
Abstract
Inactivating mutations affecting key mismatch repair (MMR) components lead to microsatellite instability (MSI) and cancer. However, a number of patients with MSI-tumors do not present alterations in classical MMR genes. Here we discovered that specific missense mutations in the MutL homolog MLH2, which is dispensable for MMR, confer a dominant mutator phenotype in S. cerevisiae. MLH2 mutations elevated frameshift mutation rates, and caused accumulation of long-lasting nuclear MMR foci. Both aspects of this phenotype were suppressed by mutations predicted to prevent the binding of Mlh2 to DNA. Genetic analysis revealed that mlh2 dominant mutations interfere with both Exonuclease 1 (Exo1)-dependent and Exo1-independent MMR. Lastly, we demonstrate that a homolog mutation in human hPMS1 results in a dominant mutator phenotype. Our data support a model in which yeast Mlh1-Mlh2 or hMLH1-hPMS1 mutant complexes act as roadblocks on DNA preventing MMR, unraveling a novel mechanism that can account for MSI in human cancer.
Similar content being viewed by others
Introduction
The DNA mismatch repair (MMR) system corrects mispaired nucleotide bases that arise on DNA1,2,3,4. DNA mispairs occur during DNA synthesis and escape the proofreading action of replicative DNA polymerases; others are introduced by error-prone DNA polymerases at sites of DNA damage or are caused by enzymatic or chemical modifications of nitrogen bases on DNA. Importantly, inactivating mutations (or epigenetic silencing) of key MMR components result in elevated mutation rates and cancer predisposition5,6,7.
In eukaryotes, the recognition of mispaired bases is performed by three MutS homologs (MSH) (Msh2, Msh3, and Msh6) that form two heterodimeric complexes, Msh2–Msh6 and Msh2–Msh3 (also referred to as MutSα and MutSβ, respectively), with partially redundant substrate specificity8.
Additional members of the MMR family are the MutL homolog (MLH) proteins, which are represented in S. cerevisiae by Mlh1, Pms1, Mlh2, and Mlh3. Yeast Mlh1 heterodimerizes with other MLH subunits forming MutLα (Mlh1–Pms1), MutLβ (Mlh1–Mlh2), and MutLγ (Mlh1–Mlh3). In humans, MutLα, MutLβ, and MutLγ heterodimers are represented by hMLH1–hPMS2, hMLH1–hPMS1, and hMLH1–hMLH3, respectively.
In eukaryotes, MutLα is critical for MMR function, whereas MutLβ and MutLγ play a limited role in MMR1,9,10. In S. cerevisiae Mlh1–Mlh2 is recruited to the mispair site and facilitates the MMR reaction in specific situations11, while Mlh1–Mlh3 acts mainly during meiosis promoting the resolution of recombination intermediates12,13. MLH proteins are homologs to the E. coli MutL (EcMutL) MMR protein, and share a related structure consisting of an N-terminal domain (NTD) that possesses ATPase activity, an unstructured linker, followed by a C-terminal domain (CTD) that is necessary for the dimerization. The NTDs of MLH subunits can also dimerize forming a ring-like structure that has been proposed to encircle the DNA3,4. Importantly, the CTD of MutLα and MutLγ, but not MutLβ, possess endonuclease domains that allow these complexes to nick DNA14.
After recognition of mispaired bases, the MutS complex promotes the recruitment of MutLα, which introduces a nick into the newly synthesized strand in the proximity of the mispaired base. In humans, MutLα endonuclease activity is stimulated by the interaction with the Proliferating Cell Nuclear Antigen (PCNA)15 (called Pol30 in S. cerevisiae). Next, the DNA fragment containing the mispaired base is excised either by Exonuclease 1 (Exo1) or in an Exo1-independent manner16. After excision, high-fidelity DNA polymerases re-synthesize the excised DNA fragment and the remaining nick is subsequently sealed by DNA ligase I17.
Mutations inactivating key components of the human MMR system (hMLH1, hMSH2, hMSH6, or hPMS2) are responsible for the most common hereditary cancer predisposition syndrome referred to as Lynch Syndrome or hereditary non-polyposis colorectal cancer (HNPCC)5,6,7. Lynch Syndrome patients are at risk of early onset of cancer due to the accumulation of mutations, especially at repetitive sequences causing microsatellite instability (MSI). However, a significant fraction of colorectal cancer patients with MSI tumors do not present mutations or altered protein expression levels at any of the major MMR components5, suggesting that additional factors may contribute to either MMR function or DNA replication fidelity at repetitive sequences. Along this line, a previous study demonstrated that inactivation of SETD2, the histone methyltransferase that promotes H3K36 trimethylation, causes MSI by preventing the recruitment of MutSα to chromatin18.
