis because some of them can be dissociated from the RC with detergent treatments as demonstrated in Tch. tepidum46 and Rvi. gelatinosus47.
Although no structural information was presented, an RC-bound multi-heme Cyt subunit was identified in the purified RC from the aerobic purple photosynthetic bacterium Rsb. denitrificans (α3-subgroup)10,16, a species unable to carry out photosynthesis anaerobically. Resembling the Rpi. globiformis RC Cyt subunit, the subunit from this phototroph also has an intact N-terminus that was predicted from its sequence to form a transmembrane helix (Fig. 3f, Supplementary Fig. 17c). Similar cases have been inferred from sequence analyses of Rhodovulum sp.25, Chloroflexus (Cfl.) aurantiacus (Cyt c554)2 and from actual structural studies of the Cyt subunit from Rof. castenholzii3 (Fig. 3e); the latter two species are filamentous bacteria that emerge as the earliest phototrophs on a phylogenetic tree of 16 S rRNA genes44.
Discovery of the γ-like polypeptides in the Rpi. globiformis LH1 was quite unexpected, since these have been found only in BChl b-containing purple phototrophs and used for distinguishing BChl a- and BChl b-containing species. The Rpi. globiformis γ-like polypeptide is the shortest among those reported, containing only 22 residues and showed relatively low sequence similarities to γ-polypeptides from BChl b-containing purple phototrophs (Fig. 5f). Its small size and rather distant relationship to γ-polypeptides from BChl b-containing species likely explain why the gene encoding the Rpi. globiformis γ-like polypeptide was not annotated as a hypothetical protein. This gene encoding the Rpi. globiformis γ-like polypeptide is located at a considerable distance from pufLMC gene clusters and is flanked by genes encoding a hypothetical protein and a methyl-accepting chemotaxis protein (Supplementary Fig. 5b). By contrast, genes encoding the Blc. viridis γ-polypeptides are clustered in a region flanked by two genes encoding hypothetical proteins. These differences in gene synteny between the two species further support the conclusion that the Rpi. globiformis γ-like polypeptide is a distinct form of LH1 γ-polypeptides. Based on their structural characteristics and positions in the LH1–RC, the Rpi. globiformis γ-like polypeptides likely play roles in both stabilizing the LH1 ring structure and regulating quinone transport between the RC and the acidic conditions of the quinone pool in the periplasm. In addition to the γ-like polypeptides, the presence of 3-methylhopanoids in Rpi. globiformis48, lipids that help stabilize membranes and which are absent from most other anoxygenic phototrophs, is further evidence that anoxygenic photosynthesis in the warm acidic conditions of the Rpi. globiformis habitat may require special membrane components.
In summary, the structure of the Rpi. globiformis LH1–RC is a heretofore unseen version of this key photocomplex. The combination of an RC Cyt subunit anchored in the photosynthetic membrane by a polypeptide rather than an acyl lipid tail along with γ-like polypeptides and novel carotenoids makes the photocomplex of Rpi. globiformis structurally unique among all known purple bacteria. Moreover, along with the acidic, anoxic, and sulfidic warm-springs habitat of Rpi. globiformis—likely a common geological feature 3.4 billion years ago when photosynthesis first evolved—one could also speculate that the Rpi. globiformis LH1–RC contains the phylogenetic roots of PufX of Rhodobacter species and the γ-polypeptide of BChl b-containing purple bacterial species.
Methods
Preparation and characterization of the Rpi. globiformis LH1–RC complex
Cells of Rpi. globiformis strain 7950 (DSM 161T) were cultivated phototrophically (anoxic/light) at pH 5.1 and 25 °C for 7 days under incandescent light (60 W). Chromatophores were first treated with 0.30% (w/v) lauryldimethylamine N-oxide (LDAO) in 20 mM Tris-HCl buffer (pH 7.5) at 25 °C for 60 min to remove excess LH2, followed by centrifugation at 4 °C and 150,000 × g for 90 min. The pellets were then resuspended in the same buffer and extracted with 1.0% (w/v) n-dodecyl β-D-maltopyranoside (DDM) at 25 °C for 60 min followed by centrifugation at 4 °C and 150,000 × g for 90 min. The extracts were loaded onto sucrose gradient density tubes (five-stepwise concentrations: 10, 17.5, 25, 32.5, and 40% w/v) in 20 mM Tris-HCl buffer (pH 7.5) containing 0.05% w/v DDM followed by centrifugation at 4 °C and 150,000 × g for 6 h. The LH1–RC fractions were concentrated for absorption and circular dichroism (CD) measurements (Supplementary Fig. 1) and assessed by negative-stain EM using a JEM-1010 instrument (JEOL) (Supplementary Fig. 2a). Masses of the LH1 polypeptides were measured by matrix-assisted laser desorption/ionization time-of-flight mass spectroscopy (MALDI-TOF/MS)49. Carotenoids were analyzed by HPLC50. Suspension or solutions of membranes, LH1–RC and LH2 were directly injected into the HPLC system (Waters), which consisted of a μBondapak C18 column and a photodiode array detector (Shimazdu). A linear gradient from methanol:water (9:1, v/v) to methanol was applied for 20 min and then methanol at flow-rate of 1.8 ml/min. Each carotenoid was also identified by their molecular masses (Supplementary Fig. 6). Quinones were extracted from chromatophores and analyzed using reverse-phase HPLC51. Phospholipids extracted from chromatophores were analyzed using 31P-NMR52.
