
Abstract
Accumulating lines of clinical evidence support the emerging hypothesis that loss-of-function mutations of GATA2 cause inherited hematopoietic diseases, including Emberger syndrome; dendritic cell, monocyte B and NK lymphoid (DCML) deficiency; and MonoMAC syndrome. Here, we show that mice heterozygous for an arginine-to-tryptophan substitution mutation in GATA2 (G2R398W/+), which was found in a patient with DCML deficiency, substantially phenocopy human DCML deficiency. Mice heterozygous for the GATA2-null mutation (G2-/+) do not show such phenotypes. The G2R398W protein possesses a decreased DNA-binding affinity but obstructs the function of coexpressed wild-type GATA2 through specific cis-regulatory regions, which contain two GATA motifs in direct-repeat arrangements. In contrast, G2R398W is innocuous in mice containing single GATA motifs. We conclude that the dominant-negative effect of mutant GATA2 on wild-type GATA2 through specific enhancer/silencer of GATA2 target genes perturbs the GATA2 transcriptional network, leading to the development of the DCML-like phenotype. The present mouse model provides an avenue for the understanding of molecular mechanisms underlying the pathogenesis of GATA2-related hematopoietic diseases.
Similar content being viewed by others


Introduction
GATA2 is a member of the GATA family of transcription factors that recognize the GATA consensus motif ((T/A)GATA(A/G)) and transcriptionally regulate the expression of target genes1,2,3. In mammals, 6 GATA factors have been identified, each of which has two highly conserved C2C2-type zinc finger domains. Of the zinc-finger domains, the C-terminal zinc finger (CF) domain is indispensable for binding to the GATA motif and is thus required for the transcriptional activity of GATA factors4. Among the six mammalian GATA factors, GATA1, GATA2, and GATA3 are hematopoietic GATA factors that were originally identified in hematopoietic cells. However, these factors are also expressed in organs/tissues other than those of the hematopoietic system. For instance, GATA2 has been found to contribute to the development of the urinary tract, endothelial cells and the nervous system5,6,7,8,9,10. Two Gata2-knockout alleles were generated: one was generated by replacing the 5th exon that encodes the CF domain with a neomycin-resistance cassette11, and the other was generated by inserting Gfp cDNA into the translation initiation site of the 2nd exon12. In both cases, homozygotes for the targeted allele died around embryonic day 10 (E10), the stage before the start of fetal liver hematopoiesis11,12. In addition, a floxed allele of GATA2 for conditional gene knockout has been generated and used for functional analyses of GATA2 during pituitary development13.
GATA2 plays important roles in the maintenance of hematopoietic stem cells (HSCs). In fact, severe HSC failure and depletion are induced by conditional deletion of the GATA2 CF domain in fetal and adult mice14,15. The expression of the Gata2 gene changes dynamically during lineage differentiation hierarchy; therefore, GATA2 contributes to hematopoietic homeostasis, especially in the differentiation of granulocytes, mast cells, and dendritic cells16,17,18,19. An intriguing observation in this regard is that in the erythroid lineage, a complex regulatory network induces GATA2 expression in early progenitors to switch to GATA1 expression during differentiation, a process referred to as GATA factor switching20,21,22.
In a clinical study, a gain-of-function mutation caused by a single amino acid substitution in the CF domain of GATA2 was identified in myeloid transformation of chronic myelogenous leukemia23. This mutant GATA2 possesses increased transactivation activity along with enhanced inhibitory effects on the transcription factor PU.1. Soon after this discovery, many loss-of-function GATA2 mutations were identified that cause autosomal inherited and sporadic hematopoietic diseases24,25,26,27. Of note, heterozygotes harboring the latter GATA2 gene mutations were found to exhibit features of various types of hematovascular-associated immunodeficiencies, such as Emberger syndrome, which displays primary lymphedema25; dendritic cell, monocyte B, and NK lymphoid (DCML) deficiency, which is characterized by decreases in circulating B cells (BCs), natural killer cells (NKCs), dendritic cells (DCs), and monocytes26; and MonoMAC syndrome, which is defined as DCML deficiency accompanying Mycobacterium avium complex infection27. Patients with these diseases have a high risk of developing myelodysplastic syndrome (MDS), acute myeloid leukemia (AML) and chronic myelomonocytic leukemia (CMML)18,28. Since the first discovery of inherited GATA2 mutation-related diseases, accumulating evidence has supported the notion that a variety of GATA2 mutations, not only missense and short indel mutations but also large deletions, splicing errors, gene-regulatory perturbations, frame shifts, and nonsense mutations, are involved in the pathogenesis of GATA2-related diseases29.
GATA2 gene mutations can be divided into two types: one causes a reduction in GATA2 expression (quantitative deficit), while the other leads to the production of structural GATA2 mutants (qualitative defect)29. In the latter type, mutations are found to accumulate in the CF domain and are predicted to cause a loss of DNA-binding activity and transactivation activity29,30,31. While characteristic differences in clinical outcomes have not yet been clarified between these two types of GATA2 mutations, higher disease penetrance and a higher incidence of leukemia transformation have been suggested in patients with qualitative defects than in patients with quantitative deficits32,33.
We aimed to investigate these GATA2-related diseases by establishing mouse models. In this regard, we maintained heterozygous mice carrying both types of Gata2-knockout alleles; they were apparently healthy and enjoyed a natural life. Neither type of mouse showed cytopenia, immunodeficiency, or lymphoedema. No published report has examined leukemogenesis in GATA2-heterozygous null mutant mice, but closer examinations revealed that the HSC function of these mice was slightly disrupted34,35. To further explore whether the quantitative deficit of GATA2 can serve as a model of human GATA2-related diseases, we established an alternative mouse model, a hypomorphic G2fGN/fGN mutant mouse line, in which the expression level of GATA2 was reduced to as low as 20% of the endogenous GATA2 level9,36. While approximately 70% of G2fGN/fGN mice died soon after birth due to urogenital deformity, mice that survived adolescence were prone to develop myeloproliferative neoplasms (MPNs) in the adult stage, with features resembling human CMML36. Thus, although quantitative GATA2 deficits convert immature myeloid progenitors to cells that easily expand and generate many descendants, these deficits do not induce cytopenia in BCs, NKCs, DCs or monocytes, which are hallmarks of GATA2 mutation-related diseases, indicating that quantitative deficits of GATA2 are not a good model of GATA2 mutation-related diseases.
Therefore, in this study, we decided to establish a mouse line harboring a qualitative defect of GATA2, hoping to recapitulate the clinical GATA2 mutant-related phenotypes. To this end, we generated mice that expressed the mutant GATA2 protein in which arginine 398 was substituted for tryptophan (G2R398W). We found that heterozygous expression of G2R398W indeed leads to age-progressive multilineage cytopenia of BCs, NKCs, DCs and monocytes, which phenocopies human DCML deficiency, demonstrating that the G2R398W heterozygote serves as a valuable model of human GATA2 mutation-related diseases. Importantly, we also clarified that G2R398W and another mutant GATA2 protein, G2T354M, have a dominant-negative effect on the native GATA2 protein in terms of DNA-binding activity for the specific cis-element configuration. We propose that this dominant-negative effect induces perturbations of the regulation of specific GATA2 target genes, which explains, at least in part, the development of the DCML-deficient-like phenotype.
Results
Generation and characterization of G2 R398W mutant mice
We established GATA2 mutant mouse lines in which arginine 398 of GATA2 was substituted for tryptophan (R398W) by the CRISPR/Cas9 technique (Fig. 1a). This type of mutation corresponds to the human G2R398W mutation that is frequently found in inherited GATA2-related diseases, such as DCML deficiency, MonoMAC syndrome and Emberger syndrome29. We established two founder mice carrying a single-nucleotide c.1192C>T substitution mutation at the Gata2 locus (Fig. 1b) and crossed these founder mice with wild-type mice to generate heterozygous mutant mice (G2R398W/+). The mice were genotyped by using PCR with genotype-specific PCR primer sets (Fig. 1c and Supplementary Fig. 1a). As the two founder mice did not show any apparent difference, we utilized mainly Line-1 offspring for further analyses. Heterozygous G2R398W/+ pups generated by crossing G2R398W/+ and G2+/+ mice were born according to the Mendelian ratio (73 out of 156 newborns) and did not display any growth differences from their wild-type littermates.