Campbell et al.11 previously showed that overexpression of either MLH2 or MLH3 genes in S. cerevisiae completely inactivates MMR function, most likely by outcompeting Pms1 for Mlh1 binding, preventing the assembly of Mlh1–Pms1 complexes that are indispensable during MMR. In addition, this work showed that similar to Mlh1–Pms119,20, Mlh1–Mlh2 is recruited in vitro by Msh2–Msh6 to DNA containing a mispaired base. Furthermore, Mlh1–Mlh2 forms short-lived nuclear foci that colocalize with Pms1 foci at sites of repair in vivo11. Inactivation of Mlh2 in a wild-type (WT) background causes no significant changes in mutation rates10,11, however, it facilitates MMR when Mlh1–Pms1 endonuclease function is partially compromised11. Based on these previous findings, we investigated whether specific MLH2 mutations could compromise MMR function, potentially by preventing Mlh1–Pms1 complex assembly, inhibiting MMR steps downstream mispair recognition or by alternative mechanisms. Using a genetic screen in budding yeast, we identified a group of dominant MLH2 missense mutations, all of them affecting residues at the N-terminus of Mlh2 causing an increase in frameshift mutation rates up to 1000-fold in the lys2-10A assay in which a MMR defective strain shows 7000-fold higher rates compared to WT. Further characterization revealed that Mlh1–Mlh2 mutant complexes are recruited to mispair sites where they accumulate, acting as roadblocks on DNA preventing MMR. Finally, we showed that one homolog mutation introduced into hPMS1 (the human homolog of yeast MLH2) causes a dominant mutator phenotype in human cells, suggesting that these type of mutations can lead to increased mutagenesis and cancer predisposition.
Results
Identification of MLH2 dominant mutations resulting in a mutator phenotype
To search for MLH2 mutations that could compromise MMR function, we screened a low-copy plasmid library of randomly mutagenized MLH2, for a dominant mutator phenotype when expressed in a haploid WT strain. The yeast transformants were screened for a mutator phenotype using two frameshift reversion reporters (lys2-10A and hom3-10) and the CAN1 forward inactivation assay21,22. The lys2-10A and the hom3-10 reporters are sensitive to single nucleotide deletions that occur in a well-defined mononucleotide run, resulting in lysine (Lys+) and threonine (Thr+) prototrophic colonies, respectively. In contrast, the CAN1 inactivation assay reports inactivating base substitutions, insertions, deletions, and chromosomal rearrangements resulting in canavanine resistance (CanR). After screening ~71,000 transformants, we identified four Mlh2 mutant alleles resulting in a mutator phenotype: mlh2-S16P, mlh2-S18P, mlh2-P332L, and the double mutant mlh2-S16P-D219G (Fig. 1a). Interestingly, all mutations located at Mlh2-NTD, which harbors the ATPase domain, and three of them (S16P, S18P, and P332L) affected residues that are part of two disordered loops located at the interface between the two NTDs of the dimer23,24. Ser16 and Ser18 are part of the loop 1 (L1) (Fig. 1a), an unstructured region that upon ATP binding adapts an ordered conformation, extending away and interacting with the ATP-binding site of the second subunit of the dimer24 (Fig. 1b, c), while Pro332 is a conserved amino acid located in loop 3 (L3) at the interface with Mlh1-NTD (Fig. 1b, c). In addition we identified mutated Asp219, which localizes in helix αF that is predicted to be part of a linker (residues 211–233) that connects the ATPase domain (residues 20–210) with the rest of the NTD (residues 234–359)23.