Cryo-EM data collection
Proteins for cryo-EM were concentrated to ~5 mg/ml. Then 2.5 microliters of the protein solution were applied on glow-discharged holey carbon grids (200 mesh Quantifoil R2/2 molybdenum) that had been treated with H2 and O2 mixtures in a Solarus II plasma cleaner (Gatan, Pleasanton, USA) for 30 s and then blotted, and plunged into liquid ethane at –182 °C using an EM GP2 plunger (Leica, Microsystems, Vienna, Austria). The applied parameters were a blotting time of 5 s at 80% humidity and 4 °C. Data were collected on a Titan Krios (Thermo Fisher Scientific, Hillsboro, USA) electron microscope at 300 kV equipped with a Falcon 3 camera (Thermo Fisher Scientific) (Supplementary Fig. 2b). Movies were recorded using EPU software (Thermo Fisher Scientific) at a nominal magnification of 96 k in counting mode and a pixel size of 0.820 Å at the specimen level with a dose rate of 0.69 e- per physical pixel per second, corresponding to 1.03 e- per Å2 per second at the specimen level. The exposure time was 38.9 s, resulting in an accumulated dose of 40 e- per Å2. Each movie includes 40 fractioned frames.
Image processing
All of the stacked frames were subjected to motion correction with MotionCor253. Defocus was estimated using CTFFIND454. A total of 421,464 particles were selected from 2,989 micrographs using the crYOLO55 (Supplementary Fig. 3). All of the picked particles were further analyzed with RELION3.156, and 211,382 particles were selected by 2-D classification and divided into four classes by 3-D classification resulting in only one good class containing 128,119 particles. The initial 3-D model was generated in RELION. The 3-D auto refinement without any imposed symmetry (C1) produced a map at 2.35 Å resolution after contrast transfer function refinement, Bayesian polishing, masking, and post-processing. These particle projections were then subjected to subtraction of the detergent micelle density followed by 3-D auto refinement to yield the final map with a resolution of 2.24 Å according to the gold-standard Fourier shell correlation using a criterion of 0.143 (Supplementary Fig. 3b)57. The local resolution maps were calculated on RESMAP58.
Model building and refinement of the LH1–RC complex
The initial atomic model for the LH1–RC structures was generated with MODELLER v10.159 using the atomic model of the Tch. tepidum LH1–RC (PDB: 5Y5S). The model was then fitted to the cryo-EM map obtained for the Rpi. globiformis LH1–RC using Chimera60. Amino acid substitutions and real-space refinement for the peptides and cofactors were performed using COOT61. Whole regions of the LH1 γ-like polypeptides as well as both terminal regions of the Cyt subunit and LH1 α-subunit were modeled ab-initio based on the density. The manually modified model was refined in real-space on PHENIX62, and the COOT/PHENIX refinement was iterated until the refinements converged. Finally, the statistics calculated using MolProbity63 were checked. The coordinates and restraints for carotenoid R.g. keto-II were generated using AceDRG64 in the CCP4 suite65. Figures were drawn with the Pymol Molecular Graphic System (Ver2.5, Schrödinger)66 and UCSF Chimera60. Surface charge distributions were calculated using the APBS Electrostatics plugin in the Pymol.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Map and model have been deposited in the EMDB and PDB with the accession codes: EMD-33501 and PDB-7XXF. All other data are available from the authors upon reasonable request.
