
Abstract
Panicle architecture directly affects crop productivity and is a key target of high-yield rice breeding. Panicle length strongly affects panicle architecture, but the underlying regulatory mechanisms are largely unknown. Here, we show that two quantitative trait loci (QTLs), PANICLE RACHIS LENGTH5 (Prl5) and PRIMARY BRANCH LENGTH6 (Pbl6), independently regulate panicle length in rice. Prl5 encodes a gibberellin biosynthesis enzyme, OsGA20ox4. The expression of Prl5 was higher in young panicles resulting in panicle rachis elongation. Pbl6 is identical to ABERRANT PANICLE ORGANIZATION 1 (APO1), encoding an F-box-containing protein. We found a novel function that higher expression of Pbl6 is responsible for primary branch elongation. RNA-seq analysis revealed that these two genes independently regulate panicle length at the level of gene expression. QTL pyramiding of both genes increased panicle length and productivity. By combining these two genes in various combinations, we designed numerous panicle architecture without trade-off relationship.
Similar content being viewed by others


Introduction
Rice panicle architecture has long been a key target of high-yield rice breeding. Panicle architecture involves several organs, including the panicle rachis, primary branches, and secondary branches (Fig. 1a)1. Primary branches develop from the panicle rachis, and each primary branch gives rise to secondary branches. Panicle branching patterns that are mainly regulated by the number of primary and secondary branches directly determine total grain number. However, improving grain productivity by increasing panicle branching alone is limited, as panicle length and branch length remain constant. Thus, both panicle length and the length of each branch are also important factors determining grain productivity. Natural rice varieties show various panicle phenotypes due to gradual changes in these organs (Fig. 1b). Some varieties have increased numbers of primary branches, and others have increased numbers of secondary branches on each primary branch. Some varieties have longer panicle rachises and others have longer primary branches. Some varieties tend to keep primary branch length longer from lower to upper axis. If the panicle rachis length is constant, the number of primary branches that develop from the panicle rachis is limited. Primary branch length also limits the promotion of secondary branch initiation. Many high-yielding rice cultivars tend to have longer primary branches and produce more secondary branches than standard varieties. Therefore, panicle rachis length and primary branch length influence total grain number and rice productivity as well as branching number. Panicle length strongly affects panicle architecture. Panicle length involves two components: panicle rachis length and the length of the top primary branch. For instance, if only panicle rachis length increases, the plants have a panicle architecture like that shown in Fig. 1d compared to Fig. 1c. If only primary branch length increases, the plants have a panicle architecture like that shown in Fig. 1e. If both panicle rachis and primary branch length increase, the plants have a panicle architecture like that shown in Fig. 1f. Therefore, even if the sizes of only two organs change, the plants show various panicle architectures. Thus, panicle length is a key factor determining the diversity of panicle architecture in rice.

a Schematic diagram of a rice inflorescence. b Panicle phenotypes of five natural rice varieties. Green bars indicate the primary branch tips. Blue bars indicate panicle rachises. Red arrowheads indicate primary branches. Scale bar: 5 cm. c–f Diagram of four types of panicle architecture with different panicle lengths. Panel d has a longer panicle rachis than c. Panel e has longer primary branches than c. Panel f has a longer panicle rachis and primary branches than c. g–i Comparison of panicle traits. g Panicle rachis length. h Primary branch length. i Panicle length. Error bars represent means ± SD (n = 10 plants). *Significant at the 5% level (Tukey’s significant difference test).
Several genes related to panicle morphogenesis have been cloned, some of which were already shown to be agronomically important. Grain Number 1a (GN1a)2, WEALTHY FARMER’S PANICLE (WFP)3, ABERRANT PANICLE ORGANIZATION1 (APO1)4,5, and FRIZZY PANICLE (FZP)6,7,8 significantly increase branch number and total grain number. However, few genes related to panicle length have thus far been cloned. A natural allele of DENSE AND ERECT PANICLE1 (DEP1) regulates panicle length9. Specifically, DEP1 improves yield potential by pleiotropically reducing panicle internode length while increasing grain number. Identifying novel genes related to panicle length would help reveal the fundamental mechanisms that determine various types of variety-specific panicle architecture. Furthermore, these analyses further promote the rational design of ideal panicle architecture for producing elite crop varieties with high yield10,11.
In this study, we performed QTL analysis and identify two QTLs for panicle length from ST-1 (from the Stocked rice collections of Togo field and Nagoya University-1). ST-1 possesses longer panicles than Japanese rice cultivar, Koshihikari. The QTLs were mapped to PANICLE RACHIS LENGTH5 (Prl5) and PRIMARY BRANCH LENGTH6 (Pbl6). Pbl6 contains APO1, a gene previously reported to regulate panicle development. Prl5 encodes Gibberellin 20 oxidase-4. By combining Pbl6 and Prl5, we produced NILs with longer panicles and increased productivity.
Results
Two QTLs regulate panicle length
Natural rice varieties show wide phenotypic variation in panicle length. We selected five natural rice plants characterized by panicle length from the Stocked rice collections of the Togo field and Nagoya University (Fig. 1b). As panicle length is determined by primary branch length and panicle rachis length (Fig. 1a), we measured each organ (Fig. 1g–i). Koshihikari has short panicle like Fig. 1c. Habataki has short panicle rachis but quite long primary branches like Fig. 1e. ST-6 and ST-5 show middle phenotypes between Fig. 1d and f. ST-1 have both longer panicle rachis and primary branches like Fig. 1f. There is a distinct difference between Koshihikari and ST-1 in panicle length and its components (Fig. 1g–i). The main panicles of Koshihikari and ST-1, which has short culms but extremely long panicles (Fig. 2a, b), are ~20 and 35 cm long, respectively (Fig. 1i). Thus, to investigate genes that regulate panicle length, we selected these two rice lines. We produced an F2 population derived from a cross between Koshihikari and ST-1 and measured each organ (panicle length, panicle rachis length, and primary branch length) in 96 of these plants. QTL analysis identified two major QTLs for panicle length (with a log10 odds [LOD] score >3.0) on chromosomes 5 and 6 (Fig. 2c). Interestingly, we also detected the QTL related to panicle rachis length on chromosomes 5 and the QTL related to primary branch length on chromosomes 6; these QTLs were detected in the same position. The QTL on chromosomes 5, qPANICLE RACHIS LENGTH5 (qPrl5), regulates panicle rachis length. The QTL on chromosomes 6, qPRIMARY BRANCH LENGTH6 (qPbl6), regulates primary branch length. These results indicate that panicle length is regulated independently by qPrl5 and qPbl6.

