
Abstract
As the nutrient quality changes, the fractions of ribosomal proteins in the proteome are usually positively correlated with the growth rates due to the auto-catalytic nature of ribosomes. While this growth law is observed across multiple organisms, the relation between the ribosome fraction and growth rate is often more complex than linear, beyond models assuming a constant translation speed. Here, we propose a general framework of protein synthesis considering heterogeneous translation speeds and protein degradations. We demonstrate that the growth law curves are generally environment-specific, e.g., depending on the correlation between the translation speeds and ribosome allocations among proteins. Our predictions of ribosome fractions agree quantitatively with data of Saccharomyces cerevisiae. Interestingly, we find that the growth law curve of Escherichia coli nevertheless appears universal, which we prove must exhibit an upward bending in slow-growth conditions, in agreement with experiments. Our work provides insights on the connection between the heterogeneity among genes and the environment-specificity of cell behaviors.
Similar content being viewed by others

Introduction
Cells can adapt to different environments and alter the expression levels of multiple genes. The genome-wide gene expression profile can change dramatically as cells switch between different environments. However, proliferating cells, including bacteria and unicellular eukaryotes, exhibit a growth law as the nutrient quality changes: the fraction of ribosomal proteins in the proteome (ϕR) and the growth rate (μ) are positively correlated. The growth law curve (ϕR vs. μ) is often fit by a linear relation, ϕR = μ/κ + ϕ01,2,3,4,5,6, which can be rationalized by a simple translation model (STM): ribosomes are engaged in translation with a constant translation speed that is proportional to κ2,4. ϕ0 represents the fraction of inactive ribosomes that are not producing proteins, independent of environments in the STM. While the STM is simple and intuitive, it does not always provide a good empirical fitting to the experimental growth law curves, e.g., it apparently breaks down in slow-growth conditions of Escherichia coli (doubling time longer than 60 min at 37 °C) in which more ribosomes are produced than the prediction of the STM7. A similar breakdown was also observed for other bacteria1.
We remark that two important biological features beyond the STM are crucial to interpreting the growth law curve, as we show in this work. The first is the heterogeneous translation speeds of ribosomes producing different proteins. Recent studies showed that the translation speeds are highly heterogeneous among different proteins due to multiple mechanisms, including codon usages8 and amino acid compositions9. In particular, the translation speeds of ribosomal proteins are much slower than the average translation speed over non-ribosomal proteins due to the abundance of positively charged amino acids in ribosomal proteins9. Nowadays, the ribosome profiling technique allows us to quantify the allocation of ribosomes toward the production of different proteins. These experimental techniques enable us to rethink the growth law in the presence of heterogeneity in translation speeds9.
The second feature is finite protein degradation rates. The STM neglects protein degradation and predicts that at zero growth rate, ϕR = ϕ0 so that all ribosomes are inactive. However, this contradicts experiments of nongrowing bacteria in which translation activities are observed10. Protein degradation must be considered at zero growth rate to balance protein production to ensure a constant protein mass. Therefore, protein degradation must be important to the growth law, at least in slow-growth conditions.
In this work, we show that the heterogeneous translation speeds and protein degradations significantly influence the growth law by introducing a general theoretical framework of protein synthesis. We find that the fraction of ribosomal proteins ϕR depends not only on the growth rate but also on the statistical properties of environments. Besides the growth rate, ϕR depends on two correlation coefficients among proteins. One is between the translation speeds and ribosome allocations towards the production of different proteins. The other is between the degradation rates and mass fractions of proteins. Both correlation coefficients are environment-specific. We compute the above correlation coefficients using proteomics and ribosomal profiling datasets of S. cerevisiae11. Interestingly, we find that the correlation between the translation speed and ribosome allocations becomes stronger when the growth rate decreases; cells tend to produce more proteins with higher translation speeds in poor nutrients. In contrast, the correlation between the protein degradation rates and mass fractions is almost independent of growth rates.
We derive the general form of growth law involving the above correlations and demonstrate that for environments with similar correlation coefficients, the growth law curve is universal and has the following form, ϕR = (μ + c1)/(c2μ + c3) where c1, c2, and c3 are constants depending on the above correlation coefficients. In particular, c2, which sets the nonlinearity of the growth law curve, is finite due to the slow translation speed of ribosomal proteins. We prove that a universal growth law curve must be monotonically increasing and convex. Surprisingly, we find that a universal growth law applies to E. coli and our theories justify the upward bending of the growth law curve of E. coli in slow-growth conditions relative to a linear line7. However, if the experiments are implemented in multiple environments with dramatically different correlation coefficients, the growth law curve is non-universal and environment-specific. Our analysis of experimental data suggests that this scenario may apply to S. cerevisiae. We fit the experimentally measured growth law curves by our model predictions, from which we can estimate the translation speed of ribosomal and non-ribosomal proteins. Consistent with direct experimental measurements9, the estimated translation speed of ribosomal proteins is indeed much slower than non-ribosomal proteins.