References
Nitschke, W. & Dracheva, S. M. In Anoxygenic Photosynthetic Bacteria (eds. Blankenship, R. E., Madigan, M. T. & Bauer, C. E.) 775–805 (Kluwer Academic Publishers, 1995).
Dracheva, S., Williams, J. C., van Driessche, G., van Beeumen, J. J. & Blankenship, R. E. The primary structure of cytochrome c-554 from the green photosynthetic bacterium Chloroflexus aurantiacus. Biochemistry 30, 11451–11458 (1991).
Xin, Y. et al. Cryo-EM structure of the RC-LH core complex from an early branching photosynthetic prokaryote. Nat. Commun. 9, 1568 (2018).
Daldal, F., Deshmukh, M. & Prince, R. C. Membrane-anchored cytochrome c as an electron carrier in photosynthesis and respiration: past, present and future of an unexpected discovery. Photosynth. Res. 76, 127–134 (2003).
Azai, C., Tsukatani, Y., Itoh, S. & Oh-oka, H. C-type cytochromes in the photosynthetic electron transfer pathways in green sulfur bacteria and heliobacteria. Photosynth. Res. 104, 189–199 (2010).
Hirano, Y., Higuchi, M., Oh-oka, H., Miki, K. & Wang, Z.-Y. Crystal structure of the electron carrier domain of the reaction center cytochrome cz subunit from green photosynthetic bacterium Chlorobium tepidum. J. Mol. Biol. 397, 1175–1187 (2010).
Abresch, E. C. et al. Characterization of highly purified, fully active, crystallizable RC-LH1-PufX core complex from Rhodobacter sphaeroides. Photosynth. Res. 86, 61–70 (2005).
Matsuura, K. & Shimada, K. In Current Research in Photosynthesis, Vol. 1 (ed. Baltscheffsky, M.) 193–193 (Kluwer Academic Publishers, 1990).
Hayashi, S. & Wu, H. C. Lipoproteins in bacteria. J. Bioenerg. Biomembr. 22, 451–471 (1990).
Huck, O., Schiltz, E., Drews, G. & Labahn, A. Sequence analysis reveals new membrane anchor of reaction centre-bound cytochromes possibly related to PufX. FEBS Lett. 535, 166–170 (2003).
Weyer, K. A., Schäfer, W., Lottspeich, F. & Michel, H. The cytochrome subunit of the photosynthetic reaction center from Rhodopseudomonas viridis is a lipoprotein. Biochemistry 26, 2909–2914 (1987).
Roszak, A. W. et al. New insights into the structure of the reaction centre from Blastochloris viridis: evolution in the laboratory. Biochem. J. 442, 27–37 (2012).
Yu, L.-J., Suga, M., Wang-Otomo, Z.-Y. & Shen, J.-R. Structure of photosynthetic LH1-RC supercomplex at 1.9 Å resolution. Nature 556, 209–213 (2018).
Tani, K. et al. Cryo-EM structure of a Ca2+-bound photosynthetic LH1-RC complex containing multiple αβ-polypeptides. Nat. Commun. 11, 4955 (2020).
Tani, K. et al. A Ca2+-binding motif underlies the unusual properties of certain photosynthetic bacterial core light–harvesting complexes. J. Biol. Chem. 298, 101967 (2022).
Kortlüke, C., Breese, K., Gad’on, N., Labahn, A. & Drews, G. Structure of the puf operon of the obligately aerobic, bacteriochlorophyll a-containing bacterium Roseobacter denitrificans OCh114 and its expression in a Rhodobacter capsulatus puf puc deletion mutant. J. Bacteriol. 179, 5247–5258 (1997).
Axelrod, H. L. & Okamura, M. Y. The structure and function of the cytochrome c2: reaction center electron transfer complex from Rhodobacter sphaeroides. Photosynth. Res. 85, 101–114 (2005).
Bracun, L. et al. Cryo-EM structure of the photosynthetic RC-LH1-PufX supercomplex at 2.8-Å resolution. Sci. Adv. 7, eabf8864 (2021).
Tani, K. et al. A previously unrecognized membrane protein in the Rhodobacter sphaeroides LH1-RC photocomplex. Nat. Commun. 12, 6300 (2021).
Qian, P. et al. Cryo-EM structure of the monomeric Rhodobacter sphaeroides RC-LH1 core complex at 2.5 Å. Biochem. J. 478, 3775–3790 (2021).
Qian, P. et al. Cryo-EM structure of the dimeric Rhodobacter sphaeroides RC-LH1 core complex at 2.9 Å: the structural basis for dimerisation. Biochem. J. 478, 3923–3937 (2021).