a Gross morphologies of Koshihikari and ST-1. Scale bar: 20 cm. b Panicle morphologies of Koshihikari and ST-1. Scale bar: 20 cm. c QTL locations for panicle length, panicle rachis length, and primary branch length. Red bars indicate results for panicle length. Blue bars indicate results for panicle rachis length. Green bars indicate results for primary branch length, respectively. QTLs for panicle length and panicle rachis length were detected at the same position on chromosome 5. QTLs for panicle length and primary branch length were detected at the same position on chromosome 6.
Isolation and characterization of Pbl6
To identify the causal gene for qPbl6, we narrowed down the candidate region to a 14.7 kb between markers qPBL6_4 and qPBL6_6 using an F3 population derived from the cross between Koshihikari and ST-1. We identified only one gene, Os06g0665400, which was annotated as APO1 encoding an F-box-containing protein (Fig. 3a). The APO1 allele of indica rice variety, Habataki, increases the diameter of internodes and spikelet number4. The sequence of APO1 in ST-1 completely corresponds with that of the dominant Habataki allele (Fig. 3b). We compared the expression levels of APO1 in inflorescence meristems. APO1 was expressed at higher levels in ST-1 compared to Koshihikari during the primary branch initiation stage (Fig. 3c). In addition, we examined primary branch length using Strong Culm2 (SCM2), a near-isogenic line described in Ookawa et al.4, by introgressing the Habataki allele into Koshihikari. SCM2 had longer primary branches than Koshihikari (Fig. 3d). These results indicate that Pbl6 encodes APO1 and that this gene regulates primary branch elongation.

a The qPbl6 locus was detected between RM3827 and RM15753 on Chromosome 6. Numbers on the map indicate the number of recombinants. Positional cloning narrowed the qPbl6 locus to a 14.7-kb region between qPBL6-4 and qPBL6-6 using 2475 plants. Only one gene was predicted to be located in this region by RAP-DB. b Sequence differences between Koshihikari, ST-1, Habataki, and ST-6 around the APO1 region. c Relative expression of Pbl6 (APO1) during the primary branch initiation stage by quantitative RT-PCR. Relative expression levels were calibrated based on Ubiquitin expression. d Comparison of primary branch length between Koshihikari, NIL-Pbl6ST-6 and SCM2. n = 3 in c. n = 30 plants in d. Error bars represent means ± SD. *Significant at the 5% level (Tukey’s significant difference test).
Isolation and characterization of Prl5
We then attempted to isolate the causal gene for qPrl5 by positional cloning. Analysis using F3 plants narrowed the candidate region to a 28.8 kb area between RM18713 and RM18717 (Fig. 4a, b). The Rice Annotation Project Database (RAP-DB) contains two predicted genes, Os05g0421900 and Os05g0422200, in this region. Os05g0422200 encodes a conserved hypothetical protein, and Os05g0421900 encodes Oryza sativa gibberellin 20 oxidase-4 (OsGA20ox4). This gene is homologous to OsGA20ox2, also known as SEMIDWARF1 (Sd1); its null allele produces a semi-dwarf phenotype and was selected during the Green Revolution12,13. Despite being a homologous gene of a famous green revolution gene, the function of OsGA20ox4 has remained unclear. Because the effect of gibberellins (GAs) on cell proliferation is closely involved in the elongation of various plant organs, we focused on OsGA20ox4.

a The qPrl5 locus was detected between RM1237 and Q1568 on chromosome 5. b Numbers on the map indicate the number of recombinants. Positional cloning narrowed the qPrl5 locus to a 28.8-kb region between RM18713 and RM18717 using 1900 plants. Two genes were predicted to be located in this region by RAP-DB. c Sequence differences between Koshihikari and ST-1 around the OsGA20ox4 region. d, e Expression analysis of Prl5 (GA20ox4) (d) and Sd1 (GA20ox2) (e) in roots, first internodes, and young panicles. Relative expression levels were calibrated based on Ubiquitin expression. Error bars represent means ± SD (n = 3). f, g Comparison of culm length (f) and panicle rachis length (g) between plants transformed with an empty vector (vector control) and expression vector for OsGA20ox4ST-1 under the control of the Actin promoter. Error bars represent means ± SD (n = 4 plants). *Significant at the 5% level (Student’s t-test).
First, we compared the sequence of the 12 kb candidate region between the two varieties and identified several deletions and insertions in both the promoter and coding regions (Fig. 4c). GA20ox enzymes catalyze several reactions in the GA biosynthetic pathway to produce precursors that are further converted into bioactive GAs by subsequent enzymes in the pathway14. To investigate whether the single nucleotide variants in the coding region of this gene affect enzymatic activity, we compared the enzymatic activities of recombinant GA20OX4Koshihikari and GA20OX4ST-1 proteins in the GA biosynthesis pathways. Both proteins showed similar levels of enzymatic activity in the GA biosynthesis pathways (Supplementary Fig. 1). This result indicates that the single nucleotide variants in the GA20ox4ST-1 coding region are not responsible for the high GA biosynthetic activity in ST-1. We further investigated the expression patterns of OsGA20ox4 to confirm the notion that the base substitution in the GA20ox4ST-1 promoter region is responsible for the longer panicle rachis phenotype of ST-1. Quantitative RT-PCR analysis of OsGA20ox4 in young panicles revealed that this gene was expressed at higher levels in ST-1 than in Koshihikari (Fig. 4d). OsGA20ox4 was expressed at low levels in roots and internodes and at high levels in young panicles. These results suggest that the increased expression of OsGA20ox4 in young panicles promotes panicle rachis elongation in ST-1. To confirm that OsGA20ox4 is the gene underlying the Prl5 QTL, we cloned a 1.3 kb cDNA fragment of OsGA20ox4, including the coding region of OsGA20ox4ST-1 (pAct::OsGA20ox4ST-1), and transformed this genomic fragment into Nipponbare. OsGA20ox4 was expressed at high levels in the transgenic lines (Supplementary Fig. 2). The transgenic lines had a significantly higher culm length than the vector control (Fig. 4f). This result indicates that GA20ox4ST-1 also has enzymatic activity in vivo. The transgenic lines also had longer panicle rachises than the vector control (Fig. 4g), suggesting that Prl5 encodes OsGA20ox4, which regulates panicle rachis elongation. The increased expression of Prl5 presumably leads to higher GA accumulation in inflorescence meristems, resulting in panicle rachis elongation.
Expression analysis of Prl5 and Pbl6
To clarify the different functions of the homologous genes, Sd1 and Prl5, we compared their expression patterns. Sd1 was strongly expressed in internodes, with lower expression levels in ST-1 than in Koshihikari (Fig. 4e). Prl5 was highly expressed in young panicles, with higher expression levels in ST-1 (Fig. 4d). Although Sd1 and Prl5 are homologous genes, these organ-specific expression patterns contribute to their different functions. Furthermore, in ST-1, the expression level of Sd1 was lower but expression level of Prl5 was higher compared to Koshihikari. These results correspond with the phenotypes of ST-1, with a shorter culm length but longer panicles compared to Koshihikari (Fig. 2a, b).