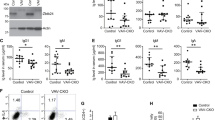
a Schematic illustration of genome editing patterns and locations. b Genomic DNA sequences of two independent founder mice at the targeted region of the Gata2 gene. The signal peaks of the original cytosine (blue) and substituting thymine (red) overlap at position c.1192. c PCR-based genotyping strategy of the G2R398W allele containing the substituted nucleotide. c.1192C>T in the 6th exon is indicated as a red line. Positions recognized by the forward and reverse primers are indicated as arrowheads. d, e Dot plots and mean values for Gata2 transcripts expressed in LSK cells (d) and in subpopulations of LSK cells (e). A primer set detecting transcripts originating from both intact and mutated alleles was used. f Dot plots and mean values for mutated Gata2 transcripts expressed in LSK cells, which were detected using a mutated allele-specific primer set. Number of mice are indicated in parentheses. **p < 0.01. g Immunoblot analysis using nuclear extracts from bone marrow cells. Four independent mice of each genotype were used. Cell extracts from original and murine GATA2-expressing HEK293T cells were used as negative (NC) and positive (PC) controls, respectively. The levels of GATA2 were evaluated by comparing the signal intensity with that of Lamin B. The average value of GATA2 in G2+/+ mice was set to 1.0. The average and individual relative values of GATA2 in individuals are shown in the right panel. *p < 0.05.
We performed quantitative RT-PCR (qRT-PCR) analysis to determine the total transcript expression level of Gata2 (wild-type plus mutant) in bone marrow Lineage–Sca1+cKit+ (LSK) cells from G2R398W/+ mice by utilizing primers covering exons 5 and 6. To our surprise, the total transcript expression level of Gata2 in the G2R398W/+ mice was significantly increased approximately 2.0-fold on average compared with that in the G2+/+ mice, while the expression level of Gata2 in heterozygous Gata2 knockout mice (G2Gfp/+)12 carrying the heterozygous Gfp knockin-knockout allele in the Gata2 locus (G2Gfp allele) was slightly but significantly decreased by ~0.8-fold on average compared with that in the G2+/+ mice (Fig. 1d). We subdivided LSK cells into long-term (LT)-HSCs, short-term (ST)-HSCs, multipotent progenitor 2s (MPP2s), MPP3s and MPP4s and found that the increase in Gata2 expression was striking in LT-HSCs of G2R398W/+ mice, in which the Gata2 expression was high (Fig. 1e). We confirmed that the expression of transcripts derived from the mutated allele was specifically identified in G2R398W/+ mice (Fig. 1f), which indicated that mRNA transcription of Gata2 from the mutant Gata2 locus is active.
We then examined the total protein expression levels of GATA2 in the bone marrow of mice by immunoblotting analysis. Consistent with the qPCR analyses, the GATA2 protein level in the G2R398W/+ mice was elevated approximately 1.2-fold compared to that in G2+/+ mice, while changes in the level of GATA2 protein in G2Gfp/+ mice were not remarkable (Fig. 1g and Supplementary Fig. 1b). These results demonstrate that GATA2 expression is significantly increased in the bone marrow of G2R398W/+ mutant mice. However, the mechanisms underlying this upregulation of Gata2 gene expression remain elusive.
Activity of the G2R398W mutation during mouse embryogenesis
To explore the extent to which the G2R398W mutant protein lost its activity and whether the mutant protein could support mouse embryogenesis, we intercrossed G2R398W/+ mice. We found that homozygous G2R398W/R398W embryos were born following the Mendelian ratio and survived beyond E12.5. However, the homozygous mice died around E14.5, and no homozygous newborns were obtained (Fig. 2a, b). As the critical developmental stage in Gata2-knockout mice is between E10.5 and E11.511, these results demonstrate that the G2R398W mutant protein retains GATA2 activity to a certain extent and is able to support mouse embryogenesis until E14.5.

Intercrossing of G2R398W/+ mice (a, b) and mating of fetuses with G2Gfp/+ mice (c, d). The numbers of embryos of each genotype at the indicated stage are shown (a, c), along with representative photos (b, d). Livers are shown in the insets (b, d). The number of dead embryos is indicated by parentheses. Note that no live G2R398W/Gfp embryo was found at E12.5-E13.5.
To further examine G2R398W mutant protein activity in the context of mouse development, we next crossed the G2R398W/+ mice with the G2Gfp/+ mice and found that compound heterozygous embryos carrying the G2R398W and G2Gfp alleles (G2R398W/Gfp) died by E12.5 (Fig. 2c, d), while heterozygotes of the G2R398W and G2Gfp alleles survived these stages and were born normally. These results indicate that the expression of the G2R398W mutant protein contributes to embryonic development. The difference in the survival of G2R398W/R398W embryos and G2R398W/Gfp embryos further supports the hypothesis that the contribution of this protein occurs in an allele number-dependent manner.
Appearance of DCML-deficient phenotypes in aged G2 R398W/+ mice
We aimed to examine whether heterozygous G2R398W/+ mice showed phenotypes that recapitulate those of human diseases heterozygous for the GATA2 mutant protein, which include Emberger syndrome, DCML deficiency, and MonoMAC syndrome29. Therefore, to explore the disease-related phenotypes in G2R398W/+ mice, we analyzed hematopoietic features in aged adult mice. To this end, we first examined the hematological phenotypes of G2R398W/+ mice at 12 months compared to those of G2+/+ siblings.
In the evaluation of peripheral blood counts, we found that the white blood cell (WBC) count was significantly decreased in G2R398W/+ mice at 12 months compared with that in G2+/+ mice, while the red blood cell (RBC) and platelet counts were maintained in the normal range (Fig. 3a). Notably, BCs, NKCs, myeloid DCs (mDCs), and plasmacytoid DCs (pDCs) were significantly reduced in number in G2R398W/+ mice (Fig. 3b). Monocytopenia was also observed in the G2R398W/+ mice, but the numbers of CD4 single-positive T cells (CD4+ TCs), CD8 single-positive T cells (CD8+ TCs) and granulocytes in mutant mice were comparable to those in control mice and were maintained within the normal range (Fig. 3b). These phenotypes of the G2R398W/+ mice nicely recapitulate the clinical observations of the diseases caused by heterozygous GATA2 mutations, including R398W29, and indicate that the G2R398W/+ mice serve as an excellent disease model mouse line.

a Summary of hematopoietic indices. A dot plot of WBC counts in the peripheral blood of G2+/+ and G2R398W/+ mice is shown in the right panel. b Dot plots of the cell counts of the indicated cell populations of G2+/+ and G2R398W/+ mice. *p < 0.05, **p < 0.01. N = 10 of each genotype.
To verify this conclusion, we next analyzed younger generations of G2R398W/+ mice. To this end, we exploited 5- to 7-month-old and 3-month-old G2R398W/+ mice and examined hematopoietic indices of the peripheral blood. We found similar decreases in the numbers of WBCs, BCs, NKCs, mDCs, and pDCs in 5- to 7-month-old G2R398W/+ mice, but monocytopenia was not observed in the mice (Supplementary Fig. 2). Decreases in WBCs, BCs, NKCs, mDCs, pDCs, and monocytes were not observed in 3-month-old G2R398W/+ mice (Supplementary Fig. 3). The numbers of CD4+ TCs, CD8+ TCs, and granulocytes in the G2R398W/+ mice were comparable to those in control mice at both the 5-7-month and 3-month time points. We further examined the second line of G2R398W/+ mice and found similar decreases in WBC count and WBC subpopulations in the G2R398W/+ mouse line at 6 months of age (Supplementary Fig. 4).
Partial decrease in hematopoietic stem and progenitor populations in G2 R398W/+ mice
As loss-of-function germline GATA2 mutations have been found in myelodysplastic syndrome and leukemias in humans29, we also examined bone marrow progenitors in G2R398W/+ mice. We analyzed the hematopoietic stem and progenitor cells of 12-month-old mice and found that the frequency of LSK in the G2R398W/+ mice was comparable to that in the G2+/+ siblings (Fig. 4a). In contrast, when we examined the LSK subpopulations, we found that the frequency of CD150-CD48-LSKs was significantly decreased (Fig. 4b).