Tani, K. et al. Asymmetric structure of the native Rhodobacter sphaeroides dimeric LH1-RC complex. Nat. Commun. 13, 1904 (2022).
Cao, P. et al. Structural basis for the assembly and quinone transport mechanisms of the dimeric photosynthetic RC–LH1 supercomplex. Nat. Commun. 13, 1977 (2022).
Lee, J. K., DeHoff, B. S., Donohue, T. J., Gumport, R. I. & Kaplan, S. Transcriptional analysis of puf operon expression in Rhodobacter sphaeroides 2.4.1 and an intercistronic transcription terminator mutant. J. Biol. Chem. 264, 19354–19365 (1989).
Tsukatani, Y. et al. Phylogenetic distribution of unusual triheme to tetraheme cytochrome subunit in the reaction center complex of purple bacteria. Photosynth. Res. 79, 83–91 (2004).
Holden-Dye, K., Crouch, L. I. & Jones, M. R. Structure, function and interaction of the PufX protein. Biochim. Biophys. Acta Bioenerg. 613-630, 2008 (1777).
Pfennig, N. Rhodopseudomonas globiformis, sp. n., a new species of the Rhodospirillaceae. Arch. Microbiol. 100, 197–206 (1974).
Imhoff, J. F. & Madigan, M. T. Genus Rhodopila Imhoff, Trüper and Phennig 1984 341VP. In Bergey’s Manual of Systematics of Archaea and Bacteria (ed. Whitman, W. B.) (Wiley Online Library, 2021).
Tani, K. et al. Cryo-EM structure of the photosynthetic LH1-RC complex from Rhodospirillum rubrum. Biochemistry 60, 2483–2491 (2021).
Qian, P. et al. Cryo-EM structure of the Rhodospirillum rubrum RC-LH1 complex at 2.5 Å. Biochem. J. 478, 3253–3263 (2021).
Imhoff, J. F., Rahn, T., Künzel, S. & Neulinger, S. C. New insights into the metabolic potential of the phototrophic purple bacterium Rhodopila globiformis DSM 161T from its draft genome sequence and evidence for a vanadium-dependent nitrogenase. Arch. Microbiol. 200, 847–857 (2018).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Brunisholz, R. A., Jay, F., Suter, F. & Zuber, H. The light-harvesting polypeptides of Rhodopseudomonas viridis. Biol. Chem. Hoppe-Seyler 366, 87–98 (1985).
Zuber, H. & Cogdell, R. J. In Anoxygenic Photosynthetic Bacteria (eds. Blankenship, R. E., Madigan, M. T. & Bauer, C. E.) 315–348 (Kluwer Academic Publishers, 1995).
Qian, P., Siebert, C. A., Wang, P., Canniffe, D. P. & Hunter, C. N. Cryo-EM structure of the Blastochloris viridis LH1-RC complex at 2.9 Å. Nature 556, 203–208 (2018).
Seto, R. et al. Lycopene-family carotenoids confer thermostability on photocomplexes from a new thermophilic bacterium. Biochemistry 59, 2351–2358 (2020).
Schmidt, K. & Liaaen-Jensen, S. Bacterial carotenoids XLII New keto-carotenoids from Rhodopseudomonas globiformis (Rhodospirillaceae). Acta Chem. Scand. 27, 3040–3052 (1973).
Kato, K. et al. High-resolution cryo-EM structure of photosystem II reveals damage from high-dose electron beams. Commun. Biol. 4, 382 (2021).
Nagashima, K. V. P., Matsuura, K., Wakao, N., Hiraishi, A. & Shimada, K. Nucleotide sequences of gene coding for photosynthetic reaction centers and light–harvesting proteins of Acidiphilium rubrum and related aerobic acidophilic bacteria. Plant Cell Physiol. 38, 1249–1258 (1997).
Kawakami, T. et al. Crystal structure of a photosynthetic LH1-RC in complex with its electron donor HiPIP. Nat. Commun. 12, 1104 (2021).
Deisenhofer, J., Epp, O., Miki, K., Huber, R. & Michel, H. Structure of the protein subunits in the photosynthetic reaction centre of Rhodopseudomonas viridis at 3Å resolution. Nature 318, 618–624 (1985).
Krogh, A., Larsson, B., von Heijne, G. & Sonnhammer, E. L. L. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580 (2001).