Because differences in panicle length may appear during the early stages of panicle development, we investigated Prl5 and Pbl6 expression levels during several stages of panicle development. The expression of Prl5 was higher in ST-1 vs. Koshihikari during the secondary branch initiation stage and the 3 mm stage (Fig. 5a). The expression level of Prl5 was highest during the later stage of secondary branch initiation; this trend was observed in both varieties. By contrast, the expression level of Pbl6 was higher in ST-1 than in Koshihikari during the primary branch initiation stage (Fig. 5b). The expression level of Pbl6 was higher during the primary branch initiation stage compared to the other stages; this trend was observed in both varieties. Pbl6 expression was activated first, followed by Prl5 expression. There were major differences in the timing of expression between Prl5 and Pbl6. Pbl6 was expressed at the earlier stages of panicle development. A high expression level of Pbl6 increases the size of the inflorescence meristem5, resulting an elongated primary branch length. By contrast, Prl5 were expressed at later stages.

a, b Expression analysis of Prl5 (a) and Pbl6 (b) in inflorescence tissue at various developmental stages. PB primary branch initiation stage, SBE secondary branch initiation earlier stage, SBL secondary branch initiation later stage, YP3.0: 3 mm stage of young panicles. Relative expression levels were calibrated based on Ubiquitin expression (a, b). Error bars represent means ± SD (n = 3). *Significant at the 5% level (Student’s t-test). c–j In situ hybridization of Prl5 (c–f) and Pbl6 (g–j) during panicle development in Koshihikari. Panels c and g are at the stage of primary branch differentiation. Panels d and h are at the earlier stage of secondary branch differentiation. Panels e and i are at the later stage of secondary branch differentiation. Panels f and j are close-up views of vascular bundles at the later stage of secondary branch differentiation. Scale bars: 100 μm.
We also examined the temporal and spatial expression patterns of these genes by in situ hybridization analyses of inflorescences at various developmental stages (Fig. 5c–j). Pbl6 expression was detected near the primary branch meristems in Koshihikari (Fig. 5g–j). Prl5 expression was detected near the primary and secondary branch meristems (Fig. 5c–e) and vascular bundles (Fig. 5e, f) in Koshihikari. SoGA20ox1, a homolog of Prl5, is known to be associated with stem elongation in spinach under long-day (LD) conditions15,16. SoGA20ox1 transcripts were also detected in vascular tissues by in situ hybridization17, which is similar to the current results. This suggests that the expression in the vascular bundle is important for regulating panicle rachis elongation.
Effect of Prl5 and Pbl6 on panicle architecture
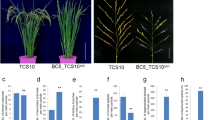
As Prl5 and Pbl6 leads to increased panicle length, which could greatly alter panicle architecture, we evaluated the effects of the Prl5 and Pbl6 loci from ST-1 on panicle architecture. Based on the mapping results, we developed NILs from BC5F2 populations that contained the Prl5 and Pbl6 region from ST-1 in the Koshihikari background (Fig. 6a). The NILs did not differ from one another with respect to plant morphology (Fig. 6b), but their panicle morphology clearly differed (Fig. 6c). We conducted detailed analysis of the panicles of these NILs. The panicle rachises of NIL-Prl5ST-1 were longer than those of Koshihikari (Fig. 6d). The primary branches of NIL-Pbl6ST-1 were longer than those of Koshihikari (Fig. 6e). Due to the effects of Prl5 and Pbl6, the panicles of NIL-Prl5ST-1 + Pbl6ST-1 were 3.7 cm longer than those of Koshihikari (Fig. 6f).

a Graphical genotypes of BC5F2 plants derived from a cross between Koshihikari and ST-1. White and blue bars indicate Koshihikari and ST-1 chromosomes, respectively. Red circles on ST-1 indicate the positions of Prl5ST-1 and Pbl6ST-1. b Gross morphologies of the NILs. Scale bar: 10 cm. c Panicle morphologies of the NILs. Scale bar: 5 cm. d–f Comparison of panicle traits. d Panicle rachis length. e Lengths of the three upper primary branches. f Panicle length. Error bars represent means ± SD (n = 30 plants). *Significant at the 5% level (Tukey’s significant difference test).
Panicle length includes panicle rachis length and the length of the top primary branch. Thus, in the current study, we focused only on upper primary branch length. However, rice plants have many primary branches from the lower to upper axis. Therefore, we investigated whether the effect of Pbl6 is limited to upper primary branch elongation or if it affects the elongation of every primary branch. We observed every primary branch length from lower to upper axis. The Local Regression (LOESS) curves, which represent the length of every primary branch in all four lines, indicate that Pbl6 has an effect on upper primary branch elongation (Fig. 7a). Interestingly, this result also suggests that Prl5 has an effect on lower primary branch elongation (Fig. 7b). In fact, there was no difference in the average length of any primary branch length between NIL-Prl5ST-1 and NIL-Pbl6ST-1 (Fig. 7d). The lower primary branches of NIL-Prl5ST-1 and NIL-Prl5ST-1 + Pbl6ST-1 were longer than those of Koshihikari and NIL-Pbl6ST-1 (Fig. 7e). Due to the combination of these two genes, every primary branch from the lower to upper axis was longer in these lines than in Koshihikari (Fig. 7c).

a–c Comparison of every primary branch length. Solid lines show regression curves. Orange line and dots indicate Koshihikari. Yellow line and dots indicate NIL-Pbl6ST-1 (a). Blue line and dots indicate NIL-Prl5ST-1 (b). Green line and dots indicate NIL-Prl5ST-1 + Pbl6ST-1 (c). n = 42 plants. d–f Comparison of panicle traits and culm length. d every primary branch length. e Lower three primary branch length. f Culm length. Error bars represent means ± SD (n = 30 plants in d and e, n = 35 plants in f). *Significant at the 5% level (Tukey’s significant difference test).
Detailed observation of panicle architecture revealed that Prl5 affects panicle rachis and lower primary branch elongation, whereas Pbl6 affects upper primary branch elongation. Prl5 was expressed during the secondary branch initiation stage. Higher GA levels in vascular bundles led to panicle rachis elongation (Fig. 5a, e, f). Prl5 was expressed in the vascular bundles of young panicles, which are located near the lower primary branch meristems, indicating that bioactive GAs promote not only the elongation of the panicle rachis but also the lower primary branches. By combining Prl5 and Pbl6 in various ways, we could design plants with panicle architecture like that shown in Fig. 1c–f. Because the morphogenesis of multiple organs occurs simultaneously in parallel during panicle development, Prl5 and Pbl6 affect multiple panicle organs. However, when considering only panicle length, we confirmed that Prl5 and Pbl6 regulate panicle length independently.