a Quantification of LSKs in the bone marrow of G2+/+ and G2R398W/+ mice. b, c Quantification of stem cell populations subdivided according to CD150 and CD48 profiles (b) and progenitor populations (c) in the bone marrow of G2+/+ and G2R398W/+ mice. *p < 0.05. N = 10 of each genotype.
We also examined the frequencies of myeloid-restricted progenitors, e.g., common myeloid progenitors (CMPs), granulocyte-macrophage progenitors (GMPs), and megakaryocyte-erythroid progenitors (MEPs). The frequencies of these progenitors did not change between G2R398W/+ and G2+/+ mice (Fig. 4c). During the course of the 12-month follow-up period, we carefully checked for the onset of leukemia, but we could not detect the development of hematological neoplasms in the G2R398W/+ mice. Thus, these results imply that while heterozygous status for the G2R398W allele affects the differentiation and/or maintenance of hematopoietic progenitors, this influence is not strong enough to drive the development of hematopoietic malignancies.
The G2 R398W/+ hematopoietic phenotype does not exist in G2 Gfp/+ mice
We next examined whether the hematopoietic phenotypes observed in the G2R398W/+ mice could be seen in heterozygous GATA2-knockout mice. For this purpose, we used the Gfp knockin/Gata2 knockout mouse line at the 6-7-month timepoint12. We found that the analyses of peripheral blood hematopoietic indices and cell populations in the heterozygous Gata2-knockout (G2Gfp/+) mice generated results that stood in stark contrast to those of the G2R398W/+ mice.
The WBC numbers were comparable between the mice of these two genotypes (Fig. 5a), in good agreement with a previous report34. The numbers of BCs, NKCs, mDCs, pDCs, and monocytes did not change substantially (Fig. 5b). Therefore, these results support the hypothesis that the heterozygous G2R398W mutation-induced hematological phenotypes, which are substantially different from those induced by the heterozygous GATA2-null mutation.

a Summary of hematopoietic indices. A dot plot of WBC counts in the peripheral blood of G2+/+ and G2R398W/+ mice and their G2+/+ littermates is shown in the right panel. b Dot plots of the cell count of the indicated cell population in the peripheral blood of G2+/+ and G2R398W/+ mice. *p < 0.05, N = 4–5 of each genotype.
GATA2 dimerizes regardless of mutations at residues R398 and T354
The DNA-binding activity and transcriptional activity of G2R398W have been reported to be significantly reduced31. Therefore, we initially assumed that the mutated G2R398W allele would be equivalent to a null allele, but the comparison of the loss-of-function phenotypes in the previous subsection revealed that the G2R398W/+ mice showed specific phenotypes that are not found in the G2Gfp/+ mice (Fig. 5), suggesting that the heterozygous expression of the G2R398W protein may interfere with the functions of the intact GATA2 protein by a physical interaction. In this regard, it is interesting to note that two GATA1 molecules form a functional homodimer via reciprocal interactions of the CF and NF domains of each monomer37,38,39. Similarly, GATA3 also forms a homodimer, but in this case, the CF domains interact with each other40. Therefore, first performed a pull-down assay using maltose-binding protein (MBP)- and glutathione S-transferase (GST)-fused recombinant proteins covering the GATA2 finger region containing the NF and CF domains (from 289 aa to 480 aa) (Fig. 6a). We found that GST-fused G2WT was successfully pulled down by MBP-fused G2WT but not by MBP alone (Fig. 6b and Supplementary Fig. 5a), indicating that mutant and wild-type GATA2 physically interact with each other and form a homodimer.

a Schematic illustration of recombinant MBP-fused and GST-fused GATA2 proteins and the positions of the substituted amino acids. b–d Pull-down assays of MBP-fused (b, c) and GST-fused (d) GATA2. The indicated recombinant proteins were mixed. MBP- and GST-fused GATA2 were detected by the corresponding antibodies. White and black arrowheads indicate the pull-down signals of MBP-fused and GST-fused GATA2, respectively. Quantification of band intensities in Western blots was performed by the ratio of the GST (c) or MBP (d) pull-down signal to the input signal. The value in the left-most lane was set to 1.0. Experiments were performed in duplicate, and the results were reproducible.
To verify whether the R398W and T354M mutations affect the association of GATA2, we constructed GST-fused and MBP-fused G2R398W and G2T354M (Fig. 6a). G2T354M is another germline GATA2 mutation found in patients with DCML deficiency and MonoMAC syndrome patients29. We confirmed that both mutant proteins were pulled down by MBP-fused G2WT, and the efficiencies were comparable between G2WT and these two mutant proteins (Fig. 6c and Supplementary Fig. 5b). We further examined whether MBP-fused G2R398W and G2T354M could be pulled down by GST-fused G2WT. The results clearly demonstrated that the G2R398W and G2T354M proteins were both pulled down by GST-fused G2WT (Fig. 6d and Supplementary Fig. 5c), as was the case for the MBP-fused G2WT experiments. These results thus indicate that the G2R398W and G2T354M proteins have the capacity to dimerize with the G2WT protein and that the homodimerization of G2R398W and G2WT may underlie the pathological basis of the observed phenotypes of the G2R398W/+ mice.
Both G2R398W and G2T354M impair the binding of G2WT to the tandem GATA motif
The GATA1 monomer or single GATA1 molecule monovalently binds to the single GATA motif, while the GATA1 homodimer binds bivalently to the tandem GATA motif in which two GATA-binding motifs align in a direct-repeat arrangement using the CF domains of each molecule39. Since the homodimerization ability of GATA2 was not changed by the R398W and T354M substitution mutations, we examined whether the DNA-binding ability of G2R398W and G2T354M to the single GATA motif and/or G2R398W-G2WT and G2T354M-G2WT heterodimers to the tandem GATA motif was affected.
To this end, we prepared the recombinant proteins G2WT, G2R398W, and G2T354M and conducted surface plasmon resonance (SPR) analyses of the G2R398W and G2T354M monomers binding to the single GATA motif and G2R398W-G2WT and G2T354M-G2WT heterodimers binding to the tandem GATA motif.
As outlined in Fig. 7a, we first examined the binding of the G2WT monomer to a 21-mer DNA containing the single GATA motif. We found that the sensorgram curve of G2WT monomer binding to the single GATA motif showed a very rapid increase, and the binding reached equilibrium within a short time depending on the protein concentrations. In contrast, the binding of G2R398W and G2T354M was weak and took time to reach equilibrium under the conditions tested (Fig. 7b). The dissociation constant (KD) values of G2R398W and G2T354M were approximately 10-fold and 4-fold lower than those of G2WT, respectively (Fig. 7b). These results indicate that the DNA-binding strength of G2R398W and G2T354M was significantly affected by the substitution mutations.

SPR analyses of GATA2 binding to single (a–c) and tandem (d–f) GATA motifs. Schematic diagrams of the binding model of G2WT to single and tandem GATA motifs are shown on the right in a, d, respectively. The influences of the flow rate of the respective GATA2 proteins at the indicated protein concentrations are shown on the left. Kinetic parameters of the respective GATA2 proteins are shown in the tables. A mixture of equal amounts of the G2WT and G2mutant proteins was used in c, f. Experiments were performed in duplicate, and the results were reproducible.
We then analyzed the binding of a 1:1 mixture of G2WT:G2R398W and G2WT:G2T354M to oligonucleotides containing the single GATA motif. We found that although the ka values of the mixture were reduced by one order compared to those of G2WT alone, high-quality sensorgrams were obtained (Fig. 7c). Furthermore, the fit of the data was well correlated with the monovalent binding model. We also found that the kd values of the mixtures were reduced by one order compared to that of G2WT alone; consequently, the KD values were almost equivalent or within a less than twofold difference between G2WT alone and the mixtures of G2WT:G2R398W and G2WT:G2T354M (Fig. 7c). This indicates that wild-type GATA2 shows a comparable affinity for the single GATA motif regardless of the coexistence of mutant GATA2.
