
Abstract
Central to the diversity of wheat products was the origin of hexaploid bread wheat, which added the D-genome of Aegilops tauschii to tetraploid wheat giving rise to superior dough properties in leavened breads. The polyploidization, however, imposed a genetic bottleneck, with only limited diversity introduced in the wheat D-subgenome. To understand genetic variants for quality, we sequenced 273 accessions spanning the known diversity of Ae. tauschii. We discovered 45 haplotypes in Glu-D1, a major determinant of quality, relative to the two predominant haplotypes in wheat. The wheat allele 2 + 12 was found in Ae. tauschii Lineage 2, the donor of the wheat D-subgenome. Conversely, the superior quality wheat allele 5 + 10 allele originated in Lineage 3, a recently characterized lineage of Ae. tauschii, showing a unique origin of this important allele. These two wheat alleles were also quite similar relative to the total observed molecular diversity in Ae. tauschii at Glu-D1. Ae. tauschii is thus a reservoir for unique Glu-D1 alleles and provides the genomic resource to begin utilizing new alleles for end-use quality improvement in wheat breeding programs.
Similar content being viewed by others


Introduction
Originating in the Fertile Crescent some 10,000 years ago, hexaploid wheat (Triticum aestivum) is now grown and consumed around the world1. The global consumption of wheat as a staple crop is owed principally to the unique viscoelastic properties of wheat dough that lend it the capacity to make diverse baked products such as leavened bread, tortillas, chapati, pastries, and noodles. The uniqueness of wheat dough can also be described as the strength to resist deformation and elasticity to recover the original shape as well as the viscosity to permanently deform under persistent stress. Elasticity is important for the product to hold shape, while viscosity allows the dough to be worked and formed. The balance of the competing properties determines what baked goods a dough is suitable for, such as a dough with greater strength for leavened pan bread compared to the more extensible dough that is desired for a chapati or tortilla.
Bread wheat is an allohexaploid with the A-, B- and D-subgenomes contributed by different, but related, species. The closest relative to the wheat A-subgenome is diploid Triticum urartu, with other diploid A-genome species including the wild and domesticated Einkorn wheat (Triticum monococcum). While the exact ancestor of the B-genome is unknown and presumed extinct, it is believed that Ae. speltoides (S-genome) is the closest living relative. These two species were brought together to form a tetraploid wheat species with AABB genome composition, which is known as durum or pasta wheat (Triticum durum). The D genome from Aegilops tauschii was the most recent addition forming a hexaploid wheat species. This addition of the D-subgenome led to a much broader adaptation and superior bread-making quality compared to the tetraploid and diploid ancestors2. However, the original hexaploid species originated from very few Ae. tauschii accessions and limited subsequent cross-hybridization likely caused by ploidy barriers with the diploid Ae. tauschii3. This genetic bottleneck resulted in limited genetic diversity in the wheat D-subgenome4.
Aegilops tauschii as a species is comprised of three subspecies, commonly referred to as lineages, that can be discerned through subtle phenotypic differences but more readily differentiated genetically by large variation among the lineages5,6. Lineage 1, Ae. tauschii spp. tauschii, is found across the native range of Ae. tauschii while Lineage 2, Ae. tauschii spp. strangulata is found primarily near the Caspian Sea in Iran and Azerbaijan5. Recent work has confirmed the existence of another lineage, Lineage 3, which is not yet designated by one of the historical subspecies names. While Lineage 2 is known to be a wheat D-subgenome donor, recent work has shown that Lineage 3 also contributed to the modern wheat genome6.
The utility of wheat, the variation of wheat products, and therefore consumption is driven by the strength and elasticity of the dough which is determined by the structure of the gluten matrix. This matrix is formed from a combination of high-molecular weight (HMW) and low-molecular weight (LMW) glutenin proteins and gliadins7. The backbone of the gluten matrix is developed during dough mixing by the covalent disulfide bonds between cysteine residues in HMW glutenins8. These glutenins, therefore, are some of the most important genes giving wheat its unique dough properties. They are encoded by a relatively simple locus on the long arm of the group one chromosomes of the Triticeae. Hexaploid wheat, comprised of the A-, B- and D-genomes, thus contains three HMW glutenin loci; Glu-A1, Glu-B1 and Glu-D1, respectively. Each locus harbors two HMW glutenin genes known as the x and y subunit, which are tightly linked but separated by tens to hundreds of kilobase pairs (kb)9,10,11. Each subunit consists of short, unique N and C terminal domains which flank a highly repetitive central region that accounts for 74–84% of the total protein length12.
Allelic differences in all three glutenin loci contribute to the conformation of the gluten matrix and variability in end-use quality. The D-subgenome locus, however, is the major driver of bread quality and the absence of the D-genome leads to substantially different dough qualities found in tetraploid pasta wheats13. The two common alleles at Glu-D1 found in bread wheat are Glu-D1a (SDS-PAGE allele designation 2 + 12) and Glu-D1d (5 + 10), with the latter being associated with superior bread-making quality14,15,16,17. Following the domestication and breeding of wheat, there is limited variation at the Glu-D1 locus in the D-genome with only these two alleles found in the vast majority of bread wheat throughout the world18,19. Thus, the addition of the wheat D-subgenome and specifically variation at Glu-D1 has a substantial impact on wheat quality globally. This is arguably the single greatest defining feature of bread wheat.