Results
Model of protein synthesis
Given a constant environment, we consider a population of cells with a constant growth rate, and the protein synthesis processes are in a steady state. Ribosome profiling allows us to quantify the fraction of ribosomes in the pool of total active ribosomes producing protein i, which we call ribosome allocation χi. Here the index i represents one particular protein i. Mass spectrometry also allows us to measure the mass fractions ϕi of all proteins in the proteome12. The translation speed of ribosomes on the corresponding mRNAs is ki, which is the averaged mass of translated amino acids per unit time. Note that ki is averaged over the sequence of the corresponding mRNA so that each protein has one ki. We also assume that protein i degrades with a constant rate αi. The mass production rate of protein i becomes
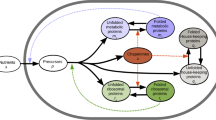
Here R is the number of ribosomes, and R0 is the number of inactive ribosomes. Our model is summarized in Fig. 1.

Given a constant environment, cells allocate different fractions of active ribosomes (χi) to translate mRNAs corresponding to different proteins. In general, the translation speeds ki are heterogeneous among proteins. αi is the degradation rate of protein i. χi, ki and αi together determine the mass fraction of protein i. The ribosome allocation strategies reflect the adaptation of cells to different environments. In this schematic, we show three proteins for simplicity.
In this work, we focus on the effects of heterogeneous translation speeds ki and finite degradation rates αi. Therefore, for simplicity, we assume them to be invariant of environments. We also mainly consider the effects of nutrient quality and do not consider the impact of antibiotics in this work, which can decrease the overall effective translation speed and increase ϕR as the growth rate decreases4.
We define the total protein mass M = ∑iMi, and the protein mass fraction ϕi = Mi/M. Using Eq. (1), we find the fraction of ribosomal proteins in the proteome in the steady state, (see detailed derivations in Methods)
Here μ is the growth rate of the total protein mass \(\mu =\dot{M}/M\), and mR is the total amino acid mass of a single ribosome. ϕ0 is the mass fraction of inactive ribosomes, which we assume to be constant for simplicity. In this work, i = 1 is reserved for ribosomal proteins so that ϕ1 = ϕR, k1 = kR, and α1 = αR. Here, kR and αR are the effective translation speed and degradation rate of the coarse-grained ribosomal protein averaged over all ribosomal proteins. They are approximately independent of environments due to the tight regulation of relative doses of different ribosomal proteins13 and their generally low degradation rates. It is easy to find that if all proteins have the same translation speed (ki = k for all i) and protein degradations are negligible (αi = 0), Eq. (2) is reduced to the STM.
Universal and non-universal growth law curves
To disentangle the effects of heterogeneous translation speeds and protein degradations, we first simplify the model by taking αi = 0 for all proteins and only consider the effects of heterogeneous translation speeds ki. We rewrite \({\sum }_{i}{k}_{i}{\chi }_{i}={k}_{R}{\chi }_{R}+(1-{\chi }_{R})\mathop{\sum }\nolimits_{i = 2}^{N}{k}_{i}{\widetilde{\chi }}_{i}\) in Eq. (2). Here, N is the number of proteins and \({\chi }_{i}=(1-{\chi }_{R}){\widetilde{\chi }}_{i}\) so that \(\mathop{\sum }\nolimits_{i = 2}^{N}{\widetilde{\chi }}_{i}=1\). In the following, we define \({\langle k\rangle }_{\chi }=\mathop{\sum }\nolimits_{i = 2}^{N}{k}_{i}{\widetilde{\chi }}_{i}\) as the χ-weighted average translation speed over all non-ribosomal proteins. As we derive in Methods, the fraction of ribosomal proteins can be written exactly as a Hill function of the growth rate:
where the expressions of a, b are shown in Methods. We are particularly interested in the sign of a because it determines the shape of the ϕR(μ) curve. Interestingly, we find that a ∝ kR − 〈k〉χ. If kR is smaller than 〈k〉χ, a is negative so that the second derivative of the ϕR(μ) curve is positive. In other words, the ϕR(μ) curve is upward bent in slow-growth conditions relative to a linear line.
〈k〉χ depends on both the elongation speeds ki and the ribosome allocations χi. To find its value, we further rewrite 〈k〉χ = 〈k〉(1 + Iχ,k). Here 〈k〉 is the arithmetic average of translation speeds over all non-ribosomal proteins. Iχ,k is a metric we use to quantify the correlation between the ribosome allocations and the translation speeds:
Here, the bracket represents an average over all non-ribosomal proteins. Biologically, the higher Iχ,k is, the more ribosomes are allocated to mRNAs with higher translation speeds. Because the ribosomal allocations χi are generally different in different environments, we use Iχ,k to characterize an environment. Imagine that we grow cells in multiple environments with equal Iχ,k. We find that as long as Iχ,k is not too close to −1, which we confirm later using experimental data, a is always negative since the translation speed of ribosomal proteins kR is much lower than 〈k〉9. Therefore, Eq. (3) predicts an upward bending of the ϕR(μ) curve in slow-growth conditions.
We verify the above theoretical predictions by numerically simulating the model of protein synthesis (Methods). The translation speeds are randomly sampled among proteins and fixed for all environments, with kR < 〈k〉. We randomly sample χi for each environment and compute the resulting growth rate μ and protein mass fractions ϕi. We show the results from environments with preselected Iχ,k, which agree well with the theoretical formula Eq. (3) (Fig. 2a).

a We simulate the case of heterogeneous translation speeds without protein degradations and compare our numerical simulations with model predictions (dashed lines). Each data point has its own randomly sampled χi, and we show the results with preselected Iχ,k values. The red dash line represents the predictions of the STM in which all proteins have the same translation speed 〈k〉. b Same analysis in which we simulate the case of finite protein degradation rates without heterogeneity in translation speeds.