We then evaluated Prl5 expression in the NIL. The expression level of Prl5 was higher in the young panicles of NIL-Prl5ST-1 vs. Koshihikari (Supplementary Fig. 3a). We also examined Prl5 expression during the later stage of secondary branch initiation by in situ hybridization (Supplementary Fig. 4). We confirmed the expression patterns of Prl5 in the primary and secondary branch meristems and vascular bundles of all four lines. The expression patterns of NILs were well corresponded with that of Koshihikari. As expected, the expression level of Prl5 in vascular bundles was stronger in NIL-Prl5ST-1 and NIL-Prl5ST-1 + Pbl6ST-1 than in Koshihikari and NIL-Pbl6ST-1. To confirm the organ-specific expression of Prl5, we compared the expression patterns of Prl5 and Sd1 in NIL-Prl5ST-1 (Supplementary Fig. 3). Prl5 was expressed at higher levels in young panicles than in internodes. By contrast, Sd1 was expressed at higher levels in the internodes of both lines. There was no difference in the expression level of Sd1 between NIL-Prl5ST-1 and Koshihikari. In fact, there was no difference in culm length (Fig. 7f). These results suggest that Prl5 does not affect plant architecture, but it affects only panicle architecture.
Finally, to confirm that the increased expression of Prl5 in young panicles leads to the increased accumulation of bioactive GA, we quantified GA levels in Koshihikari and NIL-Prl5ST-1 (Supplementary Fig. 5). The levels of both GA44 and GA20 were reduced in NIL-Prl5ST-1. In addition, GA53 and GA24 levels appeared to be reduced in NIL-Prl5ST-1, but these differences were not significant. These results suggest that Prl5 contributes to the increased accumulation of bioactive GA species GA1 and GA4 during panicle development (see Supplementary Note for detailed discussion).
Gene expression analysis of the NILs
To confirm the effects of Prl5 and Pbl6 on panicle length at the gene expression level, we performed RNA-seq analysis to compare the expression patterns of these genes in young panicles of the NILs. The results are shown in Fig. 8a, where the y axis represents the changes in gene expression between Koshihikari and NIL-Prl5ST-1 and the x axis represents the changes in gene expression between Koshihikari and NIL-Pbl6ST-1 in terms of log2 values. The correlation coefficient was 0.31, suggesting that Prl5 and Pbl6 have different biological effects on gene expression. Therefore, Prl5 and Pbl6 also regulate panicle length independently at the gene expression level. Gene Ontology (GO) representation analysis indicated that the biological functions of the two genes are quite different (Fig. 8b, Supplementary Data 3). GO terms involved in cell wall organization (GO:0071555) was enriched, implying that Prl5 closely related to cell elongation of vascular bundle in young panicle. Consistent with Pbl6’s role, GO terms associated with meristem initiation (GO:0010014) was enriched. These results also strongly suggest that Prl5 and Pbl6 regulate panicle length independently.

a RNA was extracted from 2-mm stage young panicles of Koshihikari, NIL-Prl5ST-1, NIL-Pbl6ST-1, and NIL-Prl5ST-1 + Pbl6ST-1 and used for RNA sequencing. The y axis shows the log2 ratios of the changes in gene expression between Koshihikari and NIL-Prl5ST-1, whereas the x axis shows the log2 ratios of the changes in gene expression between Koshihikari and NIL-Pbl6ST-1. The solid line represents the regression line. y = 0.32 X −0.07, R = 0.31. Yellow dots indicate genes whose expression levels changed in NIL-Pbl6ST-1 compared with Koshihikari. Blue dots represent genes whose expression levels changed in NIL-Prl5ST-1 compared with Koshihikari. Orange dots represent common genes whose expression levels changed in both NIL-Prl5ST-1 and NIL-Pbl6ST-1. Gray dots represent other genes. b Heatmap show the P-value (P < 0.05 cutoff) significance of GO terms for Prl5 and Pbl6 target genes. The GO terms listed are the characteristic enriched biological process GO terms for each Prl5 (blue) and Pbl6 (orange). The complete GO term lists and their significance levels are given in Supplementary Data 3. c Venn diagram of genes upregulated or downregulated in NIL-Prl5ST-1 (shown in the blue circle), NIL-Pbl6ST-1 (shown in the yellow circle), and NIL-Prl5ST-1 + Pbl6ST-1 (shown in the green circle). d Heatmap of the expression patterns of the 228 common genes. The expression levels of these 228 genes changed in all three NILs compared with Koshihikari. The expression level of each gene compared with expression in Koshihikari is shown on the map. Red and blue indicate higher and lower expression, respectively. A color scale is shown at the bottom.
RNA-seq analysis revealed 5551 differentially expressed genes in NIL-Prl5ST-1, NIL-Pbl6ST-1, and NIL-Prl5ST-1 + Pbl6ST-1 compared to Koshihikari (Fig. 8c, Supplementary Data 1 and 2). Prl5 regulates 1726 genes and Pbl6 regulates 2337 genes independently. It is highly likely that the 228 common genes with differential expression patterns between Koshihikari and other NILs are closely associated with panicle rachis and primary branch elongation. To compare the expression patterns of the 228 common differentially expressed genes, we constructed a heatmap (Fig. 8d). Several genes had the same expression patterns in all three NILs, including BETA-EXPANSIN7 and CELLULOSE SYNTHASE LIKE F2, which have important roles in cell elongation. Furthermore, the expression patterns of these genes in NIL-Prl5ST-1 and NIL-Pbl6ST-1 were quite different, whereas those in NIL-Pbl6ST-1 and NIL-Prl5ST-1 + Pbl6ST-1 were quite similar. Unexpectedly, among the 228 common genes, Pbl6 appeared to function upstream of Prl5 at the gene expression level. Expression analysis of Koshihikari and ST-1 revealed that Pbl6 is expressed first, followed by Prl5 (Fig. 5a, b). Pbl6 has stronger effects on panicle development in NIL-Prl5ST-1 + Pbl6ST-1, as it was expressed at an earlier stage of panicle development. The differences in the timing of the highest expression levels of Prl5 and Pbl6 result in the similar expression patterns of the 228 common genes in NIL-Pbl6ST-1 and NIL-Prl5ST-1 + Pbl6ST-1.
Effects of Prl5 and Pbl6 on yield performance
We further evaluated the effects of Prl5 and Pbl6 on yield performance under filed condition. NIL-Prl5ST-1 showed no obvious effect on panicle branching (Fig. 9a–c). As Prl5 does not have pleiotropic effects, we could alter only panicle length by introducing Prl5 while maintaining variation-specific branching numbers. NIL-Pbl6ST-1 produced more spikelets per panicle than Koshihikari due to an increase in branch number (Fig. 9a–c). Pbl6 had pleiotropic effects not only primary branch length but also on panicle branching. Although both Prl5 and Pbl6 led to increased panicle length, there were distinct differences between the functions of these two genes. The introgression of Prl5 and Pbl6 had no effects on panicle number (Fig. 9d). NIL-Pbl6ST-1 showed higher yields than Koshihikari due to increased grain number per panicle. NIL-Prl5ST-1 + Pbl6ST-1 had the highest yield among lines (Fig. 9e). These results suggest that combining Prl5 with a gene related to panicle branching and total yields, such as Pbl6, represents an efficient method for breeding higher yielding rice varieties.