We next performed SPR analysis using 21-mer DNA containing the tandem GATA motif. We found that the obtained parameters for G2WT were not well fitted to the monovalent binding model. The parameters predicted that the binding fit the two-phase interactions (Fig. 7d). These results indicate that G2WT binds to the tandem GATA motif in a bivalent manner, similar to the case of GATA139.
Compared to the binding of G2WT, the binding of G2R398W and G2T354M was weak, and it took time to reach equilibrium under the conditions tested (Fig. 7e). The KD values of G2R398W and G2T354M were approximately 8-fold and 2.5-fold lower than those of G2WT, respectively (Fig. 7e). These results indicate that the DNA-binding strength of G2R398W and G2T354M to the tandem GATA motif was significantly affected by the substitution mutations. These changes were similar to those observed in the case of the single GATA motif.
Most importantly, when we used a 1:1 mixture of G2WT:G2R398W and G2WT:G2T354M, 2.3- and 1.3-fold decreases in the ka values of the first phase of interaction (ka1) and 1.7- and 2-fold increases in kd values of the first phase of interaction (kd1) were observed, respectively. Compared to that of the G2WT homodimer, these alterations in the KD values of the first phase of interaction (KD1) were increased by 4- and 3-fold, respectively (Fig. 7f). These decreases in binding affinity for the tandem GATA motif stood in clear contrast to the situation for the single GATA motif. These results thus support our hypothesis that the heterodimer of the G2mutant (G2R398W or G2T354M) markedly lost its DNA-binding ability. The coexistence of G2R398W or G2T354M with G2WT in the heterodimer seems to strongly disturb the ability of the G2WT:G2mutant heterodimer to bind to the tandem GATA motif.
The G2WT:G2mutant heterodimer loses its transactivation activity in the tandem GATA motif
To evaluate how the coexistence of the GATA2 mutant in the heterodimer affects the transcriptional activity of the G2WT:G2mutant heterodimer, we conducted luciferase reporter assays by using HEK293T cells. In particular, we focused on the transcriptional activity of GATA2 via the tandem GATA motif. For this purpose, we constructed three luciferase reporters containing single or two types of tandem GATA motifs (AGATAAGATAA-type and AGATAAAGATAA-type) placed in triplicate on the upstream side of the minimal promoter-driven Luc2CP gene and performed transient transfection assays.
When transfected individually with the single GATA and two types of tandem GATA reporters, the transactivation activities of G2R398W and G2T354M were significantly reduced compared with that of G2WT, regardless of the configuration of GATA-binding motifs (Fig. 8a–c). It is worth mentioning that the transactivation activities of G2R398W and G2T354M in the single GATA motif were increased, albeit slightly, in a dose-dependent manner, and the activity at a dose of 300 ng became equivalent to that of G2WT at a dose of 100 ng (Fig. 8a). In contrast, a 3-fold increase in the transfected constructs of G2R398W and G2T354M could not compensate for the decrease in transactivation activity in the tandem GATA motif (Fig. 8b, c). Thus, while functional defects caused by structural mutations can be partially compensated by an increase in quantity, the transactivation activity in the tandem GATA motif requires many more constructs than the single GATA motif.

a–c Scatter dot plots of the transcriptional activity measured by luciferase reporter assay in HEK293T cells following transfection with luciferase reporter constructs containing single (a) and tandem (b, c) GATA motifs together with the respective GATA2 expression constructs at doses of 100, 200, and 300 ng/well. AGATAAGATAA-type (middle) and AGATAAAGATAA-type (bottom) tandem GATA constructs were used for transient reporter/effector cotransfection assays. Upper and middle panels, n = 5 biologically independent samples and bottom panel, n = 4. d–f Similar sets of experiments in which 100 ng of the G2WT expression construct and luciferase reporter construct containing single (d) and tandem (e, f) GATA motifs were concomitantly transfected with G2WT, G2R398W, or G2T354M expression constructs at doses of 0, 100, and 200 ng/well. AGATAAGATAA-type (middle) and AGATAAAGATAA-type (bottom) tandem GATA constructs were used for transient reporter/effector cotransfection assays. Upper and middle panels, n = 5 biologically independent samples and bottom panel, n = 8. The average luciferase activity of G2WT at a dose of 100 ng/well was set to 1.0 in a–c, and fold changes of the average value of 100 ng of G2WT are shown in d–f. *p < 0.05.
To elucidate how coexpression of G2R398W and G2T354M influences the transactivation activity of G2WT, we performed a transfection assay again by using 100 ng of G2WT as the baseline amount. We then added an equal or twofold higher amount of either G2WT, G2R398W, or G2T354M to the basal 100 ng of G2WT. When we used the single GATA motif reporter, the luciferase activity was increased in a dose-dependent manner in all G2WT, G2R398W, and G2T354M cases (Fig. 8d). In this assay, the reporter activity at the basal 100-ng G2WT addition was set as 0. Compared with the incremental increase in luciferase reporter activity with the addition of the G2WT expression plasmid, the effects of G2R398W and G2T354M additions were fairly small.
In contrast, when we used the tandem GATA motif reporters, the addition of G2R398W and G2T354M to G2WT showed reverse dose-dependent effects against the transactivation activity of G2WT. While the addition of G2WT caused an increase in transactivation activity in a dose-dependent manner, both GATA2 mutants appeared to repress their transactivation activities at two conditions, i.e., with 25-ng AGATAGATAA-type tandem-GATA reporter (Fig. 8e) and 5-ng AGATAAGATAA-type tandem-GATA reporter (Fig. 8f). G2T354M worked as a less effective activator of the single GATA motif than G2R398W, but it worked as a more effective repressor of the tandem GATA motifs. Interesting observation here is that, effect size of GATA2 mutants on AGATAAGATAA-type tandem-GATA was smaller than that on AGATAGATAA-type tandem-GATA (Fig. 8e, f). Indeed, when we used 25-ng AGATAAGATAA-type tandem-GATA reporter, the dose-dependent adverse effect by coexisting G2R398W on the transactivation activity of G2WT was modest and not so significant (Supplementary Fig. 6). We speculate that, when effector/reporter ratio is low in a transient reporter assay, a single GATA2 molecule is prone to bind to the tandem GATA motif, especially in case of tandem GATA motif with multiple spacer nucleotides, while GATA2 dimers are prone to be produced and bind to the tandem GATA motif when the concentration of GATA2 molecules becomes high. Thus, leverage over the coexisting G2WT varied in accordance with the nature of the mutant GATA2, the configuration of GATA-binding motifs and probably with the expression level of GATA2 during hematopoiesis.
To further examine the negative effect of the GATA2 mutant on the transactivation activity of wild-type GATA2 from the chromatin integrated reporter, we prepared two types of cells stably carrying the reporter constructs of single or AGATAAGATAA-type tandem GATA motif and introduced GATA2 expression plasmids by episomal transfection. Showing strong agreement with the results of transient transfection reporter assays, the expression of G2R398W and G2T354M repressed the transcriptional activity of G2WT in a dose-dependent manner in the genome-integrated tandem motif but not in the single motif (Supplementary Fig. 7). These results further support the conclusion that these GATA2 mutants repress wild-type GATA2 activity in the heterodimer.
Fluctuating expression of genes in G2 R398W/+ LSKs
We next verified whether coexpressing G2R398W altered the expression of GATA2 target genes that contain the tandem GATA motif in the gene loci. To this end, we analyzed a GATA2 chromatin immunoprecipitation sequencing (ChIP-Seq) dataset of murine bone marrow lineage-negative cells obtained from the ChIP-Atlas database. A consensus GATA-binding motif was enriched in the peaks of 2193 ChIP-Seq peaks with a high enrichment value (E-value; Fig. 9a). These ChIP-Seq peak loci contain several types of tandem GATA motifs that contain various lengths of spacers (n = 0–5). In Fig. 9b, the identified tandem GATA motifs were aligned by using the E-values. Through this inspection of the 2193 ChIP-Seq peaks, we extracted 314 peaks that contained at least one tandem GATA motif with significant enrichment. We identified 103 genes that harbor tandem GATA motif(s) within ±100 kbp of the transcription start site (TSS) (Supplementary Table 1).