Reflecting the importance of Glu-D1 in determining the end-use quality of wheat, focus has been given to understanding the variation present in Ae. tauschii for this locus. Much of the work has utilized sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) protein analysis of Ae. tauschii collections20,21,22,23. From this work, over 37 SDS-PAGEGlu-D1 alleles have been named in Ae. tauschii. However, due to the limited resolution of SDS-PAGE, many of the alleles have indistinguishable SDS-PAGE mobilities from the common Glu-D1 hexaploid alleles, 2 + 12 and 5 + 10, or are difficult to reliably distinguish. By changing the polyacrylamide percentage or acidity in the SDS-PAGE, it was shown that the Ae. tauschii 2 + 12 and 5 + 10 alleles were slightly different than the common wheat alleles24. These Ae. tauschii alleles are therefore given the designations 2t + 12t and 5t + 10t. In addition to the 2t + 12t and 5t + 10t alleles, a large number of SDS-PAGE alleles have been described, supporting the hypothesis that Ae. tauschii could be a vast resource for untapped diversity at Glu-D1 and that this diversity could be utilized for wheat quality improvement.
Here we characterized the Glu-D1 allelic diversity in a panel of 273 sequenced Ae. tauschii accessions. The panel spans the known genetic diversity of Ae. tauschii and is a powerful resource for association mapping and gene identification6. From the sequenced Ae. tauschii panel, we discovered hundreds of genetic variants which defined dozens of unique haplotypes at Glu-D1. This gives the needed molecular information to track these alleles in breeding germplasm, which will, in turn, enable targeted assessment of the Ae. tauschii HMW glutenin alleles in hexaploid backgrounds leading to utilization of favorable alleles for wheat quality improvement.
Results and discussion
Molecular diversity of Glu-D1 in Ae. tauschii
Through the Open Wild Wheat Consortium, we obtained Illumina 150 bp paired-end short reads from 273 Ae. tauschii accessions, of which 234 were unique, each sequenced to greater than 7-fold coverage6. These were aligned to the Ae. tauschii ‘AL8/78’ reference genome and sequence variants at the annotated Glu-D1 locus were extracted for Glu-D1x at chr1D: 419306988-419309556 and Glu-D1y at chr1D: 419364015 -419365995 (Supplementary Data 2). We also included wheat cultivars in this analysis to compare Ae. tauschii variants to the common hexaploid wheat alleles 5 + 10 (varieties ‘CDC Stanley’ and ‘CDC Landmark’) and 2 + 12 (varieties ‘Chinese Spring’ and ‘LongReach Lancer’). From this panel, we identified a total of 308 variants at Glu-D1, which were used to generate haplotypes and evaluate molecular diversity at this locus.
From the Ae. tauschii germplasm collection we identified 32 and 33 haplotypes within the coding sequence for the x and y subunits of the Glu-D1 locus, respectively (Supplemental Data 1). When considering the complete Glu-D1 locus with a combination of the x and y subunit, a total of 45 haplotypes were identified. The various x and y subunit haplotypes were almost exclusively associated with each other, demonstrating the close physical association and limited recombination between the two genes. We included the 2500 bp up- and down-stream sequences of each subunit in our analysis to see if this resulted in further differentiation of alleles as short-read sequences often do not align uniquely to the central, highly repetitive region of the HMW glutenin genes. Including the flanking regions did not result in additional haplotypes. Thus, it appears that the identified variants are sufficient for reliably differentiating alleles at Glu-D1.
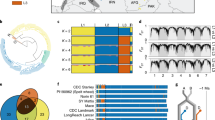
Gene-level phylogeny at Glu-D1 for all of the Ae. tauschii accessions revealed that haplotypes clustered into three major clades, two of which were associated predominantly with Lineage 2 and one with Lineage 1 (Fig. 1a). A unique group of Glu-D1 alleles from the newly characterized Lineage 3 accessions were found within a narrow clade with Lineage 2. Among the three major clades, we designated 15 subclades. Of the 15 subclades, eight were associated exclusively with Lineage 2, five with Lineage 1, and one with Lineage 3. The Lineage 3 accessions all fell within the Lineage 2 major-clades but occupied a unique subclade therein. Thus, the gene-level phylogeny at this locus agrees very closely with the previously described population structure of the Ae. tauschii lineages5,6. We also observed one clade (9) that had representative accessions from both Lineage 1 and 2. This could represent an ancestral haplotype found in both lineages which underwent incomplete lineage sorting, or a case of recent interlineage haplotype exchange. Cases of haplotypes shared across Lineages 1 and 2 were also observed for pest (Cmc4) and disease resistance (Sr46) genes6.

a Maximum likelihood tree based on 308 single-nucleotide variants found in Glu-D1 x and y subunits including 2.5 kb flanking regions on each side of the genes. The corresponding lineage of each accession is colored in yellow (Lineage 1), blue (Lineage 2), orange (Lineage 3), and green (bread wheat). Haplotypes with major-clades and subclades are designated by numbers. The wheat 2 + 12 allele in subclade 1 and the 5 + 10 allele is found in subclade 7. (https://itol.embl.de/export/17410641549711623435886). R1 and R2 designate putative recombinant Glu-D1 haplotypes in Ae. tauschii accessions TA1668 (R1) and TA2576 (R2). b Lineage-specific k-mers bins (100 Kb) as identified by Gaurav et al. anchored on genome assemblies of 11 wheat cultivar shown for Lineage 1 (yellow), Lineage 2 (blue), and Lineage 3 (orange). Arrows and black segments indicate the positions of Glu-D1 in each of the bread wheat genomes with 2 + 12 in blue and 5 + 10 in orange.
Lineage 2, the recognized ancestral diploid donor of the D-subgenome of hexaploid wheat3, had greater Glu-D1 molecular haplotype diversity than Lineage 1. Not only were there more subclades associated with Lineage 2, there were also more haplotypes. As expected, the haplotypes of wheat clustered within Lineage 2 subclades (Fig. 1). Within Lineage 2, we observed Ae. tauschii accessions with a matching sequence haplotype to the wheat 2 + 12 allele consistent with the D-subgenome origin from Lineage 2. Interestingly, the wheat 5 + 10 allele clustered within the Lineage 3 subclade indicating a unique origin of this allele. We then examined lineage-specifick-mer bin distributions as determined by Gaurav et al.6 and found that Lineage 3-specifick-mers co-colocalized with bread wheat 5 + 10 allele in the genomes of a set of 11 bread wheat cultivars (Fig. 1b), giving additional support that the superior 5 + 10 allele for bread wheat quality was derived from Lineage 3. These findings reveal that the Lineage 3 contribution to the wheat D-subgenome included the very valuable Glu-D1 5 + 10 allele, arguably one of the most important factors defining the quality of bread wheat.