a–e Comparison of yield traits. a Grain number. b Number of primary branches. c Number of secondary branches. d Panicle number. e Panicle weight per plant. n = 30 plants in a–c; n = 35 plants in d, e. Error bars represent means ± SD. *Significant at the 5% level (Tukey’s significant difference test).
GNP1 (grain number per panicle1) encodes OsGA20ox118. Wu et al. proposed that the upregulation of GNP1/GA20ox1 in the inflorescence meristems increases cytokinin activity via KNOX-mediated feedback regulation and increases GA catabolic activity. As a result, this process increases cytokinin activity, thereby rebalancing cytokinin and GA activity and increasing grain number and grain yield. However, in the current study, the increased expression of Prl5 did not affect grain number (Fig. 9a), perhaps due to the expression levels and temporal expression patterns of other genes. The expression level of GNP1/GA20ox1 was much higher than that of Sd1/GA20ox2 and Prl5/GA20ox4 in both Koshihikari and NIL-Prl5ST-1 (Supplementary Fig. 6a). In addition, the expression patterns of four KNOX genes in panicle meristems, including OSH1, OSH6, OSH15, and OSH71, did not differ between Koshihikari and NIL-Prl5ST-1 (Supplementary Fig. 6b). Consequently, the levels of several endogenous cytokinins and cytokinin biosynthesis intermediates did not differ between Koshihikari and NIL-Prl5ST-1 (Supplementary Fig. 7). These results suggest that moderately strong expression of Pr15/GA20ox4 with no induction of KNOX genes or active cytokinin production affects panicle rachis length but not grain number. An effect on length but not branching is a key feature of Prl5/GA20ox4.
Isolation and characterization of novel Pbl6 allele
Our analysis showed the possibility that panicle architecture greatly alters depending on the expression levels of Prl5 and Pbl6. To confirm genetically whether these two genes can generate tremendous morphological diversity in rice panicle, we explored novel alleles of two genes and investigated the relationship between expression levels and panicle phenotypes. Koshihikari and ST-6, which have longer panicle length next to ST-1, were chosen for parental varieties (Fig. 1i). We produced F2 population derived a cross between Koshihikari and ST-6 and measured panicle phenotypes in these plants. QTL analysis identified one QTL for primary branch length on chromosomes 6. Positional cloning narrowed the candidate region and identified only one gene, Pbl6. The sequence of Pbl6 in ST-6 is different from the Habataki and ST-1 allele (Fig. 3b). The expression level of Pbl6 in ST-6 was higher than Koshihikari but lower than ST-1 (Fig. 3c). Using BC5F2 population generated from the cross between Koshihikari and ST-6, NIL carrying the ST-6 Pbl6 allele was selected. NIL-Pbl6ST-6 had longer primary branches than koshihikari (Fig. 3d). These results indicate that Pbl6 regulates primary branch length also in ST-6. Since there was only a difference in the promoter region between the sequence of Pbl6 in Koshihikari and ST-6, we concluded that the effect on the primary branch length is due to the different expression level. There was no difference in the upper primary branch length between NIL-Pbl6ST-6 and Koshihikari (Supplementary Fig. 8a). However, the Local Regression (LOESS) curves indicate that, consistent with the expression levels (Fig. 3c), the effect of Pbl6ST-6 is moderate compare to that of Pbl6ST-1 (Supplementary Fig. 8b). These results suggest that Pbl6ST-6 is a different allele, which has moderate effects on primary branch elongation. We further evaluated the effect of Pbl6ST-6 on yield performance under field condition. The NIL-Pbl6ST-6 produced more spikelets per panicle than koshihikari due to an increase in branch number (Supplementary Fig. 8c–e). In addition, the effect of Pbl6ST-6 on panicle branching was moderate compare to Pbl6ST-1. These results suggest that Pbl6ST-6 is a different allele of Pbl6 and primary branch length is certainly modulated by the expression levels of Pbl6.
In this study, we found two genes, Prl5 and Pbl6, independently regulate panicle length. We further identified novel Pbl6 allele, Pbl6ST-6. Combining these genes, we designed numerous panicle architecture (Fig. 10). Although further exploring novel alleles will be needed, we concluded that many of various panicle architecture in rice are generated by various combinations of Prl5 and Pbl6 alleles.

Blue lines indicate the effects of Prl5 and green lines indicate the effects of Pbl6.
Discussion
Previous studies of rice panicle architecture have primarily focused on panicle branching, which directly affects grain yield. However, panicle length also has a key role in yield improvement. Increasing only panicle branching leads to plants with high grain density that tend to suffer from panicle diseases, such as false smut19. Therefore, reducing grain density by regulating panicle and branch length is important for reducing yield losses, but the underlying regulatory mechanisms have been largely unknown. In the current study, we identified two QTLs that independently increase panicle length: PANICLE RACHIS LENGTH5 (Prl5), encoding Gibberellin 20 oxidase; and PRIMARY BRANCH LENGTH6 (Pbl6), encoding APO1. Transcriptome analysis revealed that these two genes independently regulate panicle length at the gene expression level. Using various combinations of Prl5 and Pbl6, we designed numerous panicle architecture without any trade-off between panicle length and grain number. Designing ideal panicle architecture that considers the spatial arrangement of branches by regulating panicle length as well as branch number is essential for obtaining substantial yield improvements in future rice breeding programs.
GAs are crucial phytohormones for plant growth and developmental20,21. Several genes encoding GA metabolic enzymes and key factors in GA signal transduction pathways have been isolated from various plants22. Tenreira et al.23 revealed that FveGA20ox4 dictates the flowering-runnering decision in diploid strawberry, suggesting that GAs have roles in altering plant architecture and plant diversity. There are four GA20ox genes in the rice genome, two of which (OsGA20ox1 and OsGA20ox2) are identical to previously reported genes. In addition, we revealed a function for OsGA20ox4.
Sd1 (SEMIDWARF1) encodes OsGA20ox2; its null allele exhibits a semi-dwarf phenotype but causes no obvious defects in other agronomic traits. This gene was widely used to breed lodging-resistant rice plants during the Green Revolution. Although Sd1/GA20ox2 was strongly expressed in internodes, Pr15/GA20ox4 was expressed at low levels in roots and internodes but highly expressed in young panicles (Fig. 4d, e). Although Sd1/GA20ox2 and Pr15/GA20ox4 are homologous genes, these genes were specifically expressed in internodes and young panicles, respectively. These organ-specific expression patterns contribute to the different functions of these genes. Although Sd1/GA20ox2 was expressed at a lower level in ST-1 vs. Koshihikari, Pr15/GA20ox4 was expressed at a higher level in ST-1. These expression patterns closely correspond to the desirable plant morphology of ST-1 for agriculture, including its shorter culms and longer panicles (Fig. 2a, b).