We also consider another simplified model in which the translation speeds are homogeneous, but protein degradation rates are finite and heterogeneous. We find that in this model, the growth law curve is linear with a reduced slope and increased intercept compared with the STM (see details in Methods). The actual shape of the growth law curve depends on the parameter Iϕ,α, which is a metric to characterize an environment by quantifying the correlation between the protein mass fractions and degradation rates:
Here, the bracket represents an average over all non-ribosomal proteins and \({\widetilde{\phi }}_{i}=(1-{\phi }_{R}){\phi }_{i}\). Biologically, a high Iϕ,α value means that the proteome are enriched with proteins with high degradation rates. We verify the above theoretical predictions by numerical simulations and randomly sample the protein degradation rates that are fixed for all environments. We show the results from environments with preselected Iϕ,α and our theoretical predictions Eq. (18) are nicely confirmed (Fig. 2b).
Finally, we turn to the full model with both the heterogeneities in the translation speeds and protein degradation rates. We find that the growth law curve has the following general form,
where the expressions of the constants, c1, c2 and c3 are shown in Methods. We prove that given fixed Iχ,k and Iϕ,α (as long as they are not too close to −1), the growth law curve must be monotonically increasing and convex, which suggests an upward bending in slow-growth conditions (Methods). In particular, c2 ∝ kR − 〈k〉χ, which means that it is the slower translation speed of ribosomal proteins than other proteins that generates the nonlinear shape of the growth law curve. The simulation results match the theoretical predictions (Fig. 3a). We note that the uncertainness of real environments often leads to random production of proteins and random allocation of ribosomes. To address this question, we also simulate models in which noises exist in the translation speeds ki and the allocation fractions χi. We find that both noises do not affect our conclusions qualitatively (Fig. S1). Note that adding noises to the translation speeds and allocation fractions only makes the resulting growth law curves even noisier and therefore does not affect our main conclusion that the growth law curve is generally environment-specific, as we show later.

a Numerical simulations with preselected Iχ,k and Iϕ,α. The red dashed line is the prediction of the STM, and other dashed lines represent our model predictions. b, e Two-dimensional Gaussian distribution of randomly sampled Iχ,k and Iϕ,α. The mean of Iχ,k is 0.5, and the mean of Iϕ,α is 0. The standard deviations σ are indicated in the legends. c, f The resulting growth law curve where each point has randomly sampled Iχ,k and Iϕ,α from (b) and (e). d, g The distribution of the fitting RMSE corresponding to randomly chosen points in (c) and (f).
In real situations, we remark that the actual growth curve shape depends on the particular environments. To verify this, we compute the resulting growth law curve with multiple environments, and the Iχ,k and Iϕ,α of each environment are randomly sampled from Gaussian distributions (Fig. 3b, e) (Methods). We find that when the Gaussian distributions have large standard deviations, the growth law curve is non-universal and depends on the particular chosen environments (Fig. 3c). This means that if we randomly pick some environments from Fig. 3c, the resulting growth law curves are generally different. In contrast, when the Gaussian distributions have small standard deviations, the growth law curve is well captured by our theoretical predictions Eq. (6) because the environments share similar Iχ,k and Iϕ,α (Fig. 3f). To quantify the effects of heterogeneous Iχ,k and Iϕ,α across environments, we repeatedly sample 20 random points from Fig. 3c, f and fit them using Eq. (6) (Methods), imitating the sampling processes in real experiments. We find that when the chosen environments have significantly different Iχ,k and Iϕ,α, the median root mean squared error RMSE = 1.69 × 10−2 (Fig. 3d). In contrast, in the case of similar environments, RMSE = 4.44 × 10−3 (Fig. 3g). The above results suggest that we can use the fitting error as a criterion of the universality of the growth law curve, which we apply to the experimental data later.
Experimental tests of theories
In this section, we test our model using published datasets of S. cerevisiae14 (Methods). For each strain and nutrient quality, we computed the correlation coefficients between the translation speeds and ribosome allocations Iχ,k, and the correlation coefficients between the protein degradation rates and protein mass fractions Iϕ,α. Given the values of μ, Iχ,k, and Iϕ,α, we predicted the fraction of ribosomal proteins ϕR using Eq. (6) (Fig. 4a, d). We note that one parameter ϕ0 is not known experimentally. By choosing a common ϕ0 = 0.048, our model predictions nicely match the experimentally measured values of ϕR (with one data point slightly above the theoretical prediction). We find that regardless of the data processing procedures, the relative relationships between the predicted curves always agree with those of the experimental values (Methods and Fig. S2).

a Experimental measured ϕR of S. cerevisiae along with the predictions (dashed lines) of our model. b The growth rate dependences of the correlation coefficients Iχ,k and Iϕ,α. c The normalized enrichment scores (NES) of GSEA of enriched gene sets. A positive NES of ki-ordered GSEA means that the genes in the corresponding gene set are enriched in the regime of higher ki. A positive NES of \({\log }_{2}\)FC-ordered GSEA means that the genes in the corresponding gene set are enriched in the regime of increasing χi when the nutrient changes from glucose to glycerol. d Summary of the variables and parameters in the analysis of experimental data. The effective mass of ribosomal proteins mR is calculated based on molecular weights of ribosomal proteins detected in the proteome (Methods). SC synthetic complete medium, Glu glucose, Gly glycerol.