Tsukatani, Y., Hirose, Y., Harada, J., Yonekawa, C. & Tamiaki, H. Unusual features in the photosynthetic machinery of Halorhodospira halochloris DSM 1059 revealed by complete genome sequencing. Photosynth. Res. 140, 311–319 (2019).
Woese, C. R. Bacterial evolution. Microbiol. Rev. 51, 221–271 (1987).
Nagashima, S. & Nagashima, K. V. P. Comparison of photosynthesis gene clusters retrieved from total genome sequences of purple bacteria. Adv. Bot. Res. 66, 151–178 (2013).
Nozawa, T. et al. Properties of the reaction center of the thermophilic purple photosynthetic bacterium Chromatium tepidum. Biochim. Biophys. Acta 894, 468–476 (1987).
Fukushima, A., Matsuura, K., Shimada, K. & Satoh, T. Reaction center-B870 pigment protein complexes with bound cytochromes c-555 and c-551 from Rhodocyclus gelatinosus. Biochim. Biophys. Acta 933, 399–405 (1988).
Mayer, M. H. et al. Anaerobic 3-methylhopanoid production by an acidophilic photosynthetic purple bacterium. Arch. Microbiol. 203, 6041–6052 (2021).
Sekine, F. et al. Gene sequencing and characterization of the light-harvesting complex 2 from thermophilic purple sulfur bacterium Thermochromatium tepidum. Photosynth. Res. 111, 9–18 (2012).
Takaichi, S. et al. Direct injection of pigment–protein complexes and membrane fragments suspended in water from phototrophs to C18 HPLC. Photosynth. Res. 144, 101–107 (2020).
Kimura, Y. et al. Characterization of the quinones in purple sulfur bacterium Thermochromatium tepidum. FEBS Lett. 589, 1761–1765 (2015).
Nagatsuma, S. et al. Phospholipid distributions in purple phototrophic bacteria and LH1-RC core complexes. Biochim. Biophys. Acta Bioenerg. 1860, 461–468 (2019).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Rohou, A. & Grigorieff, N. Fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Wagner, T. & Raunser, S. The evolution of SPHIRE-crYOLO particle picking and its application in automated cryo-EM processing workflows. Commun. Biol. 3, 61 (2020).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 7, e42166 (2018).
Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
Kucukelbir, A., Sigworth, F. J. & D, T. H. Quantifying the local resolution of cryo-EM density maps. Nat. Methods 11, 63–65 (2014).
Šali, A. & Blumdell, T. L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 234, 779–815 (1993).
Pettersen, E. F. et al. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Cryst. D66, 486–501 (2010).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D66, 213–221 (2010).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D. Biol. Crystallogr. 66, 12–21 (2010).
Long, F. et al. AceDRG: a stereochemical description generator for ligands. Acta Cryst. D73, 112–122 (2017).
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Cryst. D67, 235–242 (2011).
DeLano, W. L. The PyMOL molecular graphics system. (DeLano Scientific, LCC, 2004).
Acknowledgements
We thank C. Tanaka for providing technical assistance. This research was partially supported by Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research (BINDS)) from AMED under Grant Numbers JP20am0101118 (support number 1758) and JP20am0101116 (support number 1878), 17am0101116j0001, 18am0101116j0002, 19am0101116j0003, and 22am121004. R.K., M.H., and B.M.H. acknowledge the generous support of the Okinawa Institute of Science and Technology (OIST), Scientific Computing & Data Analysis Section at OIST and the Japanese Cabinet Office. L.-J.Y. acknowledges the financial supports from the National Key R&D Program of China (2021YFA0909600). This work was supported in part by JSPS KAKENHI Grant Numbers JP16H04174, JP18H05153, JP20H05086, and JP20H02856, Takeda Science Foundation, and the Kurata Memorial Hitachi Science and Technology Foundation, Japan.
Author information
Authors and Affiliations
Contributions
Z.-Y.W.-O. and K.T. designed the work, M.T.M. provided materials, K.T., R. K., K.K., S.T., and M.H. performed the experiments, K.T., R.K., K.V.P.N., L.-J.Y., Y.K., M.T.M., A.M., B.M.H., and Z.-Y.W.-O. analyzed data, Z.-Y.W.-O., K.T., and M.T.M. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Xialing Xu, James Allen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Eve Rogers. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tani, K., Kanno, R., Kurosawa, K. et al. An LH1–RC photocomplex from an extremophilic phototroph provides insight into origins of two photosynthesis proteins. Commun Biol 5, 1197 (2022). https://doi.org/10.1038/s42003-022-04174-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-022-04174-2
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