Given the large difference in quality between wheat cultivars carrying 2 + 12 and 5 + 10 alleles, we hypothesized that these two haplotypes would not be similar at a molecular level. However, we found that 2 + 12 and 5 + 10 clustered relatively closely within major-clade III, with much greater overall diversity detected across Ae. tauschii particularly when including the Lineage 1 accessions which had very different haplotypes. When comparing the 2 + 12 and 5 + 10 haplotypes to those found in Lineage 1, it becomes apparent that Ae. tauschii carries alleles that are very unlike anything seen in bread wheat and may offer unique functional characteristics when introgressed into hexaploid backgrounds.
Geographic diversity
Given the known geographic structure and distribution of Ae. tauschii which is associated with various levels of population structure5, we evaluated the Glu-D1 diversity relative to the geographic origin of the Ae. tauschii accessions. Glu-D1major-clades II and III overlapped in regions around the Caspian Sea whereas major-Clade I spanned the entire geographic range (Fig. 2a). Molecular haplotypes were strongly associated with geographic origin, consistent with the overall genome-wide picture5, and genetic distances between alleles increased with the geographic distance between collection sites of the Ae. tauschii accessions (Fig. 2b). The greatest concentration of haplotype diversity was located along the shores of the Caspian Sea in Iran (Fig. 2b). Consistent with a hypothesis of admixture between Lineage 1 and Lineage 2 leading to shared gene-level haplotypes across the lineages, the accessions from Lineage 1 and 2 with the same Glu-D1 haplotype (within subclade 9) were collected very near one another.

Molecular haplotypes for Glu-D1 shown at the collection site for the given Ae. tauschii accession. a Distribution of accessions according to Glu-D1 major-clades with major-clade I in blue, major-clade II in red and major-clade III in orange. b Distribution of accessions according to Glu-D1 haplotype subclades. Haplotypes are shown on a scale from purple to yellow according to the phylogenetic tree (Fig. 1). Lineages are shown as squares for Lineage 1, circles for Lineage 2 and triangles for Lineage 3.
Molecular haplotypes identify novel Glu-D1 alleles
We employed SDS-PAGE analysis, the traditional standard for differentiating HMW glutenin alleles, to determine if the haplotype molecular sequence diversity would also reflect differences in protein mobility. We evaluated a total of 72 unique accessions with SDS-PAGE and differentiated 9 alleles for the x subunit and 8 alleles at the y subunit from this protein mobility assay. Analysis of the Lineage 1 and Lineage 2 variants revealed that molecular haplotypes were consistent with the proteins differentiated by SDS-PAGE (Supplemental Data 1). For the majority of the alleles that were differentiated by SDS-PAGE, we were able to unambiguously correlate the observed SDS-PAGE alleles with the molecular variants. Although specific molecular haplotypes were associated with specific SDS-PAGE mobilities, there was little concordance between gene-level variation and SDS-PAGE mobility as similar alleles at the molecular level were observed with very different SDS-PAGE mobilities. Alternatively, very different molecular haplotypes were observed with the same SDS-PAGE allele. This supports our hypothesis that the observed sequence variants are effectively in complete linkage disequilibrium and tagging the variation from the central repeat region. Similarly, the SDS-PAGE diversity was lower than that of the molecular haplotypes, thus SDS-PAGE has less power to differentiate alleles than the molecular haplotypes. As noted, the same SDS-PAGE mobilities were observed in both Lineage 1 and Lineage 2 haplotypes, but the molecular haplotypes were clearly differentiated (Fig. 1a and Supplemental Data 1). The protein mobility differences are considered to be primarily due to variation in the central repetitive region and therefore are not directly detectable with short-read sequencing, though the variable central repeats are completely phased with diagnostic haplotype variants within the terminal coding regions. Thus, we conclude that a sequence-based resource such as this Ae. tauschii panel provides a superior tool for the identification and tracking of unique Glu-D1 alleles in molecular breeding.
We also examined the connection between the glutenin protein mobility in Ae. tauschii compared to hexaploid wheat. Ae. tauschii haplotype Dx1a + Dy1a matched with the wheat 2 + 12 haplotype and exhibited the same SDS-PAGE mobility. Although we found an Ae. tauschii haplotype identical to the wheat 2 + 12 allele haplotype, the exact wheat 5 + 10 haplotype was not detected in this panel, however, a very closely related Lineage 3 haplotype was found. Additionally, no 5 + 10SDS-PAGE mobilities were observed. This was a surprising observation given that previous studies reported Ae. tauschii alleles with a 5 + 10SDS-PAGE mobility20. However, Williams et al.20 did not report the identities of the Ae. tauschii accessions with 5 + 10SDS-PAGE mobility. The haplotype Dx7a + Dy7a in the newly characterized Lineage 36 was most similar to 5 + 10 on the molecular level, however, it carried eight variant differences. This current panel, however, only has five unique accessions representing Lineage 3. It is likely therefore that exploration of additional Lineage 3 accessions could reveal a haplotype exactly matching the wheat 5 + 10 with the same mobility.