Our model is simplified as we assume that the translation speeds and protein degradation rates do not depend on environments. Remarkably, our model predictions still quantitatively match the experimental observations, suggesting that our assumptions may be reasonable for most situations. While our model cannot predict the growth rate dependence of ϕ0, our results show that a constant ϕ0 is consistent with three of four data points in Fig. 4a, and the outlier may have a higher ϕ0 in that particular environment. Our analysis cannot exclude the possibility that ϕ0 is also environment-specific.
Interestingly, we found that Iϕ,α ≈ −0.33 for all the conditions we computed. However, Iχ,k are negatively correlated with the growth rates, suggesting cells tend to allocate more ribosomes to translate mRNAs with higher ki in poor nutrient conditions (Fig. 4b). To find out what genes acquire more resources when the environment is shifted, we perform Gene Set Enrichment Analysis (GSEA)15,16 for wild type cells (Methods) and find that eight gene sets from the Gene Ontology (GO)17,18 database are enriched in both the GSEA where genes are ordered by ki and the GSEA where genes are ordered by \({\log }_{2}\) fold change (\({\log }_{2}\)FC) of χi when the nutrient changes (Fig. S3a).
We find that five gene sets related to stress response are enriched in the regime of higher ki and increasing χi when the environment is changed from 2% glucose to 2% glycerol (Fig. 4c). This is consistent with the environmental stress response (ESR) of S. cerevisiae as an adaptation to the shifts of environments19. We propose that higher translation speeds of stress response proteins enable cells to respond rapidly to environmental changes, which is evolutionarily advantageous. We also find two gene sets related to the rRNA process enriched in the regime of lower ki and decreasing χi (Fig. 4c). We also perform GSEA for natAΔ cells and get similar results (Fig. S3b, c).
Applications of theories to experimental growth law curves
An important application of our theories is that one can estimate the translation speeds by fitting the experimental growth law curve to our model prediction Eq. (6) (Methods). Because there are 6 unknown parameters in the definition of c1, c2, and c3 (Eqs. (23)–(25)), we can estimate three of the parameters given the values of the other three. For the S. cerevisiae data from Ref. 6, we use the experimentally measured degradation rate of ribosomal proteins αR and the mass of ribosomal proteins mR as given. We approximate the ϕ-averaged degradation rate 〈α〉ϕ by 〈α〉(1 + Iϕ,α) where Iϕ,α = −0.33, justified by the observations that Iϕ,α is independent of environments (Fig. 4b). We find that the fitted parameters c1, c2 and c3 having a wide range of 95% confidence intervals (Fig. 5a, c) with RMSE = 1.35 × 10−2. The inferred ranges of ϕ0, kR and 〈k〉χ are also unreasonably large (Fig. 5c). All these results suggest that the growth law curves of S. cerevisiae are non-universal and large variations of Iχ,k and Iϕ,α exist among environments (Fig. 3d).

a The non-linear fitting to data of S. cerevisiae from Ref. 6. The shadow represents the 95% prediction interval. b The non-linear fitting to data of E. coli from Ref. 7. c Detailed fitting results of (a) and (b). Note that the reference value of 〈k〉χ of (a) is approximated by 〈k〉(1 + Iχ,k) where the minimal Iχ,k = 0.608 and the maximal Iχ,k = 2.415 (Fig. 4d).
We also apply our theories to E. coli7 (Fig. 5b). Because most proteins are non-degradable in bacteria20,21, we set αR and 〈α〉ϕ as 0, and the mass of ribosomal protein mR = 8.07 × 105Da12. In this case, the fitted parameters have much smaller range of 95% confidence intervals with RMSE = 3.60 × 10−3. The estimated kR, and 〈k〉χ are consistent with previous studies22,23,24 (Fig. 5c). Our analysis of experimental data demonstrates that the translation speed of ribosomal proteins is indeed smaller than the χ − averaged translation speed, in agreement with experimental observations9. Our results suggest that E. coli has similar values of Iχ,k and Iϕ,α in the chosen environments of Ref. 7 so that it has a universal growth law curve. In contrast, S. cerevisiae appears to have significantly different Iχ,k and Iϕ,α across different environments of Ref. 6 so that the growth law curve depends on the chosen environments and therefore non-universal.
Discussion
It has been known since the 1950s that the chemical compositions and cell size of bacteria are functions of growth rate and seem to be independent of the medium used to achieve the growth rate25. This view has been broadly accepted in the study of bacteria physiologies in the past decades. The growth law acquired its name because of the independence of the environment. However, recent findings hint at an unforeseen complexity in the growth law. For example, bacterial cell sizes have been shown to depend on the presence of antibiotics, and overexpression of useless proteins26, and dramatically different cell sizes can exist at the same growth rate27. Our study focuses on the growth law regarding the fraction of ribosomal proteins in the proteome and further uncovers the importance of environment-specificity to microbial physiologies. We go beyond the simple translation model and take account of the heterogeneous translation speeds and finite protein degradations. Given the translation speeds and protein degradation rates, our model is completely general and virtually applies to any cells, including both proliferating cells (μ > 0) and non-proliferating cells (μ = 0). In this work, we mainly consider the scenario in which the growth rate changes due to the nutrient quality and the fraction of ribosomal proteins (ϕR) increases monotonically as the growth rate increases.