a GATA2-binding motif in ChIP-Seq peak regions in the GATA2 ChIP-Seq dataset of murine bone marrow lineage-negative cells. b Tandem GATA motifs were used to search ChIP-Seq peak regions containing tandem GATA motifs. Motifs are arranged in ascending order according to the E-value. The right panel shows the distance distributions between ChIP-Seq peak summits and the center of the motifs with color-matched lines. c–e Dot plots for the expression of genes containing the tandem GATA motif(s) in bone marrow LSK cells of 5- to 6-month-old mice. Genes commonly found in two datasets (c), found in the ENCODE dataset (d), and found in the ChIP-Atlas dataset (e) are shown. Tandem GATA motif types found in the ChIP-seq peaks in individual genes are shown. f Dot plots for the expression of genes harboring only a single GATA motif(s) in their proximity in bone marrow LSK cells of 5- to 6-month-old mice. *p < 0.05. Numbers of mice are indicated in the figure.
We further analyzed the ENCODE GATA2 ChIP-Seq dataset of G1E cells41 and found 39 genes containing tandem GATA motif(s) (Supplementary Table 1). Ten genes overlapped in two different datasets.
We then performed qRT-PCR analysis and confirmed the expression of 4 genes out of 10 in bone marrow LSK cells. In particular, the expression of Gfi1 and Smad5, both of which have been shown to be important hematopoietic regulators42,43, was significantly increased in the G2R398W/+ mice but not in the G2Gfp/+ mice (Fig. 9c). In addition, we chose the Eef2k gene as a tandem GATA motif-containing gene from the ENCODE dataset since Eef2k is known to regulate HSC apoptosis44 and found changes in expression in G2R398W/+ mice (Fig. 9d). Furthermore, we chose the Bcor, Jdp2, Gtpbp1, Acvrl1, Limk2, and Rasgrp4 genes from the ChIP-Atlas database based on their involvement in hematopoiesis and/or leukemogenesis45,46,47,48,49,50 and found that the expression of the Bcor and Jdp2 genes was significantly elevated only in the G2R398W/+ mice but not in the G2Gfp/+ mice (Fig. 9e). Genes whose expression was changed in both G2R398W/+ and G2Gfp/+ mice or only in G2Gfp/+ mice were also identified (Fig. 9e).
We next analyzed the expression of genes that have a single GATA motif but without any tandem GATA motifs in proximity. We chose the Tal1, Stat3, Tmem56, Zfpm1, and Il6ra genes, since they were reported as downstream genes of GATA23,30,36,51,52. While we could not find any genes whose expression was changed solely in the G2R398W/+ mice, we could identify genes changed only in G2Gfp/+ mice (Fig. 9f). Taken together, our data suggest that heterozygous expression of G2R398W affects GATA2 regulatory effects through highly inactive heterodimer formation, which preferentially affects the tandem GATA motif-mediated regulation of GATA2.
Discussion
Orchestrated and elaborate regulation of hematopoiesis-associated genes is critical for the maintenance of hematopoietic homeostasis, and it has been shown that the transcription factor GATA2 plays a central role in the maintenance of hematopoietic homeostasis14,15,16,17,18,19,20,34. Recent advances in clinical research have led to an emerging theory that the GATA2 gene is causative for congenital hematopoietic diseases in autosomal dominant traits, including Emberger syndrome, DCML deficiency, and MonoMAC syndrome. However, since simple heterozygous Gata2-knockout mice did not show phenotypes recapitulating these human diseases16,34, we surmise that the pathogenesis of GATA2-related immunodeficiency disease includes mechanisms much more complex than a simple quantitative deficit. To address such mechanisms, it seems necessary to develop more elaborate mouse models. To this end, we generated mouse lines carrying a heterozygous single-nucleotide substitution mutation (c.1192C>T, p.398Arg>Trp) in the Gata2 gene by a genome editing strategy in this study. This mutation was found in a family with DCML deficiency. We found that, in contrast to the heterozygous GATA2-null (G2–/+) mutant mice, the G2R398W/+ mice had significantly decreased numbers of BCs, NKCs, mDCs/pDCs and monocytes, but the numbers of both CD4+ TCs and CD8+ TCs as well as granulocytes were comparable to those of the control mice. Thus, the phenotypes of the G2R398W/+ mice nicely recapitulate the clinical observations of the diseases caused by the heterozygous GATA2 mutation, demonstrating that the G2R398W/+ mice serve as an excellent disease model mouse line.
Mutations found in GATA2-related diseases are divided into two categories29. One is quantitative deficits of the GATA2 protein, while the other is qualitative defects of the GATA2 protein. In the former case, a massive reduction in GATA2 gene expression usually occurs, and only a low level of the GATA2 protein is expected to reside in the cells, suggesting that a reduced amount of intact GATA2 protein is involved in the pathogenesis of these diseases. In contrast, in the latter cases, the mutated alleles produce structural mutants of GATA2 that coexist with intact GATA2 from the other wild-type allele. Most of the mutations in this category are concentrated in the CF domain of GATA229, severely impairing the DNA-binding activity of GATA231,53. As summarized in Fig. 10, we demonstrated in this study that two of the latter GATA2 mutants (p.398Arg>Trp and p.354Thr>Met) harbor weak binding activity to single and tandem GATA motifs. However, the coexistence of these GATA2 mutants significantly affected the function of coexisting wild-type GATA2 specifically through tandem GATA motifs, although the effect size of GATA2 mutants varied in accordance with the nature of tandem GATA motif, whereas the pristine function of wild-type GATA2 through the single GATA motif was not substantially altered by the coexistence of the GATA2 mutant. Based on these observations, we propose that the expression of GATA2 target genes, which is heavily dependent on the tandem GATA motif, is markedly influenced by the presence of heterozygous missense mutations, which results in the disorganized regulation of GATA2 and disruption of hematopoietic homeostasis, leading to the onset of DCML deficiency.

G2WT and G2mutant coexist in cells of affected individuals carrying heterozygous GATA2 gene mutations that produce GATA2 structural mutants. In cases of the G2mutant with impaired DNA-binding activity to the CF domain, the G2mutant effectively disrupts the function of G2WT on the tandem GATA motif and is barely involved in the function of G2WT on the single GATA motif. Genes whose regulation is heavily dependent on the tandem GATA motif are predicted to be subject to strong interference by the presence of the G2mutant.
We identified several genes that showed significant differences in expression levels between G2R398W/+ and G2+/+ mice through our qRT-PCR analysis coupled with ChIP-Seq database analysis. The expression of these genes was not changed between G2Gfp/+ and G2+/+ mice. These results suggest that the products of those genes may be responsible for the phenotypes of the qualitative defect of GATA2. In particular, Smad5 has been shown to be a GATA2-target gene and a key regulator of BMP signaling52,54,55, which supports HSC heterogeneity and lineage output decisions in the myeloid and lymphoid axes56. We envisage that perturbations in the expression of these GATA2 target genes may disturb the proliferation and differentiation of hematopoietic lineage cells.
We generated compound heterozygous mutant mice (G2R398W/Gfp) utilizing mice harboring the G2Gfp knockin-knockout allele12 and G2R398W allele. This G2R398W/Gfp mouse carries G2R398W and G2Gfp alleles so that GATA2 from the single G2R398W allele supports the function of GATA2 in the mice. We found that the G2R398W/Gfp mice survived at E10.5, which was beyond the critical lethal point of complete GATA2-knockout embryos11. We also found that G2R398W homozygotes (G2R398W/R398W) survived beyond E12.5, which is markedly longer than G2R398W/Gfp mice. However, the G2R398W/R398W embryos are not alive at the time of birth. In a reporter cotransfection-transactivation assay, we showed that G2R398W retains some extent of its transactivation activity. We surmise that the residual transcriptional activity of G2R398W contributes to the prolonged survival of G2R398W/R398W and G2R398W/Gfp embryos beyond the critical period in an allele number-dependent manner.
The G2R398W/+ mice showed a phenotype recapitulating that of DCML deficiency, but the mice never developed hematopoietic neoplasms. In contrast, mice heterozygous for the Gata2-null mutation (G2-/+) did not show an obvious phenotype except for a large number of defective hematopoietic stem and progenitor cells16,34. The expression of the Gata2 gene in the LSK cells of G2-/+ mice was reduced by approximately 50% of the wild-type level34 contrast, G2fGN/fGN mice that harbor approximately 20% of the GATA2 expression of wild-type mice are prone to develop progressive myelomonocytosis resembling CMML without leukocytopenia36. These observations revealed that the 20% expression level of GATA2 in the mice cannot fully explain human GATA2-related diseases. While quantitative deficits of GATA2 are indeed involved in the pathogenesis of MPNs, this study unequivocally demonstrates that the GATA2-related diseases provoked by heterozygous substitution mutations utilize specific mechanisms linked to qualitative deficits.