Cryptic haplotypes
One of the most valuable findings of this study was the high prevalence of cryptic molecular haplotypes hidden within SDS-PAGE mobilities. Within every SDS-PAGE mobility pattern, there were multiple molecular haplotypes, often from very different subclades and occasionally from entirely different major clades. The cryptic SDS-PAGE haplotypes, accordingly, were geographically disperse (Fig. 3). For example, within SDS-PAGE 2 + 12 were four haplotypes; one which was the same as wheat 2 + 12 (Dx1a + Dy1a), another which was within the same subclade (Dx1c + Dy1d), and two from entirely different major-clades (Dx9a + Dy9b and Dx13b + Dy13a). Also, within subclade 9 were the SDS-PAGE mobilities Dx2 + Dy10 and Dx2 + Dy11, and within subclade 13 were the SDS-PAGE mobilities 1t + 12, 2.1* + 12.1*, and 4 + 10 further supporting that these haplotypes are not all similar to the wheat 2 + 12 haplotype at the molecular level. However, the proteins still migrate similarly on an SDS-PAGE. These results suggest that SDS-PAGE alone is insufficient when characterizing HMW glutenin diversity in wild relatives and will not be a suitable tool for tracking unique alleles in the hexaploid wheat germplasm.

The molecular haplotypes for Ae. tauschii accessions at the sites where the accessions were collected. Corresponding SDS-PAGE allele is noted for 2 + 12 mobility alleles.
While most molecular haplotypes delineated along the three Ae. tauschii lineages (Fig. 1), a notable exception was within the predominantly Lineage 1 major-clade, subclade 9, where the same three haplotypes (Dx9a + Dy9a, Dx9a + Dy9b, and Dx9a + Dy9c) were observed in both Lineage 1 and Lineage 2 accessions. Interestingly, while there were three haplotypes at the y subunit, there was only a single x haplotype associated with all three of these. The x subunit mobility was the same for all three haplotypes, indicating that the x allele is in fact the same. However, the y subunit was differentiated with the mobility Dy9b was faster than that of Dy9a and Dy10c (Supplemental Fig. 1).
Recombinant haplotypes identified
The close proximity of the glutenin genes results in such tight linkage that recombination is extremely rare. To date, a recombination between the x and y subunit of any HMW-GS locus has yet to be verified. Among the 242 Ae. tauschii accessions studied here, we found a clear example of a historical recombination at Glu-D1 in the Lineage 2 accession TA1668 (BW_01111). SDS-PAGE mobility of TA1668 matches that of TA10081 (BW_01039), Dx2 + Dy10.2. TA1668 and TA10081 share the same y subunit haplotype, Dy5a of major-clade III. However, the x subunit of TA1668 corresponds to haplotype Dx9a of major-clade I (Fig. 4a). The corresponding subclade 9 is the only subclade found in both Lineage 1 and Lineage 2 accessions, indicating that there was incomplete lineage sorting or admixture between the two lineages that lead to the introgression of a Lineage 1 Glu-D1 haplotype into the Lineage 2 population. In the overlapping geographic range of both haplotypes, it appears there was a rare recombination between the Lineage 1 and Lineage 2 Glu-D1 alleles, leading to the recombinant Dx9a + Dy5a found in TA1688 (Fig. 4c).

Molecular haplotype representation of recombinant Glu-D1 haplotypes for accessions a TA1668 (Lineage 2) and b TA2576 (Lineage 3). Vertical purple bars represent the alternate allele variants as called against the AL8 7/8 genome assembly. SDS-PAGE allele for the given haplotypes are shown to the right of each gene model. Haplotype Dx9a is present in both Lineage 1 and Lineage 2 accessions. The closest potential x and y subunit haplotypes involved in the recombinant haplotype x7b + y15b of TA2576 are Dx7a (Lineage 3) and Dy15a (Lineage 2). SDS-PAGE protein mobilities for Dx7a + Dy7a and x7b + y15b were not analyzed. Geographical distribution of recombinant Glu-D1 haplotypes for c TA1668 (BW_01111), Lineage 2, and d TA2576, Lineage 3. Collection site of recombinant accessions is marked in lime green, whereas turquoise and orange designate the collection sites of accessions carrying x subunit and y subunit haplotypes, respectively. Accessions with unrelated haplotypes are in light gray. Lineages are shown in squares (Lineage 1), circles (Lineage 2), or triangles (Lineage 3).
The Lineage 3 accession TA2576 also appears to carry a recombinant haplotype (Dx7b + Dy15b) (Fig. 4b). However, our data set did not contain the exact haplotypes involved in the recombination that led to Dx7b + Dy15b. The closest x subunit haplotype is Dx7a, the only other Lineage 3 haplotype, from major-clade III and the closest y subunit is the Lineage 2 haplotype Dy15a from major-clade II (Lineage 2). We, therefore, designated the x and y subunit haplotypes of TA2576 haplotypes within subclades 7 and 15. Geographical analysis reveals that TA2576 was collected from a region shared with other Lineage 3 accessions (Fig. 4d). However, the accessions containing Dy15a haplotype were not collected from a shared region with the Lineage 3 accessions. Although not conclusive, the most parsimonious explanation is therefore that Dx7b + Dy15b represents a recombinant haplotype between the x and y subunits from two different alleles. Within our current panel, however, we are unable to differentiate exactly which original haplotypes gave rise to this recombinant haplotype.
Functional variation in Glu-D1
To see if any of the variants with the coding region were potentially functional, we compared deduced amino acid sequences based on the reference genome and single nucleotide variant information. We were specifically looking for changes in cysteine residues and other non-synonymous mutations leading to the amino acid changes, but we did not detect any changes in cysteine residue composition in the N or C terminal regions (Supplemental Data 3). However, our results do show that a majority of variant sites resulted in non-synonymous changes in amino acid compositions (e.g., 39 out of 68 in Glu-D1y) supporting a hypothesis that these storage proteins are not under a purifying selection and any of these mutations could have a potentially substantial impact on quality (Supplemental Data 3). Nonetheless, the superior 5 + 10 allele again clustered very tightly with Lineage 3 accessions based on the protein sequence (Supplemental Figs. 2 and 3).