We demonstrate that the growth law curve generally has the form of Eq. (6). The actual shape of the growth law curve depends on two correlation coefficients: one is between the ribosome allocations and the translation speeds (Iχ,k); the other is between the protein mass fractions and protein degradation rates (Iϕ,α). By analyzing the dataset from14, we found that Iϕ,α is independent of growth rate, while Iχ,k appears to be negatively correlated with the growth rate. This means that cells tend to produce proteins with faster translation speeds in slow-growth conditions, which can be an economic strategy under evolutionary selection. Remarkably, our theoretical predictions of ϕR reasonably match the experimentally measured values14. We note that the upward bending of the growth law curves of bacteria compared with a linear relation appears to hint at an increasing fraction of inactive ribosomes in slow-growth conditions. While this mechanism appears plausible, it has not been confirmed experimentally as we realize. In our work, we demonstrate that the apparent upward bending can be merely a consequence of heterogeneous translation speeds among proteins and therefore raises caution on the biological interpretation of the shape of the growth law curve.
We apply our model predictions to the growth law curves of S. cerevisiae6 and E. coli7. In the former case, data fitting to our model prediction is subject to very large uncertainty. This observation agrees with the computed Iχ,k that are variable across conditions using the ribosome profiling and mass spectrometry data from14 (Fig. 4b). In contrast, the fitting of E. coli data exhibits a much smaller uncertainty, suggesting that common Iχ,k and Iϕ,α may apply to all the nutrient qualities used in the experiments of Ref. 7. We expect that this idea can be tested when genome-wide measurements, such as the translation speeds of E. coli are available in the future so that critical parameters such as Iχ,k can be calculated for E. coli.
We remark that in the absence of heterogeneous translation speeds and protein degradation, the mass fraction of protein i, ϕi must equal the ribosome allocation χi. Indeed, these two datasets are often highly correlated among proteins in E. coli12,28. However, in a more realistic scenario, ϕi also depends on the translation speed and protein degradation rate. Given the same χi, proteins with higher translation speeds or lower degradation rates should have higher mass fractions (Methods). We note that using the current genome-wide datasets of S. cerevisiae, the predicted protein mass fractions ϕi,pre based on the ribosome allocations χi14, the translation speeds ki9, and the protein degradation rates αi11 do not correlate strong enough with the measured ϕi. We note that these datasets are from different references, and the deviation is likely due to the noise in the measurements of ki (Table S2). We expect our theories to be further verified when more accurate measurements of translation speeds are available.
For simplicity, in this work, we assume that the translation speeds and protein degradation rates are invariant as the nutrient quality changes. Therefore, we can use the two correlation coefficients Iχ,k and Iϕ,α to characterize a particular environment. We remark that our model can be generalized to more complex scenarios in which the translation speeds or protein degradation rates depend on the growth rate7. In this case, one just needs to include four additional environment-specific parameters: kR, 〈k〉, αR, and 〈α〉.
Methods
Derivations of Equation (2)
All variables are summarized in Table S3.
Because the total protein mass M = ∑iMi, we sum over all proteins on both sides of Eq. (1) and obtain
We then divide both sides by M and obtain the expression of the growth rate
from which Eq. (2) is obtained.
We can also find the changing rate of ϕi = Mi/M using Eq. (1),
which leads to the expression of ϕi in the steady state as
Since all proteins grow in the same rate in the steady-state, the growth rates of protein i defined as
must be equal to μ, which can be easily verified using Eq. (10). Using the normalization condition ∑iϕi = 1, we can write ϕi using Eq. (10) as
We can also write the normalization condition as
Details of the simplified model without protein degradation
In deriving Eq. (3), we neglect protein degradation and rewrite Eq. (2) as
Meanwhile, we compute the growth rate using the auto-catalytic nature of ribosomal proteins,
The above equation allows us to replace χR by μ in Eq. (14), from which we obtain Eq. (3) where
Details of the simplified model with finite protein degradation rates
We now discuss the effects of finite protein degradation rates and assume that the translation speeds are homogeneous and equal to k for all proteins. We rewrite the ∑iαiϕi term in Eq. (2) such that \({\sum }_{i}{\alpha }_{i}{\phi }_{i}={\alpha }_{R}{\phi }_{R}+(1-{\phi }_{R})\mathop{\sum }\nolimits_{i = 2}^{N}{\alpha }_{i}{\widetilde{\phi }}_{i}\). Here, \({\phi }_{i}=(1-{\phi }_{R}){\widetilde{\phi }}_{i}\) so that \(\mathop{\sum }\nolimits_{i = 2}^{N}{\widetilde{\phi }}_{i}=1\). We define the ϕ − averaged degradation rates over all non-ribosomal proteins as \({\langle \alpha \rangle }_{\phi }=\mathop{\sum }\nolimits_{i = 2}^{N}{\alpha }_{i}{\widetilde{\phi }}_{i}\). Therefore, Eq. (2) can be written as
where
To find the sign of d, we further rewrite 〈α〉ϕ as 〈α〉ϕ = 〈α〉(1 + Iϕ,α) where 〈α〉 is the arithmetic average of degradation rates over all non-ribosomal proteins.