GNP1 (Grain Number Panicle 1) encodes OsGA20ox1, which increase grain number. Although both GNP1/GA20ox1 and Prl5/GA20ox4 expressed in young panicles, our results indicate that the functions of two genes were different (Fig. 9a, Supplementary Figs. 6 and 7). This difference would result from the expression patterns. The expression level of Prl5/GA20ox4 was lower than that of GNP1/GA20ox1, but the moderate high expression of Prl5/GA20ox4 was enough for panicle rachis elongation. The expression pattern of Prl5/GA20ox4, in the vascular bundles at the secondary branch initiation stage, could contribute to regulate panicle rachis length.
APO1 has pleiotropic effects on rice, as it simultaneously enhances culm strength and increases spikelet number4,5. Our findings reveal a novel effect of APO1. Pbl6 increases grain number while elongating primary branches (Figs. 6e and 9a–c). As a result, introducing Pbl6 did not have a negative effect on the density of branches on each primary branch (Supplementary Fig. 9). Thus, both Prl5 and Pbl6 help maintain the proper spatial arrangement of grains. Although both Prl5 and Pbl6 regulate panicle length, there are obvious differences in the functions of these two genes.
DEP1 improves yield potential by reducing panicle internode length while increasing grain number9. As a result, introducing DEP1 into rice increases panicle density. DEP1 has already been incorporated into Chinese rice breeding. However, Japanese breeders have not used DEP1 for rice breeding due to frequent rains, especially during the rice-cropping season. The high humidity increases the growth of fungi, which cause many diseases. Plants with erect, dense panicles are poorly ventilated, facilitating the growth of fungi. Therefore, rice plants with dense panicles are not suitable for growth in countries with humid seasons such as Japan.
Unlike DEP1, Prl5 and Pbl6 are useful for maintaining the appropriate density of panicle branches. By combining Prl5 and Pbl6 in various ways, we designed five patterns of panicle architecture (Fig. 10). The introduction of Prl5ST-1 helped maintain low primary branch density in NIL-Prl5ST-1, as shown in Fig. 10e. The introduction of Pbl6ST-1 helped maintain a low density of secondary rachilla in NIL-Pbl6ST-1, as shown Fig. 10a. The introduction of Pbl6ST-6 moderately helped maintain a low density of secondary rachilla in NIL-Pbl6ST-6, as shown Fig. 10c. Both Prl5ST-1 and Pbl6ST-1 helped maintain an appropriate spatial arrangement of grains in Prl5ST-1 + Pbl6ST-1 by elongating panicle rachis length and primary branch length, as shown in Fig. 10b. The panicle architecture of NIL-Prl5ST-1 + Pbl6ST-1 is suitable for high-humidity cultivation and has high yields due to the additive effects of Pbl6ST-1.
We further identified different Pbl6 allele, Pbl6ST-6. This result suggests that there are various alleles of Pbl6 in natural rice varieties. The differences in Pbl6 expression levels determine primary branch length of each varieties and generate its diversity. It is assumed that diverse panicle rachis length result from various Prl5 alleles. Our findings showed the first example that combinations of two genes generate diverse panicle architecture.
Novel alleles and additional genes that regulate panicle phenotypes must be identified in order to design ideal panicle architecture for each cultivation area. In addition, our findings indicate that combining genes related to panicle branching and panicle length is a useful approach for improving crop productivity. Several genes have already been shown to increase branch number, such as GN1a, WFP, and FZP2,3,6. We believe that combining Prl5 and Pbl6 with these genes represents an excellent strategy for developing higher yielding rice varieties.
In this study, we produced NILs with various panicle architectures. These NILs are useful materials for rice breeding and should facilitate further research for improving yield potential and grain quality. Grain number has been a key breeding target for improving yield potential. However, simultaneously improving grain number and grain quality is a major challenge due to the negative correlation between grain number and quality. Increasing seed productions often leads to smaller, low-quality grains. Indeed, natural variants of the rice G-protein γ subunit genes DEP1 and GS3 have increased grain yields but typically show only mediocre grain quality24. Filling rate directly affects grain quality and is one of the main factors regulating yield potential. Breeders have recently begun to focus on filling rate as well as grain number. Therefore, in order to further improve yield potential, overcoming the problem of poor grain filling is an important issue that should be addressed in the future25. The grain-filling rate varies greatly depending on panicle position, but how the position of each grain and panicle length affect filling rate has not yet been clarified due to the lack of suitable experimental materials. By using NILs produced in the current and future studies combined with known genes such as GN1a and FZP, it should possible to investigate whether panicle length and the location of each grain affects filling rate. Such information would be useful for identifying the ideal panicle architecture for high-quality grain production to steadily improve rice productivity, as well as designing panicle architecture suitable for different environments and cultivation areas. Identifying genes related to the diversity of rice panicle architecture could provide a deeper understanding of panicle morphogenesis and help breeders create new varieties.
Methods
Plant materials and growth conditions
ST-1, ST-5, and ST-6 was selected from the rice collections of the Field Science Center at Nagoya University. The ST-1 × Koshihikari F1 plant was backcrossed five times with Koshihikari to generate near-isogenic lines. NIL-Prl5ST-1, NIL-Pbl6ST-1, and NIL-Prl5ST-1 + Pbl6ST-1 were selected from the BC5F2 population using single-sequence repeat (SSR) markers. The ST-6 × Koshihikari F1 plant was backcrossed five times with Koshihikari to generate near-isogenic lines. NIL-Pbl6ST-6 was selected from the BC5F2 population using SSR markers. All nontransgenic materials were grown under natural conditions in a paddy field at the Field Science Center at Nagoya University, Togo, Aichi, Japan. Seeds were germinated in a seedbed in early May and transplanted to the field in late June. Plants were grown in the paddy field by single transplanting under conventional cultivation conditions. Transgenic plants were grown in isolated greenhouses at 24 °C under a natural light environment.
Observation of panicle architecture
A main panicle was sampled from each plant for observation. For QTL analysis, panicle length, panicle rachis length, and primary branch length were measured. To evaluate panicle architecture in the NILs, panicle length, panicle rachis length, primary branch length, number of primary branches, number of secondary branches, and total grain number were measured.