The Glu-D1 locus of wheat provides the greatest contribution to gluten strength, regardless of the allele present13. The allelic diversity of Glu-D1 in wheat is limited to two predominant alleles 2 + 12 and 5 + 10, and a few rare alleles (i.e., 3 + 12, 4 + 10) which are associated with similar end-use quality as 2 + 1225,26. The unique 2.2 + 12 SDS-PAGE allele, which is found at high frequency in Japanese wheat, was shown to be identical to the 2 + 12 haplotype with the exception of additional repeats in the internal repeat domain of the x subunit25,27,28. The x subunit protein from 5 + 10 has a unique cysteine residue just within the central repeat domain which is suspected to increase disulfide bonds in the forming dough. The early expression and transcription of this allele are also higher than that of the other Glu-D1 alleles, in particular 2 + 1216. Other factors underlying the functionality of high-molecular weight glutenin alleles in quality have also been proposed, such as the length and composition of the central repeat domain29,30, the position and number of cysteine residues for disulfide bonds31, the glutamine composition for hydrogen bonds31,32, and promoter elements33,34. At this time, it is unclear which of these characteristics, or the combination and interaction of different features, lend to superior quality characteristics and laboratory testing of end-use quality is necessary to determine superior and inferior alleles. As the first step needed to identify superior quality alleles, we developed Ae. tauschii Glu-D1 haplotype groups from dense molecular markers. This haplotype information will help breeders target novel alleles not currently found in the hexaploid wheat germplasm for advancement to quality testing stages. While much work remains to be done on the functional variants within Glu-D1 and other glutenin genes, the determination of novel variants and unique haplotypes gives priority for the extensive germplasm development and testing. In the absence of functional variant knowledge, the assessment of molecular diversity in this study, however, supports the approach of identifying novel haplotypes followed by targeted testing in the breeding pipelines as the best path forward for wheat improvement. The unique origin of 5 + 10 from Lineage 3, however, further supports the important contributions of genetic diversity from Ae. tauschii to the wheat D genome consistent with the findings by Gaurav et al.6.
Conclusions
Our haplotype analysis revealed that the x and y subunits are strongly associated even in diverse germplasm and that the Glu-D1 haplotypes were clustered to specific geographic origins. Consistent with the findings of Gaurav et al.6, we found evidence of two lineages (Lineage 2 and Lineage 3) contributing to the D genome of wheat with the superior 5 + 10 allele found associated with Lineage 3 accessions. Given the excellent end-use quality imparted by 5 + 10, understanding this unique origin of the wheat allele support the further exploration and evaluation of novel Glu-D1 alleles to further improve wheat quality. This also supports the potential of unique alleles and haplotypes from the breadth of Ae. tauschii diversity.
Wheat grain quality remains one of the most important targets for breeders to develop superior wheat cultivars. Wild wheat relatives have been shown as a valuable resource for accessing novel genetic diversity to improve a range of wheat breeding targets including yield and disease resistance. For quality evaluation, however, the large quantity of grain needed for milling and baking and the confounding morphological characteristic needed for quality evaluation, such as suitable seed size for milling, make a direct evaluation of various end-use quality traits intractable to phenotype directly these wild relatives, including Ae. tauschii. In this work, we, therefore, took the first step in a reverse genetics approach in Ae. tauschii by identifying and characterizing variants at the important Glu-D1 locus. This demonstrated the unique origin of the Glu-D1 allele in wheat as well as uncovering novel allele variants and haplotypes that can now be targeted for breeding. We also established the relation of wheat alleles to those of Ae. tauschii and have shown that Ae. tauschii contains a trove of unique Glu-D1 alleles very unlike the alleles in the current wheat germplasm. With accessible germplasm resources such as synthetic hexaploids6, the diagnostic variants will enable marked-assisted selection of novel Ae. tauschii introgressions into wheat, characterization of their end-use quality, and utilization in wheat improvement.
Methods
Plant material
This study included 273 Aegilops tauschii accessions, of which 241 were from the Wheat Genetics Resource Center (WGRC) collection at Kansas State University in Manhattan, KS, USA. Another 28 were from the National Institute for Agricultural Botany (NIAB) in Cambridge, United Kingdom. An additional 2 were from the Commonwealth Scientific and Industrial Research Organisation (CSIRO) in Canberra, Australia. The final accession, AL8/78, was obtained from the John Innes Center (JIC) in Norwich, Norfolk, England. Data regarding original collection sites for the WGRC accessions are detailed in Supplementary Data 15. Aegilops tauschii is divided into two subspecies, spp. tauschii (Lineage 1) and the wheat D-genome donor spp. strangulata (Lineage 2). In this data set, 117 accessions were Lineage 1 and 143 Lineage 2. An additional eight accessions (five non-redundant) belonged to the newly described Lineage 36.