Imagine that we grow cells in multiple environments with equal Iϕ,α. We assume that the degradation rate of ribosomal protein αR is slower than the average of non-ribosomal proteins 〈α〉, which is biologically reasonable since ribosomal proteins are generally non-degraded. Therefore, as long as Iϕ,α is not too close to −1, which we confirm using experimental data, d is positive since αR is always smaller than 〈α〉ϕ. Therefore, our model predicts that the growth law curve is linear given a constant Iϕ,α and finite protein degradation decreases the slope relative to the STM. The intercept at μ = 0 is also larger than ϕ0. Therefore, a finite fraction of ribosomes are still translating at zero growth rate. We verify the above theoretical predictions by numerical simulations and randomly sample the protein degradation rates that are fixed for all environments, with αR < 〈α〉 satisfied.
Derivations of the full model
In this section we derive the full model considering both the heterogeneities in the translation speeds and protein degradation rates. We rewrite Eq. (2) in the main text as
Meanwhile, the growth rate is
Combining Eq. (21) and Eq. (22) allows us to solve ϕR as a function of μ and we obtain Eq. (6)
where
It is straightforward to find that the condition for Eq. (6) to be monotonically increasing is that c3 > c1c2. Using the above expressions, we find that
We find that the first two terms are always positive, and the last term is positive as long as Iα,ϕ is not too close to −1. Therefore, the ϕR(μ) curve must increase monotonically. It is straightforward to find that the second derivative of the ϕR(μ) curve is proportional to (c1c2 − c3)c2, which is always positive as long as Iχ,k is not too close to −1.
Details of the numerical simulations
We summarize the parameters we use in the numerical simulations in Table S1. We consider a cell with 4000 genes. We set the elongation speed ki and the degradation rates αi of non-ribosomal genes to follow lognormal distributions. We set kR = 2.07 × 104 Da/min, 〈k〉 = 4.80 × 104 Da/min, αR = 4.83 × 10−4min−1, and 〈α〉 = 1.10 × 10−3min−1 as the experimentally measured values of S. cerevisiae9,11. The coefficients of variation (CV) of the lognormal distributions can be found in Table S1. In all simulations, we set ϕ0 = 0.08. We note that in Fig. 2a, we set αi = 0 for all proteins and in Fig. 2b, we set ki = 〈k〉 for all proteins.
To simulate a random environment, we generate a random χR. Meanwhile, a lognormal distribution of χi of non-ribosomal genes is also randomly generated. The CV of the lognormal distribution is included in Table S1. We then search for the ϕR and μ that simultaneously satisfy Eq. (22) and Eq. (13). ϕi, Iχ,k and Iϕ,α are then calculated using Eq. (10), Eq. (4) and Eq. (5), respectively. For a chosen pair of Iχ,k and Iϕ,α, the predicted ϕR(μ) curve is obtained using Eq. (6). To obtain Fig. 3d, g, we randomly sample 20 points from Fig. 3c, f respectively, fit them using Eq. (6), and calculate the resulting RMSE. We repeat the above process 5000 times.
Details of the experimental data analysis
For the ribosome profiling data14, we first trim the adapter with Cutadapt (version 3.4)29. Then we use Bowtie2 (version 2.4.2)30 to eliminate ribosomal RNAs (rRNA) as mentioned in31. The cleaned reads are then mapped to S. cerevisiae genome R64.1.1 with HISAT2 (version 2.2.1)32. Read counts are then generated with featureCount (version 2.0.1)33. We then manually eliminate the non-coding RNAs. The ribosome allocation χi is calculated based on the mean count fraction of all samples (Supplementary Data 3).
For the proteomics data14, we perform the absolute quantification (or the in-sample relative quantification) of proteins based on the intensities of peptides using xTop (version 1.2)12. The intensity ratio of 2 proteins in the same sample of proteomics data does not directly represent the real abundance (either the mass or the copy number) ratio, so the abundance fraction can not be replaced with the intensity fraction12,34. xTop is a software that accurately calculates the in-sample relative protein copy number with the maximum a posteriori probability (MAP) algorithm12. We then calculate all proteins’ mass fraction ϕi with the xTop results and the protein molecular mass (Supplementary Data 2). In12, the authors further calibrated ϕi with ribosome profiling data assuming homogeneous ki. In this work, we alternatively calibrate ϕi with L−0.57 where L is the protein length, as mentioned in12. Calibration with L−0.57 is independent of ribosome profiling data, although it reduces the distance between χi and calibrated ϕi12. We also show the result with calibration of L−1 or without calibration in Fig. S2b, c. To compute ϕR, we sum up the ϕi of all proteins annotated as the cytoplasmic ribosomal protein in the Saccharomyces Genome Database (SGD).
For the elongation speed ki, we first calculate vi as mentioned in9. ki is then calculated using the relationship ki = viai where \(v_i\) is the number of translated amino acids per unit time, and \(a_i\) is the averaged mass of amino acids over the sequence of protein \(i\) (Supplementary Data 1). For the degradation rate αi, data is obtained from11. We calculate the experimental Iχ,k, Iϕ,α, 〈k〉 and 〈α〉 for non-ribosomal genes that exist in all data sets of χi, ϕi, ki and αi. We also calculate the χ-averaged k of ribosomal proteins as kR and ϕ-averaged α of ribosomal proteins as αR.
For the molecular mass of the ribosome, we calculate the effective mR. Considering the efficiency of the mass spectrometry (MS), not all proteins can be detected. Therefore, we define the effective mR as the molecular weights of ribosomal proteins detected in the proteome. Because most of the ribosomal proteins can be expressed by two paralogous genes in S. cerevisiae, we count the average molecular mass when both proteins of the paralogs are detected in the proteome. We also show our predictions of ϕR using the actual ribosome mass (mR = 1.40 × 106 Da) in Fig. S2a.