QTL analysis and fine mapping
QTL analysis was performed with 96 F2 plants obtained from a cross between Koshihikari and ST-1. F2 recombinant plants were genotyped to identify homozygous recombinant plants for qPrl5 and qPbl6, respectively. Selected F2 plants were selfed, and fine mapping was performed with the F3 populations. Also, QTL analysis was performed with 96 F2 plants obtained from a cross between Koshihikari and ST-6. Fine mapping was performed with the BC6F2 populations. Genomic DNA was extracted from leaves using the cetyltrimethylammonium bromide (CTAB) method, and pure DNA samples were used for genotyping using the SSR markers listed in Supplementary Table 1. PCR for genotyping was performed with Taq polymerase. Data analyses were performed using the software package R/QTL (R version 3.3.3; R/qtl package 1.41–6). QTLs were identified using Haley–Knott regression, and the significance threshold was set using 1000 permutations.
Sequence analysis
Sequence analysis of Prl5 was carried out by comparing the sequences of this gene in ST-1 and Koshihikari. Sequence analysis of Pbl6 was carried out by comparing the sequences of this gene in ST-1, Habataki, ST-6, and Koshihikari. The promoter and coding regions of Prl5 and Pbl6 were amplified by PCR with KOD FX Neo (Toyobo) and the primers listed in Supplementary Table 2. PCR products were purified with Wizard® SV Gel and PCR Clean-Up System (Promega) and sequenced with the primers listed in Supplementary Table 2.
RNA extraction and quantitative RT-PCR analysis
Total RNA was extracted from roots, first internodes, and inflorescence meristems as described by Sambrook et al.26 and treated with DNase I. First-strand cDNA was synthesized from 1 μg of total RNA using a QuantiTect Reverse Transcription Kit (Qiagen). Quantitative RT-PCR was performed using a QuantiTect SYBR Green PCR Kit (Qiagen) and a LightCycler System (LightCycler 1.5; Roche Applied Science). The relative standard curve method was used to calculate the relative transcript levels of the target genes. The results were confirmed using three independent biological replicates. For all experiments, the transcript level of each gene was normalized with that of endogenous OsUBC32. The primer sets used for PCR are listed in Supplementary Table 3.
In situ hybridization
Inflorescence meristems in fixed with 4% (w/v) paraformaldehyde and 0.25% glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.2) overnight at 4 °C and embedded in SCEM compound (SECTION-LAB, Japan). The cut surface was covered with an adhesive film (Cryofilm type IIC9, SECTION-LAB, Japan), and frozen sections (8–14 μm) were prepared with a cryostat (CM 1850 Leica Microsystems, Germany) as previously described27. Digoxigenin-labeled anti-sense RNA probes for Prl5 and Pbl6 transcripts were prepared as described by Komatsu et al.8. Hybridization and immunological detection of the digoxigenin-labeled probes were performed as previously described with some modifications28. The primers used to amplify the cDNAs are listed in Supplementary Table 4.
Generation of transgene constructs and plant transformation
To produce transgenic lines harboring Prl5, the coding sequence of Prl5 was amplified from ST-1 cDNA using a forward primer (5′-GCAAGCTTATGCATGCATCTCCTCACCCATTAC-3′) containing a HindIII site and a reverse primer (5′-ATACTAGTCTAGACGGTGGCCGCCTGGGTCG-3′) containing a SpeI site. The fragment was cloned into the HindIII–SpeI sites of pAct::nos-pCAMBIA1380 and introduced into, japonica rice variety Nipponbare by Agrobacterium tumefaciens (EHA105)-mediated transformation as described by Ozawa et al.29. Transformed cells and plants were selected based on hygromycin resistance, and regenerated seedlings were grown to maturity in pots in a greenhouse. Four T0 plants were analyzed. Control plants were generated via transformation with empty vector.
Measuring endogenous cytokinin and GA levels
Two-mm stage young panicles of Koshihikari and NIL-Prl5ST-1 were collected. The endogenous cytokinin and gibberellin levels were measured as described in Kojima et al.30.
GA20-oxidase activity tests
The coding sequence of each GA20ox gene was cloned into the EcoRI and XhoI sites of a modified pGEX-6p-1 (Novagen) vector. E. coli Rosetta (DE3) pLysS (Novagen) cells were used for protein expression. The bacterial culture (5 mL) was grown at 37 °C overnight and inoculated into fresh LB medium (500 mL) containing 50 μg/mL ampicillin. After the bacterial culture reached an OD600 of 0.6–0.7 at 37 °C, 0.1 mM IPTG was added and the culture was incubated at 18 °C for 16–18 h. The cells were collected by centrifugation and sonicated in lysis buffer (10 mM Na-phosphate [pH 7.5], 150 mM NaCl, 1 mM DTT, Protease Inhibitor Cocktail [Roche]). After centrifugation, the supernatant from the resulting lysate was purified using glutathione Sepharose 4B (GE Healthcare). The resulting proteins were used for the enzyme activity tests. After enzymatic cleavage of the GST tag with PreScission Protease (GE Healthcare), the de-tagged proteins were purified on a Superdex 200 16/60 column (GE Healthcare) or further purified by passage through Glutathione Sepharose 4B resin to remove GST tag contaminants. Protein fractions were concentrated with an Amicon Ultra-4 concentrator unit (10 kDa molecular weight cutoff) and used for subsequent enzyme assays. The purified proteins were quickly frozen in liquid N2 and stored at −80 °C until use. In vitro enzyme activity assays were performed as described for rice GA20ox31. GA19 and GA20 were included in the reaction mixture as a substrate for GA20ox, respectively. Deuterated [17,17-2 H2] GA4 were purchased from OlChemIm and used as internal standards for the GA20ox reactions. Full-scan GC-MS analysis was carried out with a JEOL JMS- K9 mass spectrometer and Agilent Technology 6890N gas chromatograph.
RNA sequence (RNA-seq) and data analysis
RNA was extracted from 2-mm stage young panicles of Koshihikari, NIL-Prl5ST-1, NIL-Pbl6ST-1, and NIL-Prl5ST-1 + Pbl6ST-1 and used. RNA-seq was performed as described by Minami et al.32. Statistical analysis was performed by R, using a P-value of 0.05 as minimum. The edgeR package was used to normalize raw data counts and to generate cpm values33. GO-term enrichment analysis was performed using GOseq package in R statistical environment as described by Nozue et al.34, using a P-value of 0.05 as minimum for statistically significant enrichment. All data generated and analyzed during RNA-seq analysis are included in Supplementary Data 1–3.
Statistic and reproducibility
All the experiments were conducted in at least three biological replicates to ensure the reproducibility. Exact sample number was indicated in the figure legends. Mean from at least three independent biological experiments was presented in each figure, in which error bars represented standard deviation. A number n suggested biological replications indicated in the figure legends. Statical analysis was performed by using R (https://www.r-project.org/). Statistical differences were determined by Student’s t-test and one-way ANOVA with multi-comparison Tukey’s HSD post hoc test. A value of P < 0.05 was considered to be statistically significant. Significant differences are indicated in figures using a star. All experiments were repeated at least three time.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The accession code for the raw sequencing data is GSE151043. The normalized data of RNA-seq are provided as Supplementary Data 1. The analyzed data of RNA-seq are provided as Supplementary Data 2 and 3. Newly generated plasmid is available from the corresponding author on reasonable request. The authors declare that all data supporting the findings of this study are available within the article, its Supplementary Information files and from the corresponding author on reasonable request.