SDS-PAGE analysis
The sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis of 72 of the Ae. tauschii accessions was conducted at the Wheat Chemistry and Quality Laboratory at the International Maize and Wheat Improvement Center (CIMMYT) Texcoco, Mexico according to Singh et al.35 with the following modifications. Specifically, 20 mg of wholemeal flour were mixed at 1400 rpm with 0.75 ml of 50% propanol (v/v) for 30 min at 65 °C in a Thermomixer Comfort (Eppendorf). The tubes were then centrifuged for 2 min at 10,000 rpm, and the supernatant containing the gliadins was discarded. The pellet was then mixed with 0.1 ml of a 1.5% (w/v) DTT solution in a Thermomixer for 30 min at 65 °C, 1400 rpm, and centrifuged for 2 min at 10,000 rpm. A 0.1 ml volume of a 1.4% (v/v) vinylpyridine solution was then added to the tube which was subsequently placed again in a Thermomixer for 15 min at 65 °C, 1400 rpm, and centrifuged for 5 min at 13,000 rpm. The supernatant was mixed with the same volume of sample buffer (2% SDS (w/v), 40% glycerol (w/v), and 0.02% (w/v) bromophenol blue, pH 6.8) and incubated in the Thermomixer for 5 min at 90 °C and 1400 rpm. Tubes were centrifuged for 5 min at 10,000 rpm, and 8 ml of the supernatant were used for the glutenin gel. Glutenins were separated in polyacrylamide gels (15% or 13% T) prepared using 1 M Tris buffer, pH of 8.5. Gels were run at 12.5 mA for ~19 h. Alleles were identified using the nomenclatures proposed by Payne and Lawrence19 for bread wheat high-molecular weight glutenins and Lagudah and Halloran24 for previously described Ae. tauschii high-molecular weight glutenins.
DNA sequencing
Whole-genome Illumina paired-end sequencing to 10× coverage for most accessions, and 30× coverage for select accessions, was obtained from TruSeq PCR-free libraries with 350 bp insert with Illumina paired-end sequencing of 150 bp according to manufactures recommendations. Sequence datasets are detailed in Gaurav et al.6 and publicly available through NCBI sequence repositories PRJNA68512536 and PRJNA69498037.
Variant calling and duplicated accessions
Paired-end reads of the 306 Ae. tauschii samples were aligned to the Ae. tauschii ‘AL8/78’ genome assembly38. Hexaploid wheat samples were aligned to an in silico reference assembly including the hexaploid wheat A and B genomes from ‘Jagger’39 combined with the Aetv4.0 D genome using HISAT2 version 2.1.0 with default parameters40. Alignments were sorted and indexed using samtools v1.941. Allele calls were made for all sites within the coding and 2.5 kb flanking regions of the x and y subunits of Glu-D1 for the tauschii and wheat samples independently using bcftools version 1.942 ‘mpileup’ and ‘call’ commands with a minimum alignment quality of 20 (—q 20). The two resulting genomic variant call files for the Ae. tauschii and wheat samples were then merged with the ‘merge’ command of bcftools. All sites were called regardless of variant presence to avoid missing fixed variants within the two populations. Duplicated accessions were identified as sharing greater than 99.8% variant calls across their entire genome43.
Molecular haplotype analysis
After merging the Ae. tauschii and wheat genomic variant call files, the genotype calls with mapping quality (QUAL) >40 were made on the ratios of the read depths of alternate allele (DV) to total read depths (DP) with an AWK script first presented in Milner et al., 201944. Monomorphic sites with minor allele frequency less than 1% or missing calls in greater than 90% of samples were filtered out. Molecular haplotypes for Glu-D1 were developed on the 308 resulting variant sites present in 273 of the Ae. tauschii accessions (listed in Supplemental Data 1) and 4 of the hexaploid wheat varieties (‘Chinese Spring’, ‘LongReach Lancer’, ‘CDC Stanley’, and ‘CDC Landmark’). Samples sharing the same variants were considered to share the same molecular haplotype.
Genetic distances were calculated as the Euclidean distance on the A matrix of the variants in R. The A matrix was calculated with ‘A.mat()’ from the rrBLUP package45 and Euclidean distances with ‘dist()’. Hierarchical clustering of the genetic distances was found using hclust() and converted to a dendrogram object before plotting with the dendextend package46.
Molecular haplotypes were designated by the subclade number of the x and y subunits together and then by the letter corresponding to the individual gene-level haplotype within. For example, molecular haplotype x1a + y1b represents the ath x haplotype and bth y haplotype within the subclade 1. It should be noted that letter designations across subclades have no correspondence. The ath x haplotype of subclade 1 is different than that of subclade 2. We later used the variant sites to generate a maximum likelihood tree using iqtree2 (https://github.com/iqtree/iqtree2) and visualized the unrooted version of it using iTOL47 (Online interactive access: https://itol.embl.de/tree/70179110196411141623118598).
Variant annotations and protein-based phylogeny
Variants were annotated using SNPeff software48 using standard codon tables and the AetV4 reference genome. Variant file (vcf) were further converted to nucleotide sequences using BCFtools (consensus) and translated into proteins using EMBOSS_transeq (https://www.ebi.ac.uk/Tools/st/emboss_transeq/). Protein sequences were used for phylogenetic tree construction using iqtree2 with ModelFinder49 and 1000 bootstraps. The tree was visualized and annotated using iTOL47.
Colocalization of 5 + 10 allele with Lineage 3 k-mers on 10+ bread wheat genomes
K-mers were produced using Jellyfish (https://github.com/gmarcais/Jellyfish) and lineage-specifick-mer bins were determined as described in Gaurav et al.6,50. Briefly, if within a 100 kb bin, Lineage 3-specifick-mers counts are greater than either Lineage 1 and Lineage 2 counts by a margin (0.01% of 100k), the bin will be considered Lineage 3-specific bins. Colocalization of lineage-specific regions and 5 + 10 alleles were plotted using ggplot2 based on lineage-specifick-mer bin distribution datasets and Glu-D1 positions on the 10+ Wheat Genomes39.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Sequencing data can be accessed through OWWC main paper (journal submission ID: NBT-PI52743) with the corresponding NCBI sequence repositories PRJNA685125 and PRJNA694980. Variant call file of the entire genome of all Ae. tauschii accessions can be accessed from Zenodo under https://doi.org/10.5281/zenodo.4317950. The variant calls for the Glu-D1 locus are provided as Supplementary Data Set 2 with this manuscript. The population information for the Aegilops tauschii accessions is provided as Supplementary Data Set 1. Ae. tauschii lineage-specifick-mer counts in wheat genome assemblies can be accessed from Zenodo under https://doi.org/10.5281/zenodo.4474428.