For the growth rate μ (Supplementary Data 4), it is obtained from the growth curve, OD600 versus time from14 with the method mentioned in35. Briefly, the slopes of \(\ln ({{{\mbox{OD}}}}_{600})\) versus time in 5-point windows are calculated. Then windows with slopes at least 95% of the maximum slope are extracted. The slope of points within these windows is calculated as the growth rate. With these results, we predict the corresponding ϕR(μ) curves and compare them with the experimental data points.
We further calculate the predicted mass fraction ϕi,pre of non-ribosomal proteins with Eq. (12). Pearson correlation coefficients ρ between ϕi,pre and ϕi are calculated. We also compute ρ under the assumptions that αi = 0 or ki = 〈k〉 (Table S2).
For GSEA analysis, we first perform the differential expression analysis on the ribosome profiling data of WT or natAΔ cells using the package DEseq2 (version 1.24.0)36 in R (version 3.6.1). The \({\log }_{2}\) fold changes in counts when cells changed from SC+2% glucose to SC+2% glycerol, the p-value of the two-sided Wald test, and the FDR q-values are calculated. Ribosomal genes and genes with FDR q-value > 0.05 are eliminated. We then pick out genes that also exist in the data sets of ki. GSEA on these genes is performed twice using the R package clusterProfiler (version 3.12.0)37 and org.Sc.sgd.db (version 3.8.2)38. In the first GSEA, genes are ordered by the \({\log }_{2}\) fold change (denoted as \({\log }_{2}\)FC-ordered GSEA). In the second GSEA, genes are ordered by ki (denoted as ki-ordered GSEA). We then find the common gene sets from GO database17,18 enriched in these two GSEA. The cut-off criteria are set as the p-value < 0.05 (single-sided permutation test) and the FDR q-value < 0.25. The number of permutations used in the analysis is 105.
Details of fitting in Fig. 5
Nonlinear fitting is performed with MATLAB (version R2020b). We obtain the fitting parameters c1, c2 and c3 with their 95% confidence intervals, and then compute ϕ0, kR and 〈k〉χ using Eqs. (23), (24), (25). To compute the ranges of these values, we numerically find the maximum and the minimum value of the multivariate functions ϕ0(c1, c2, c3), kR(c1, c2, c3) and 〈k〉χ(c1, c2, c3) as their upper and lower bounds, where the ranges of c1, c2 and c3 are their 95% confidence intervals.
Statistics and reproducibility
We use the two-sided Wald test in the differential expression analysis of the ribosome profiling data. In GSEA, we use a single-sided permutation test. As for reproducibility, no biological experiments are performed in our work, and all data are acquired from public repositories (see Data Availability).
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The Ribosome profiling data from14 was deposited at GEO (GSE140255) (link https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE140255). The Proteomics data from14 was deposited at ProteomeXchange (PXD015217) (link http://proteomecentral.proteomexchange.org/cgi/GetDataset?ID=PXD015217). The growth rate data was acquired from the figures of OD600-versus-time curves in14. The Ribosomal protein list was acquired from Saccharomyces Genome Database (SGD) (link https://yeastgenome.org). The data needed calculating elongation speeds was acquired from the supplementary materials of9. The protein degradation rate data was acquired from the supplementary materials of11. The ϕR-μ data of E. coli was acquired from the supplementary materials of7. The ϕR-μ data of S. cerevisiae was acquired from the figure in6. Calculated data of \(k_i\), \(\phi_i\), \(\chi_i\), and \(\mu\) have been provided in Supplementary Data 1-4.
Code availability
All codes for mathematical simulations and data analysis are available in the following link (https://github.com/QirunWang/Codes-for-Environment-specificity-and-universality-of-the-growth-law).
References
Neidhardt, F. C. & Magasanik, B. Studies on the role of ribonucleic acid in the growth of bacteria. Bioch. Biophys. Acta 42, 99–116 (1960).
Maaløe, O. Regulation of the protein-synthesizing machinery—ribosomes, trna, factors, and so on. In Biological regulation and development, 487–542 (Springer, 1979).
Bremer, H. & Dennis, P. P. Modulation of chemical composition and other parameters of the cell at different exponential growth rates. EcoSal Plus 3 (2008).
Scott, M., Gunderson, C. W., Mateescu, E. M., Zhang, Z. & Hwa, T. Interdependence of cell growth and gene expression: origins and consequences. Science 330, 1099 (2010).
Hui, S. et al. Quantitative proteomic analysis reveals a simple strategy of global resource allocation in bacteria. Mol. Syst. Biol. 11, 784 (2015).
Metzl-Raz, E. et al. Principles of cellular resource allocation revealed by condition-dependent proteome profiling. Elife 6, e28034 (2017).
Dai, X. et al. Reduction of translating ribosomes enables escherichia coli to maintain elongation rates during slow growth. Nat. Microbiol. 2, 1–9 (2016).
Klumpp, S., Dong, J. & Hwa, T. On ribosome load, codon bias and protein abundance. PLoS ONE 7, e48542 (2012).
Riba, A. et al. Protein synthesis rates and ribosome occupancies reveal determinants of translation elongation rates. Proc. Natl Acad. Sci. 116, 15023–15032 (2019).