In conclusion, we identified in this study that mice harboring a heterozygous GATA2 mutant with weakened DNA-binding activity develop multilineage cytopenia of BCs, NKCs, mDCs, pDCs, and monocytes due to interference with coexisting wild-type GATA2 activity in a GATA motif configuration-dependent manner. Mutant GATA2 interferes with wild-type GATA2 activity via a dominant-negative mechanism. While in human patients, GATA2 structural mutations were scattered over a wide area of the GATA2 protein, we believe that the mechanism clarified in this study explains one type of GATA2-related disease. Thus, our G2R398W/+ mice will serve as a unique and vital model to study qualitative deficits of GATA2.
Methods
Animal studies
Experimental procedures for animals were approved by the Institutional Animal Experiment Committee of Tohoku University. Experiments were carried out in accordance with the Regulation for Animal Experiments at Tohoku University. The plasmid vector pX330 expressing Cas9 and gRNA was digested with BbsI, and a pair of DNA oligos recognizing the Gata2 target site (5′-AGG GAT CCA GAC CCG GAA T −3′) was ligated to the linearized vector. Donor DNA oligos encoding an arginine to tryptophan substitution were designed as follows: 5′-ACC ATG AAG AAG GAA GGG ATC CAG ACC CGG AAT TGG AAG ATG TCC AGC AAA TCC AAG AAG AGC AAG AAA G-3′ (the nucleotide corresponding to the c.1192 C > T substitution is underlined). The plasmid and donor DNA were coinjected into BDF1-fertilized eggs. The founder mice were crossed with C57BL/6 J mice for at least 6 generations, and progeny were used in this study. Primer sequences for genotyping PCR are listed in Supplementary Table 2. Images of embryos and fetal livers were captured with an MZFL III stereomicroscope (Leica) and a DP73 CCD camera (Olympus) using the manufacturer’s standard software. Blood samples were obtained from the retro-orbital plexuses of mutant mice and their unaffected littermates, and hematopoietic indices were measured with a Celltac-α autohemocytometer (Nihon Koden).
Flow cytometry analysis
Whole blood was hemolyzed with ammonium chloride hemolytic buffer and washed twice with phosphate-buffered saline. Lineage depletion of bone marrow cells was performed using a cocktail of biotinylated antibodies against Ter119, B220, Gr1, and CD8 (purchased from BioLegend) and CD4, CD11b and CD127 (eBioscience), followed by removal using Dynabeads M-280 streptavidin-conjugated magnetic beads (Thermo Fisher Scientific). The combinations of fluorescently labeled antibodies used in the comparison between G2R398W/+ and G2Gfp/+ mice with G2+/+ mice are shown in Supplementary Tables 3 and 4, respectively. The stained cells were analyzed and sorted by means of BD FACSAria II and BD FACSDiva software (Becton Dickinson). BCs, NKCs, mDCs, pDCs, monocytes, CD4+ TCs, and CD8+ TCs and granulocytes were defined as B220+CD19+CD4–CD8–, CD49b+NK1.1+NKG2D+CD11b+CD1d–CD3–, CD11b+CD11c+F4/80–, CD11b–CD11c+F4/80–Gr1+, CD11b+CD11c–F4/80+Gr1–, B220–CD19–CD4+CD8–, and B220–CD19–CD4–CD8+, respectively. Absolute numbers of each cell subset were calculated by the following formula: absolute count/μl = WBC count (/μl) x percentage of cells in total mononuclear cells determined by flow cytometry. Long-term (LT)-HSCs, ST-HSCs, MPP2s, MPP3s, and MPP4s were defined as CD150+Flk2–CD48–, CD150–Flk2–CD48–, CD150+Flk2–CD48+, CD150–Flk2–CD48+, and CD150–Flk2+CD48+, respectively, in the LSK-gated fraction57. Occasionally, LSK cells were subfractionated based on the expression of CD150 and CD48 into CD150+CD48–LSKs (HSCs), CD150–CD48–LSKs (MPPs), and CD150–CD48+LSKs (restricted progenitors)58. CMPs, GMPs, and MEPs were defined as CD34+CD16/32med, CD34low/+CD16/32high, and CD34lowCD16/32–, respectively, in the LK-gated fraction59.
Purification of recombinant protein and interaction analysis
MBP-fused and GST-fused murine G2WT, G2R398W, and G2T354M were produced in Escherichia coli BL21(DE3)pLysS-competent cells (Novagen)60, and the recombinant proteins were purified by affinity chromatography utilizing a Profinia instrument (Bio-Rad) according to the manufacturer’s protocol. For the pull-down assay, the recombinant MBP-fused proteins and each of the corresponding GST-fused proteins were incubated in binding buffer60,61 at room temperature. Protein complexes were purified with amylose magnetic beads (New England Biolabs) or glutathione magnetic beads (Thermo Fisher Scientific) and then detected by immunoblot analysis. SPR analysis was performed with a Biacore-X100 instrument (GE Healthcare). Double-stranded DNA probes in which the 5′ ends of sense strands were biotinylated were purified by native PAGE. Subsequently, each probe was immobilized in one of two streptavidin-coated flow cells in a Sensor chip-SA (GE Healthcare) as an active flow cell of 1000 resonance units (RU) DNA according to the manufacturer’s protocol. The other flow cell was left blank for reference subtraction. Data processing was performed with Biacore-X100 evaluation software (GE Healthcare). The sequences of the DNA probes were 5′-GCGCTCAGAGATAAGGCCTTG-3′ and 5′-GCGAGATAAGATAAGGCCTTG-3′.
Luciferase reporter analysis
For transient reporter/effector cotransfection assays, single GATA and two types of tandem GATA reporter constructs were generated by inserting fragments of TCCGGCAACAGATAAGGAATCCCTG-3′, 5′-TCCGGCAACAGATAAGATAAGGAATCCCTG-3′ or 5′-TCCGGCAACAGATAAAGATAAGGAATCCCTG-3′ in triplicate into the multiple cloning site of pGL4.28 vector (Promega). HEK293T cells (ATCC CRL-3216) were washed twice with phosphate-buffered saline (PBS) and resuspended in 500 μl of fresh medium at the concentration of 1.5 × 105/ml. Effector plasmids containing EF1α-promoter-driven murine G2WT and/or G2R398W or G2T354M expression vectors were adjusted in total amount to 300 ng with empty vector and transfected into HEK293T cells with reporter constructs. 25 ng of single GATA, 25 ng of AGATAAGATAA-type tandem GATA, and 5 and 25 ng of AGATAAAGATAA-type tandem GATA constructs were used for transient reporter/effector cotransfection assays. 0.75 ng of Renilla Luc expression vector was included in the transfections as an internal control. After 24 h of transfection, luciferase activity was measured at each dosage point of effector expression vectors with the Dual-Luciferase reporter assay system (Promega) with Lumat LB 9507 (Berthold Technologies) according to the manufacturer’s protocol. Data are expressed as the mean of duplicate or triplicate well from ≥4 independent experiments. For transient transfection assays, single GATA and tandem GATA reporter constructs were generated by inserting fragments of 5′-TCCGGCAACAGATAAGGAATCCCTG-3′ or 5′-TCCGGCAACAGATAAGATAAGGAATCCCTG-3′ in triplicate into the multiple cloning site of pNL1.3 reporter vector (Promega). HEK293T reporter cell lines carrying the single GATA or AGATAAGATAA-type tandem GATA reporter construct were established. Cells were washed twice with PBS and resuspended in 500 μl of fresh medium at the concentration of 1.5 × 105/ml followed by transfection with G2WT and/or G2R398W or G2T354M expression vector at the indicated dosages. The total amount of expression vector was adjusted to 300 ng with empty vector. Five microliters of culture medium was collected 24 h after transfection and used for measurement of luciferase activity with the Nano-Glo luciferase assay system (Promega) with the PHERAstar multiplate reader (BMG Labtech) according to the manufacturer’s protocol. Luciferase value was defined as the subtraction of luciferase activity measured in the cultured medium of untransfected cells. Date are expressed as the mean of triplicate well from ≥3 independent experiments.