Code availability
Code for sequence alignments and variant calling is available at https://github.com/wheatgenetics/owwc/tree/master/variant_call_and_redundancy. R code used to generate figures is available at https://github.com/emilydelorean/Glu-D1_Aegilops-tauschii. Code for SNP effects (SNPeff) and iqtree and k-mer co-distributions and related analysis can be found here: https://github.com/umngao/GluD1_commsBio2021.
References
Salamini, F., Özkan, H., Brandolini, A., Schäfer-Pregl, R. & Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 3, 429–441 (2002).
Dubcovsky, J. & Dvorak, J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 316, 1862–1866 (2007).
Wang, J. et al. Aegilops tauschii single nucleotide polymorphisms shed light on the origins of wheat D-genome genetic diversity and pinpoint the geographic origin of hexaploid wheat. N. Phytol. 198, 925–937 (2013).
Zhou, Y. et al. Triticum population sequencing provides insights into wheat adaptation. Nat. Genet. 52, 1412–1422 (2020).
Singh, N. et al. Genomic analysis confirms population structure and identifies inter-lineage hybrids in Aegilops tauschii. Front. Plant Sci. 10, https://doi.org/10.3389/fpls.2019.00009 (2019).
Gaurav, K. et al. Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement. Nat Biotechnol https://doi.org/10.1038/s41587-021-01058-4 (2021).
Shewry, P. What is gluten—why is it special? Front. Nutr. 6, https://doi.org/10.3389/fnut.2019.00101 (2019).
Lutz, E., Wieser, H. & Koehler, P. Identification of disulfide bonds in wheat gluten proteins by means of mass spectrometry/electron transfer dissociation. J. Agric. Food Chem. 60, 3708–3716 (2012).
Anderson, O., Rausch, C., Moullet, O. & Lagudah, E. The wheat D-genome HMW-glutenin locus: BAC sequencing, gene distribution, and retrotransposon clusters. Funct. Integr. Genomics 3, 56–68 (2003).
Kong, X. Y., Gu, Y. Q., You, F. M., Dubcovsky, J. & Anderson, O. D. Dynamics of the evolution of orthologous and paralogous portions of a complex locus region in two genomes of allopolyploid wheat. Plant Mol. Biol. 54, 55–69 (2004).
Gu, Y. Q. et al. Types and rates of sequence evolution at the high-molecular-weight glutenin locus in hexaploid wheat and its ancestral genomes. Genetics 174, 1493–1504 (2006).
Shewry, P. R., Halford, N. G., Belton, P. S. & Tatham, A. S. The structure and properties of gluten: an elastic protein from wheat grain. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 357, 133–142 (2002).
Wang, Z. et al. New insight into the function of wheat glutenin proteins as investigated with two series of genetic mutants. Sci. Rep. 7, 3428 (2017).
Rooke, L. et al. Overexpression of a gluten protein in transgenic wheat results in greatly increased dough strength. J. Cereal Sci. 30, 115–120 (1999).
Lafiandra, D., D’Ovidio, R., Porceddu, E., Margiotta, B. & Colaprico, G. New data supporting high mr glutenin subunit 5 as the determinant of quality differences among the pairs 5 + 10 vs. 2 + 12. J. Cereal Sci. 18, 197–205 (1993).
Don, C., Lookhart, G., Naeem, H., MacRitchie, F. & Hamer, R. J. Heat stress and genotype affect the glutenin particles of the glutenin macropolymer-gel fraction. J. Cereal Sci. 42, 69–80 (2005).
Zhang, P. P., Ma, H. X., Yao, J. B. & He, Z. H. Effect of allelic variation and expression quantity at glu-1 loci on size distribution of glutenin polymer in common wheat. Acta Agron. Sin. 35, 1606–1612 (2009).
Payne, P. I., Holt, L. M. & Law, C. N. Structural and genetical studies on the high-molecular-weight subunits of wheat glutenin: Part 1: Allelic variation in subunits amongst varieties of wheat (Triticum aestivum). Theor. Appl. Genet. 60, 229–236 (1981).
Payne, P. I. & Lawrence, G. J. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun. 11, 29–35 (1983).
William, M. D. H. M., Peña, R. J. & Mujeeb-Kazi, A. Seed protein and isozyme variations in Triticum tauschii (Aegilops squarrosa). Theor. Appl. Genet. 87, 257–263 (1993).
Xu, S., Khan, K., Klindworth, D. & Nygard, G. Evaluation and characterization of high-molecular weight 1D glutenin subunits from Aegilops tauschii in synthetic hexaploid wheats. J. Cereal Sci. 52, https://doi.org/10.1016/j.jcs.2010.05.004 (2010).
Mackie, A. M., Lagudah, E. S., Sharp, P. J. & Lafiandra, D. Molecular and biochemical characterisation of HMW glutenin subunits from T. tauschii and the D genome of hexaploid wheat. J. Cereal Sci. 23, 213–225 (1996).
Gianibelli, M. C., Gupta, R. B., Lafiandra, D., Margiotta, B. & MacRitchie, F. Polymorphism of high mr glutenin subunits in Triticum tauschii: characterisation by chromatography and electrophoretic methods. J. Cereal Sci. 33, 39–52 (2001).
Lagudah, E. S. & Halloran, G. M. Phylogenetic relationships of Triticum tauschii the D genome donor to hexaploid wheat. Theor. Appl. Genet. 75, 592–598 (1988).
Wan, Y. et al. Comparative analysis of the D genome-encoded high-molecular weight subunits of glutenin. Theor. Appl. Genet. 111, 1183–1190 (2005).
Dong, Z. et al. Haplotype variation of Glu-D1 locus and the origin of glu-D1d allele conferring superior end-use qualities in common wheat. PLoS ONE 8, e74859 (2013).