Code availability
Data analysis of RNA-seq was performed by R as explained in the Methods section. The code that supports the findings of this study is available upon reasonable request to the corresponding author.
References
Ikeda, M. et al. Analysis of rice panicle traits and detection of QTLs using an image analyzing method. Breed. Sci. 60, 55–64 (2010).
Ashikari, M. et al. Cytokinin oxidase regulates rice grain production. Science 309, 741–745 (2005).
Miura, K. et al. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat. Genet. 42, 545–549 (2010).
Ookawa, T. et al. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 1, 132 (2010).
Ikeda-Kawakatsu, K. et al. Expression level of ABERRANT PANICLE ORGANIZATION1 determines rice inflorescence form through control of cell proliferation in the meristem. Plant Physiol. 150, 736–747 (2009).
Fujishiro, Y. et al. Comprehensive panicle phenotyping reveals that qSrn7/FZP influences higher-order branching. Sci. Rep. 8, 1–9 (2018).
Bai, X. et al. Regulatory role of FZP in the determination of panicle branching and spikelet formation in rice. Sci. Rep. 6, 1–11 (2016).
Komatsu, Mai, Chujo, Atsushi, Nagato, Yasuo, Shimamoto, K. O. & Kyozuka, J. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 130, 3841–3850 (2003).
Huang, X. et al. Natural variation at the DEP1 locus enhances grain yield in rice. Nat. Genet. 41, 494–497 (2009).
Jiao, Y. Designing plants: modeling ideal shapes. Mol. Plant 12, 130–132 (2019).
Wang, B. & Li, J. Understanding the molecular bases of agronomic trait improvement in rice. Plant Cell 31, 1416–1417 (2019).
Sasaki, A. et al. A mutant gibberellin-synthesis gene in rice. Nature 416, 701–702 (2002).
Ashikari, M. et al. Loss-of-function of a rice gibberellin biosynthetic gene, GA20 oxidase (GA20ox-2), led to the rice ‘Green Revolution’. Breed. Sci. 52, 143–150 (2002).
Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 59, 225–251 (2008).
Lee, D. J. & Zeevaart, J. A. D. Differential regulation of RNA levels of gibberellin dioxygenases by photoperiod in spinach. Plant Physiol. 130, 2085–2094 (2002).
Zeevaart, J. A., Gage, D. A. & Talon, M. Gibberellin A1 is required for stem elongation in spinach. Proc. Natl Acad. Sci. USA 90, 7401–5 (1993).
Lee, D. J. & Zeevaart, J. A. D. Regulation of gibberellin 20-oxidase1 expression in spinach by photoperiod. Planta 226, 35–44 (2007).
Wu, Y. et al. The QTL GNP1 encodes GA20ox1, which increases grain number and yield by increasing cytokinin activity in rice panicle meristems. PLoS Genet. 12, 1–21 (2016).
Kong, F. N. et al. Molecular tagging and mapping of the erect panicle gene in rice. Mol. Breed. 19, 297–304 (2007).
Richards, D. E., King, K. E., Ait-ali, T. & Harberd, N. P. How Giberellin regulates plant growth and development: a molecular genetic analysis of gibberellin signaling. Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 67–88 (2001).
Olszewski, N. et al. Gibberellin signaling: biosyntheis, catabolism, and response pathways. Plant Cell 14, S61–S80 (2002).
Hedden, P. & Phillips, A. L. Gibberellin metabolism: new insights revealed by the genes. Trends Plant Sci. 5, 523–530 (2000).
Tenreira, T. et al. A specific gibberellin 20-oxidase dictates the flowering-runnering decision in diploid strawberry. Plant Cell 29, 2168–2182 (2017).
Liu, Q. et al. G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice. Nat. Commun. 9, 1–12 (2018).
Peng, S., Khush, G. S., Virk, P., Tang, Q. & Zou, Y. Progress in ideotype breeding to increase rice yield potential. F. Crop. Res. 108, 32–38 (2008).
Sambrook, J., Fritsch, E. F. & Maniatis, T. Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory, New York, 1989).
Kawamoto, T. Use of a new adhesive film for the preparation of multi-purpose fresh-frozen sections from hard tissues, whole-animals, insects and plants. Arch. Histol. Cytol. 66, 123–143 (2003).
Kouchi, H. & Hata, S. Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Mol. Gen. Genet. 238, 106–119 (1993).
Ozawa, K. A high-efficiency agrobacterium-mediated transformation system of rice (Oryza sativa L.). Methods Mol. Biol. 847, 51–57 (2012).
Kojima, M. et al. Highly sensitive and high-throughput analysis of plant hormones using ms-probe modification and liquid chromatography–tandem mass spectrometry: an application for hormone profiling in Oryza sativa. Plant Cell Physiol. 50, 1201–1214 (2009).
Asano, K. et al. Artificial selection for a green revolution gene during japonica rice domestication. Proc. Natl Acad. Sci. USA 108, 11034–9 (2011).
Minami, A. et al. Time-Course transcriptomics analysis reveals key responses of submerged deepwater rice to flooding. Plant Physiol. 176, 3081–3102 (2018).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Nozue, K. et al. Shade avoidance components and pathways in adult plants revealed by phenotypic profiling. PLoS Genet. 11, 1–26 (2015).
Acknowledgements
We thank M. Sato, R. Ishihara, and M. Inari for helping us analyze panicle branching patterns. This work was supported by a grant from the Ministry of Agriculture, Forestry, and Fisheries of Japan (Genomics-based Technology for Agricultural Improvement, RBS-2002). This work was supported by the Grant-in-Aid for Scientific Research on Innovative Areas (JP16H06468 to M.U.-T.). This research was supported by the Integrative Graduate Education and Research Program in Green Natural Sciences of Nagoya University.
Author information
Authors and Affiliations
Contributions
A.A., T.H., and H.K. designed the research; A.A., K.A., S.O., M.K., Y.T., S.T., K.D., M.U.-T., T.S., H.S., M.M., M.A., Y.I., H.K., and T.H. performed the experiments; K.A. and T.H. performed the positional cloning experiments; A.A., K.A., and T.H. analyzed the data; A.A., T.H., and H.K. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Agata, A., Ando, K., Ota, S. et al. Diverse panicle architecture results from various combinations of Prl5/GA20ox4 and Pbl6/APO1 alleles. Commun Biol 3, 302 (2020). https://doi.org/10.1038/s42003-020-1036-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-020-1036-8
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