Immunoblot analysis
Bone marrow cell nuclear extracts and a recombinant protein solution were mixed 1:1 with 2X Laemmli buffer and subjected to SDS-PAGE in a 10% polyacrylamide gel. The Precision Plus Protein Kaleidoscope Standard (Bio-Rad) was loaded into neighboring lanes as a molecular-weight marker. After electrophoresis, the separated proteins were transferred to PVDF membranes. The proteins were probed with primary anti-GATA2 (dilution ratio: 1:5000, Perseus Proteomics), anti-Lamin B (1:5000, Santa Cruz Biotechnology), anti-MBP (1:10,000, Santa Cruz Biotechnology), and anti-GST (1:5000, Santa Cruz Biotechnology) antibodies, followed by detection of the primary antibodies with horseradish peroxidase-conjugated secondary antibodies (1:5000-1:20,000, Invitrogen-Life Technologies). Signals were visualized on X-ray film using ECL-Prime Western blotting Detection Reagents (GE Healthcare) according to the manufacturer’s protocol. Signal values were quantified using ImageJ software.
Quantitative RT-PCR analysis
RNA was isolated from sorted cells using ISOGEN-LS (NIPPON GENE). Subsequently, first-strand cDNA was synthesized using ReverTra Ace (TOYOBO). Quantitative real-time PCR was carried out with a StepOnePlus Real-Time PCR System (Applied Biosystems) using THUNDERBIRD SYBR qPCR mix (TOYOBO) in triplicate wells for each reaction, and the results of the triplicate wells were averaged. The data were normalized to the expression level of Gapdh as an internal control. Primer sequences are described in Supplementary Table 5. Data for which relevant values were obtained from all mice are shown.
Analysis of GATA2 ChIP-Seq data
The GATA2 ChIP-Seq dataset (SRX035992) in murine bone marrow lineage-negative cells from the ChIP-Atlas database (https://chip-atlas.org/) and dataset (ENCSR000DIE) in the G1E cell line from the ENCODE consortium were analyzed. Motif enrichment analysis and motif searching were performed with the MEME-ChIP and FIMO programs in MEME Suite62, respectively. The discovered motifs were mapped to the reference genome using the Genomic Regions Enrichment of Annotations Tool (GREAT)63.
Statistics and reproducibility
In all, 4–10 mice for each genotype (i.e. WT, G2R398W/+, and G2Gfp/+) were used for flowcytometry, qRT-PCR, and immunoblot and hematopoietic indices analyses. 13 and 8 G2R398W/+ females were mated with G2R398W/+ and G2Gfp/+ males, respectively, and analyzed the genotypes of pups. Luciferase assays were performed using at least 4 biologically independent samples. Statistical analyses were performed with the Mann–Whitney U test. p-values were considered significant if p < 0.05. The pull-down and SPR analyses were performed in two technical replicates in two independent experiments.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Source data underlying Figs. 1d–g, 3a, b, 4a, b, 5a, b, 7a–f, 8a, b, 9c–f, S1a, b, S2a, b, S3a, b, and S4a, b are presented in Supplementary Data 1. Original source data for the Figs. 1c, g, and 6b–d are also presented in Supplementary Data 1. The GATA2 ChIP-Seq datasets in murine bone marrow lineage-negative cells and in the G1E cell line are available from the ChIP-Atlas database (accession code SRX035992) and from the ENCODE consortium (accession code ENCSR000DIE), respectively.
References
Yamamoto, M. et al. Activity and tissue-specific expression of the transcription factor NF-E1 multigene family. Genes Dev. 4, 1650–1662 (1990).
Fujiwara, T. et al. 2009 Discovering hematopoietic mechanisms through genome-wide analysis of GATA factor chromatin occupancy. Mol. Cell 36, 667–681 (2009).
Suzuki, M. et al. 2013 GATA factor switching from GATA2 to GATA1 contributes to erythroid differentiation. Genes Cells 18, 921–933 (2013).
Lowry, J. A. & Atchley, W. R. Molecular evolution of the GATA family of transcription factors: conservation within the DNA-binding domain. J. Mol. Evol. 50, 103–115 (2000).
Tsarovina, K. et al. Essential role of Gata transcription factors in sympathetic neuron development. Development 131, 4775–4786 (2004).
Khandekar, M. et al. A Gata2 intronic enhancer confers its pan-endothelia-specific regulation. Development 134, 1703–1712 (2007).
Hoshino, T. et al. Reduced BMP4 abundance in Gata2 hypomorphic mutant mice result in uropathies resembling human CAKUT. Genes Cells 13, 159–170 (2008).
Linnemann, A. K., O’Geen, H., Keles, S., Farnham, P. J. & Bresnick, E. H. Genetic framework for GATA factor function in vascular biology. Proc. Natl Acad. Sci. USA 108, 13641–13546 (2011).
Ainoya, K. et al. UG4 enhancer-driven GATA-2 and bone morphogenetic protein 4 complementation remedies the CAKUT phenotype in Gata2 hypomorphic mutant mice. Mol. Cell Biol. 32, 2312–2322 (2012).
Yu, L. et al. Reducing inflammatory cytokine production from renal collecting duct cells by inhibiting GATA2 ameliorates acute kidney injury. Mol. Cell Biol. 37, e00211–e00217 (2017).
Tsai, F. Y. et al. An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature 371, 221–226 (1994).
Suzuki, N. et al. Combinatorial Gata2 and Sca1 expression defines hematopoietic stem cells in the Bone Marrow Niche. Proc. Natl Acad. Sci. USA 103, 2202–2207 (2006).
Charles, M. A. et al. Pituitary-specific Gata2 knockout: effects on gonadotrope and thyrotrope function. Mol. Endocrinol. 20, 1366–1377 (2006).
Tipping, A. J. et al. High GATA-2 expression inhibits human hematopoietic stem and progenitor cell function by effects on cell cycle. Blood 113, 2661–2672 (2009).
de Pater, E. et al. Gata2 is required for HSC generation and survival. J. Exp. Med. 210, 2843–2850 (2013).
Rodrigues, N. P. et al. GATA-2 regulates granulocyte-macrophage progenitor cell function. Blood 112, 4862–4873 (2008).
Li, Y., Qi, X., Liu, B. & Huang, H. The STAT5–GATA2 Pathway is critical in basophil and mast cell differentiation and maintenance. J. Immunol. 194, 4328–4338 (2015).
Collin, M., Dickinson, R. & Bigley, V. Haematopoietic and immune defects associated with GATA2 mutation. Br. J. Haematol. 169, 173–187 (2015).
Onodera, K. et al. GATA2 regulates dendritic cell differentiation. Blood 128, 508–518 (2016).
Ohneda, K. & Yamamoto, M. Roles of hematopoietic transcription factors GATA-1 and GATA-2 in the development of red blood cell lineage. Acta Haematol. 108, 237–245 (2002).
Grass, J. A. et al. GATA-1-dependent transcriptional repression of GATA-2 via disruption of positive autoregulation and domain-wide chromatin remodeling. Proc. Natl Acad. Sci. USA 100, 8811–8816 (2003).
Kaneko, H., Shimizu, R. & Yamamoto, M. GATA factor switching during erythroid differentiation. Curr. Opin. Hematol. 17, 163–168 (2010).
Zhang, S. J. et al. Gain-of-function mutation of GATA-2 in acute myeloid transformation of chronic myeloid leukemia. Proc. Natl Acad. Sci. USA 105, 2076–2081 (2008).
Hahn, C. N. et al. Heritable GATA2 mutations associated with familial myelodysplastic syndrome and acute myeloid leukemia. Nat. Genet. 43, 1012–1017 (2011).
Ostergaard, P. et al. Mutations in GATA2 cause primary lymphedema associated with a predisposition to acute myeloid leukemia (Emberger syndrome). Nat. Genet. 43, 929–931 (2011).
Dickinson, R. M. et al. Exome sequencing identifies GATA-2 mutation as the cause of dendritic cell, monocyte, B and NK lymphoid deficiency. Blood 118, 2656–2658 (2011).