Payne, P. I., Holt, L. M. & Lawrence, G. J. Detection of a novel high molecular weight subunit of glutenin in some Japanese hexaploid wheats. J. Cereal Sci. 1, 3–8 (1983).
Shimizu, K. K. et al. De novo genome assembly of the japanese wheat cultivar Norin 61 highlights functional variation in flowering time and fusarium resistance genes in East Asian genotypes. Plant Cell Physiol. https://doi.org/10.1093/pcp/pcaa152 (2020).
Pézolet, M., Bonenfant, S., Dousseau, F. & Popineau, Y. Conformation of wheat gluten proteins Comparison between functional and solution states as determined by infrared spectroscopy. FEBS Lett. 299, 247–250 (1992).
D’Ovidio, R., Anderson, O. D., Masci, S., Skerritt, J. & Porceddu, E. Construction of novel wheat high-mrglutenin subunit gene variability: modification of the repetitive domain and expression in E. coli. J. Cereal Sci. 25, 1–8 (1997).
Shewry, P. R., Popineau, Y., Lafiandra, D. & Belton, P. Wheat glutenin subunits and dough elasticity: findings of the EUROWHEAT project. Trends Food Sci. Technol. 11, 433–441 (2000).
Lafiandra, D., Masci, S. & D’Ovidio, R. The Gluten Proteins Vol. 295 (Royal Society of Chemistry, 2004).
Ravel, C. et al. Conserved cis-regulatory modules in promoters of genes encoding wheat high-molecular-weight glutenin subunits. Front. Plant Sci. 5, 621–621, (2014).
Gao, Y. et al. The endosperm-specific transcription factor TaNAC019 regulates glutenin and starch accumulation and its elite allele improves wheat grain quality. Plant Cell 33, 603–622 (2021).
Singh, N. K., Shepherd, K. W. & Cornish, G. B. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J. Cereal Sci. 14, 203–208 (1991).
John Innes Centre. Exploring and Exploiting Diversity in Aegilops tauschii. http://www.ncbi.nlm.nih.gov/bioproject/PRJNA685125 (National Center for Biotechnology Information, 2020).
Kansas State University. Hexaploid Bread Wheat Cultivars or Elite Lines (United States mostly) Plus Two Aegilops tauschii. http://www.ncbi.nlm.nih.gov/bioproject/PRJNA694980 (National Center for Biotechnology Information, 2021).
Johns Hopkins University. Aegilops tauschii subsp. strangulata cultivar:AL8/78 Genome Sequencing and Assembly. http://www.ncbi.nlm.nih.gov/bioproject/PRJNA341983 (National Center for Biotechnology Information, 2017).
Walkowiak, S. et al. Multiple wheat genomes reveal global variation in modern breeding. Nature 588, 277–283 (2020).
Kim, D., Paggi, J. M., Park, C., Bennett, C. & Salzberg, S. L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 37, 907–915 (2019).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 27, 2987–2993 (2011).
Gao, L. & Open Wild Wheat Consortium. Open Wild Wheat Consortium Variant (SNP) dataset. Zenodo https://doi.org/10.5281/zenodo.4317950 (2020).
Milner, S. G. et al. Genebank genomics highlights the diversity of a global barley collection. Nat. Genet. 51, 319–326 (2019).
Endelman, J. B. Ridge regression and other kernels for genomic selection with R package rrBLUP. Plant Genome 4, https://doi.org/10.3835/plantgenome2011.08.0024 (2011).
Galili, T. dendextend: an R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 31, 3718–3720 (2015).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. https://doi.org/10.1093/nar/gkab301 (2021).
Cingolani, P. et al. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 6, 80–92 (2012).
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A. & Jermiin, L. S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589 (2017).
Kumar, G. & Open Wild Wheat Consortium. Aegilops tauschii lineage admixture in wheat D subgenome data set. Zenodo https://doi.org/10.5281/zenodo.4474428 (2021).
Acknowledgements
E.D. was supported through Monsanto’s Beachell-Borlaug International Scholars Program. This material is based upon work supported by the National Science Foundation under Award No. 1822162 “Phase II IUCRC at Kansas State University Center for Wheat Genetic Resources WGRC” and Award No. 1339389 “GPF-PG: Genome Structure and Diversity of Wheat and Its Wild Relatives”. B.B.H.W. was supported by the UK Biotechnology and Biological Sciences Research Council Designing Future Wheat Institute Strategic Programme BB/P016855/1. Any opinions, findings, and conclusions, or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Author information
Authors and Affiliations
Consortia
Contributions
E.D. analyzed data, interpreted results, designated haplotypes, composed figures, and wrote the manuscript. L.G. contributed to sequence alignment, variant calling and annotation, and explored GATK method of variant calling to identify potential cysteine changes, Fig. 1 generation, and manuscript editing. M.I.B. contributed to SDS-PAGE analysis and manuscript editing. J.F.C.L. contributed to SDS-PAGE analysis. B.B.H.W. contributed to experimental conceptualization and design, and manuscript editing. OWWC provided Ae. tauschii accessions, geographic sample information and DNA sequence data. J.P. conceptualized experiments, experimental design and analysis, wrote and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Communications Biology thanks Kay Trafford and the other, anonymous, reviewers for their contribution to the peer review of this work. Primary Handling Editor: Luke R. Grinham. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Delorean, E., Gao, L., Lopez, J.F.C. et al. High molecular weight glutenin gene diversity in Aegilops tauschii demonstrates unique origin of superior wheat quality. Commun Biol 4, 1242 (2021). https://doi.org/10.1038/s42003-021-02563-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-021-02563-7
This article is cited by
-
Origin and evolution of the bread wheat D genome
Nature (2024)
-
Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement
Nature Biotechnology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