Gefen, O., Fridman, O., Ronin, I. & Balaban, N. Q. Direct observation of single stationary-phase bacteria reveals a surprisingly long period of constant protein production activity. Proc. Natl Acad. Sci. 111, 556–561 (2014).
Lahtvee, P.-J. et al. Absolute quantification of protein and mrna abundances demonstrate variability in gene-specific translation efficiency in yeast. Cell Syst. 4, 495–504.e5 (2017).
Mori, M. et al. From coarse to fine: the absolute escherichia coli proteome under diverse growth conditions. Mol. Syst. Biol. 17, e9536 (2021).
Li, G.-W., Burkhardt, D., Gross, C. & Weissman, J. S. Quantifying absolute protein synthesis rates reveals principles underlying allocation of cellular resources. Cell 157, 624–635 (2014).
Friedrich, U. A. et al. Nα-terminal acetylation of proteins by nata and natb serves distinct physiological roles in saccharomyces cerevisiae. Cell Rep. 34, 108711 (2021).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. 102, 15545–15550 (2005).
Mootha, V. K. et al. Pgc-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 34, 267–273 (2003).
Ashburner, M. et al. Gene ontology: tool for the unification of biology. Nat. Genet. 25, 25–29 (2000).
Consortium, T. G. O. The Gene Ontology resource: enriching a Gold mine. Nucleic Acids Res. 49, D325–D334 (2020).
Gutin, J., Sadeh, A., Rahat, A., Aharoni, A. & Friedman, N. Condition-specific genetic interaction maps reveal crosstalk between the camp/pka and the hog mapk pathways in the activation of the general stress response. Mol. Syst. Biol. 11, 829 (2015).
Erickson, D. W. et al. A global resource allocation strategy governs growth transition kinetics of escherichia coli. Nature. 551, 119–123 (2017).
Goldberg, A. L. & John, A. C. S. Intracellular protein degradation in mammalian and bacterial cells: Part 2. Annu. Rev. Biochem. 45, 747–804 (1976).
Arkin, A., Ross, J. & McAdams, H. H. Stochastic kinetic analysis of developmental pathway bifurcation in phage lambda-infected escherichia coli cells. Genetics. 149, 1633–1648 (1998).
Gouy, M. & Grantham, R. Polypeptide elongation and trna cycling in escherichia coli: a dynamic approach. FEBS Lett. 115, 151–155 (1980).
Karpinets, T. V., Greenwood, D. J., Sams, C. E. & Ammons, J. T. RNA: protein ratio of the unicellular organism as a characteristic of phosphorous and nitrogen stoichiometry and of the cellular requirement of ribosomes for protein synthesis. BMC Biol. 4, 1–10 (2006).
Schaechter, M., MaalØe, O. & Kjeldgaard, N. O. Dependency on medium and temperature of cell size and chemical composition during balanced growth of salmonella typhimurium. Microbiology 19, 592–606 (1958).
Basan, M. et al. Inflating bacterial cells by increased protein synthesis. Mol. Syst. Biol. 11, 836 (2015).
Vadia, S. & Levin, P. A. Growth rate and cell size: a re-examination of the growth law. Curr. Opin. Microbiol. 24, 96–103 (2015).
Liu, T.-Y. et al. Time-resolved proteomics extends ribosome profiling-based measurements of protein synthesis dynamics. Cell Systems 4, 636–644.e9 (2017).
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. Journal 17, 10–12 (2011).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359 (2012).
Galmozzi, C. V., Merker, D., Friedrich, U. A., Döring, K. & Kramer, G. Selective ribosome profiling to study interactions of translating ribosomes in yeast. Nat. Protocols 14, 2279–2317 (2019).
Kim, D., Paggi, J. M., Park, C., Bennett, C. & Salzberg, S. L. Graph-based genome alignment and genotyping with hisat2 and hisat-genotype. Nat. Biotechnol. 37, 907–915 (2019).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2013).
Calderón-Celis, F., Encinar, J. R. & Sanz-Medel, A. Standardization approaches in absolute quantitative proteomics with mass spectrometry. Mass Spectrometry Rev. 37, 715–737 (2018).
Hall, B. G., Acar, H., Nandipati, A. & Barlow, M. Growth rates made easy. Mol. Biol. Evol. 31, 232–238 (2013).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 15, 550 (2014).
Yu, G., Wang, L.-G., Han, Y. & He, Q.-Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS: J. Integrative Biol. 16, 284–287 (2012).
Carlson, M. org.Sc.sgd.db: genome wide annotation for yeast. https://bioconductor.org/packages/release/data/annotation/html/org.Sc.sgd.db.html (2019).
Acknowledgements
We thank Poyi Ho for useful discussions related to this work. The research was funded by National Key R&D Program of China (2021YFF1200500) and supported by grants from Peking-Tsinghua Center for Life Sciences.
Author information
Authors and Affiliations
Contributions
Q.W. and J.L. conceived, designed, and carried out the theoretical and numerical part of this work. Q.W. performed the analysis of experimental data. All the authors contributed to the preparation of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Tianshou Zhou and Trinh Hoang for their contribution to the peer review of this work. Primary Handling Editors: Quan-Xing Liu and Gene Chong. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, Q., Lin, J. Environment-specificity and universality of the microbial growth law. Commun Biol 5, 891 (2022). https://doi.org/10.1038/s42003-022-03815-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-022-03815-w
This article is cited by
-
Phenotypic heterogeneity follows a growth-viability tradeoff in response to amino acid identity
Nature Communications (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