Hsu, A. P. et al. Mutations in GATA2 are associated with the autosomal dominant and sporadic monocytopenia and mycobacterial infection (MonoMAC) syndrome. Blood 118, 2653–2655 (2011).
Hsu, A. P., McReynolds, L. J. & Holland, S. M. GATA2 deficiency. Curr. Opin. Allergy Clin. Immunol. 15, 104–109 (2015).
Shimizu, R. & Yamamoto, M. Quantitative and qualitative impairments in GATA2 and myeloid neoplasms. IUBMB Life 72, 142–150 (2020).
Cortés-Lavaud, X. et al. GATA2 germline mutations impair GATA2 transcription, causing haploinsufficiency: functional analysis of the p.Arg396Gln mutation. J. Immunol. 194, 2190–2198 (2015).
Chong, C. E. et al. Differential effects on gene transcription and hematopoietic differentiation correlate with GATA2 mutant disease phenotypes. Leukemia 32, 194–202 (2018).
Wlodarski, M. W. et al. Prevalence, clinical characteristics, and prognosis of GATA2-related myelodysplastic syndromes in children and adolescents. Blood 127, 1387–1397 (2016).
Donadieu, J. et al. Natural history of GATA2 deficiency in a survey of 79 French and Belgian patients. Haematologica 103, 1278–1287 (2018).
Rodrigues, N. P. et al. Haploinsufficiency of GATA-2 perturbs adult hematopoietic stem-cell homeostasis. Blood 106, 477–484 (2005).
Ling, K. W. et al. GATA-2 plays two functionally distinct roles during the ontogeny of hematopoietic stem cells. J. Exp. Med. 200, 871–882 (2004).
Harada, N., Hasegawa, A., Hirano, I., Yamamoto, M. & Shimizu, R. GATA2 hypomorphism induces chronic myelomonocytic leukemia in mice. Cancer Sci. 110, 1183–1193 (2019).
Crossley, M., Merika, M. & Orkin, S. H. Self-association of the erythroid transcription factor GATA-1 mediated by its zinc finger domains. Mol. Cell Biol. 15, 2448–2456 (1995).
Mackay, J. P. et al. Involvement of the N-finger in the self-association of GATA-1. J. Biol. Chem. 273, 30560–30567 (1998).
Hasegawa, A. et al. GATA1 Binding kinetics on conformation-specific binding sites elicit differential transcriptional regulation. Mol. Cell Biol. 36, 2151–2167 (2016).
Bates, D. L., Chen, Y., Kim, G., Guo, L. & Chen, L. Crystal structures of multiple GATA zinc fingers bound to DNA reveal new insights into DNA recognition and self-association by GATA. J. Mol. Biol. 381, 1292–1306 (2008).
Weiss, M. J., Yu, C. & Orkin, S. H. Erythroid-cell-specific properties of transcription factor GATA-1 revealed by phenotypic rescue of a gene-targeted cell line. Mol. Cell. Biol. 17, 1642–1651 (1997).
van der Meer, L. T., Jansen, J. H. & van der Reijden, B. A. Gfi1 and Gfi1b: key regulators of hematopoiesis. Leukemia 24, 1834–1843 (2010).
Singbrant, S. et al. Smad5 is dispensable for adult murine hematopoiesis. Blood 108, 3707–3712 (2006).
Liao, Y. et al. Paradoxical Roles of Elongation Factor-2 Kinase in Stem Cell Survival. J. Biol. Chem. 291, 19545–19557 (2016).
Cao, Q. et al. BCOR regulates myeloid cell proliferation and differentiation. Leukemia 30, 1155–1165 (2016).
Baik, J. et al. Endoglin integrates BMP and Wnt signalling to induce haematopoiesis through JDP2. Nat. Commun. 7, 13101 (2016).
Senju, S., Iyama, K., Kudo, H., Aizawa, S. & Nishimura, Y. Immunocytochemical analyses and targeted gene disruption of GTPBP1. Mol. Cell. Biol. 20, 6195–6200 (2000).
Zhang, L. et al. Modulation of TGF-β signaling by endoglin in murine hemangioblast development and primitive hematopoiesis. Blood 118, 88–97 (2011).
Jensen, P. et al. Requirement for LIM kinases in acute myeloid leukemia. Leukemia 34, 3173–3185 (2020).
Watanabe-Okochi, N. et al. Possible involvement of RasGRP4 in leukemogenesis. Int. J. Hematol. 89, 470–481 (2009).
Thoms, A. I. J. et al. Disruption of a GATA2, TAL1, ERG regulatory circuit promotes erythroid transition in healthy and leukemic stem cells. Blood. 138, 1441–1455 (2021).
Tolkachov, A. et al. Loss of the hematopoietic stem cell factor GATA2 in the osteogenic lineage impairs trabecularization and mechanical strength of bone. Mol. Cell. Biol. 38, e00599–17 (2018).
Kazenwadel, J. et al. GATA2 is required for lymphatic vessel valve development and maintenance. J. Clin. Invest. 125, 2979–2994 (2015).
Suzuki, A., Chang, C., Yingling, J. M., Wang, X. F. & Hemmati-Brivanlou, A. Smad5 induces ventral fates in Xenopus embryo. Dev. Biol. 184, 402–405 (1997).
Marks-Bluth, J. et al. SMAD1 and SMAD5 expression is coordinately regulated by FLI1 and GATA2 during endothelial development. Mol. Cell. Biol. 35, 2165–2172 (2015).
Crisan, M. et al. BMP signalling differentially regulates distinct haematopoietic stem cell types. Nat. Commun. 6, 8040 (2015).
Pietras, E. M. et al. Functionally distinct subsets of lineage-biased multipotent progenitors control blood production in nor- mal and regenerative conditions. Cell Stem Cell 17, 35–46 (2015).
Ali, M. A. E. et al. Functional dissection of hematopoietic stem cell populations with a stemness monitoring system based on NS-GFP transgene expression. Sci. Rep. 7, 11442 (2017).
Akashi, K., Traver, D., Miyamoto, T. & Weissman, I. L. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 404, 193–197 (2000).
Kaneko, H., Kobayashi, E., Yamamoto, M. & Shimizu, R. N- and C-terminal transactivation domains of GATA1 protein coordinate hematopoietic program. J. Biol. Chem. 287, 21439–21449 (2012).
Shimizu, R. et al. GATA-1 self-association controls erythroid development in vivo. J. Biol. Chem. 282, 15862–15871 (2007).
Machanick, P. & Bailey, T. L. MEME-ChIP: motif analysis of large DNA datasets. Bioinformatics 27, 1696–1697 (2011).
McLean, C. Y. et al. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 28, 495–501 (2010).
Acknowledgements
We thank Drs Fumiki Katsuoka and Hiroshi Kaneko for helpful comments, and Satoshi Sagai, Daishi Ishihara, Aya Ashizawa, Hiromi Suda, and Tatsuro Iso for technical help. We also thank the Biomedical Research Core of Tohoku University Graduate School of Medicine for technical support. This work was supported in part by the Japan Society for the Promotion of Science (JSPS) KAKENHI (18K16074, A.H.; 19H03555, R.S.), the Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research) from AMED under Grant Number 20am0101095 (M.Y. & R.S.), and the Project for Cancer Research and Therapeutic Evolution from AMED under Grant Number JP20cm0106101 (M.Y.).
Author information
Authors and Affiliations
Contributions
A.H. and R.S. designed the study. A.H. and M.M. designed and established mutant mouse lines. A., Y.Hayasaka, Y.T., and Y.Hosaka were involved in the acquisition of data. A.H., Y. Hayasaka, and R.S. analyzed data. A.H., Y.Hayasaka, I.H., M.Y., and R.S. were involved in the interpretation of the data. A.H. wrote the draft. M.Y. and R.S. were involved in reviewing and editing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Matthew Collin and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: George Inglis.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hasegawa, A., Hayasaka, Y., Morita, M. et al. Heterozygous variants in GATA2 contribute to DCML deficiency in mice by disrupting tandem protein binding. Commun Biol 5, 376 (2022). https://doi.org/10.1038/s42003-022-03316-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-022-03316-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