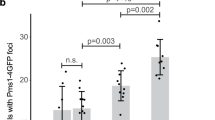
a Schematic representation of the Mlh2 protein. Mutated residues are indicated with arrows. Red boxes represent conserved ATPase motifs (I–IV) in MutL homologs. Below, protein sequence alignment of S. cerevisiae Mlh2 and MutL homologs in H. sapiens, M. musculus, and E. coli MutL. Residues affected by mutations are marked in red. Conserved hydrophobic residues are shaded in yellow, basic in blue, acidic in orange, and others in purple. * denotes invariable residue across species. Disordered loops L1 and L3 are indicated. b Model of Mlh1–Mlh2 heterodimer (Mlh1 in gray, Mlh2 in orange) based on E. coli MutL-NTD crystal structure (PDB: 1b62). Mutated residues are indicated as blue spheres. Relevant S. cerevisiae amino acid numbers are indicated, followed by the EcMutL homolog residue numbers in parentheses. ADP is indicated in red/gray and Mg2+ metal ion in green (only in the Mlh2 monomer). c Top view of the heterodimer structure shown in b. Models shown in b and c were made using Protean 3D, Lasergene 15.1, DNASTAR. d Mutation rate analysis, represented as box plots with whiskers, using the lys2-10A frameshift reversion assay of strains carrying mlh2 dominant mutations integrated at the chromosomal locus. Numbers on top indicate fold increase in the mutation rate relative to the WT strain. Black dots indicate outliers. e Box plots with whiskers indicate the percentage cells containing Pms1-4GFP foci. Black and red lines indicate median and average, respectively. Black dots represent outliers. n = total number of counted cells. ***p < 0.001; n.s not significant.
Expression of these mlh2 mutant alleles (including mlh2-V15P and mlh2-S17P, that were generated by site-directed mutagenesis) on a low copy plasmid in a WT strain resulted in elevated mutation rates on the lys2-10A reporter, compared to strains expressing WT Mlh2 (Supplementary Table 1). In some cases, a small increase in mutation rates was observed with the less sensitive hom3-10 frameshift reporter, and no major changes were detected in the CAN1 forward inactivation assay, likely due to the low sensitivity of this reporter for frameshift mutations (Supplementary Table 1). Similar results were obtained when mlh2 mutations were introduced at MLH2’s chromosomal locus (Fig. 1d and Supplementary Table 2). Furthermore, most of these mutations resulted in an increased abundance of Pms1 foci (Fig. 1e and Supplementary Fig. 1a), which mark sites of repair and accumulate when downstream steps of the MMR reaction are compromised20,25,26. Logarithmic cultures of strains expressing mlh2 mutations showed similar Mlh2 protein expression levels as a WT strain (Supplementary Fig. 1b), ruling out the possibility that the mutator phenotype was caused by increased Mlh2 protein levels, which would outcompete Pms1 for Mlh1 binding11.
Identification of MLH2 mutations acting as mlh2-S16P mutational enhancers
Among the isolated mlh2 mutant alleles, the double mutant mlh2-S16P-D219G showed the highest mutation rate (737-fold increase in the lys2-10A reporter) and the strongest accumulation of Pms1 foci (9 times higher than WT levels) (Fig. 1d, e). Instead, the D219G mutation by itself neither caused a mutator phenotype (Supplementary Table 2) nor resulted in increased Pms1 foci abundance (Supplementary Fig. 1a). These results suggest that S16P and D219G mutations affect different processes of the Mlh1–Mlh2 complex behavior, and the D219G mutation acts as an enhancer of S16P mutator phenotype, causing a synergistic increase in the mutation rate.
To test whether other MLH2 mutations could further enhance mlh2-S16P mutator phenotype, we performed an additional screen where we randomly mutagenized mlh2-S16P and searched for transformants that grew in at least two of the mutator reporter plates. Among ~37,000 transformants, we identified six distinct mlh2-S16P enhancer mutations: D45N, E99K, F177L, D178N, E216G, T217A and the D219G mutation previously identified. All isolated plasmids contained missense mutations affecting residues at Mlh2-NTD (Fig. 2a, b); most of them (with exception of E99K) were predicted to locate at the surface of the protein. Asp45 is part of the β1 sheet flanking the first ATPase motif (Supplementary Fig. 2a, b), and its side chain is likely exposed to solvent. Glu99 is immersed in a conserved region among MutL homologs, characterized by the “GFRGEAL” sequence, which is part of the ATP-binding motif III2,23 (Supplementary Fig. 2a). The remaining amino acid substitutions clustered in two regions: Phe177 and Asp178 located along the helix αE in EcMutL structure23, a region with a relative conservation in hydrophobic residues among MutL homologs (Supplementary Fig. 2a), and residues Glu216, Thr217, and Asp219 clustered in fairly well-exposed linker region with low amino acid conservation (Fig. 2a, b and Supplementary Fig. 2a) that connects the ATPase domain with the rest of the NTD23.

a Model of the Mlh1–Mlh2 heterodimer structure (Mlh1 in gray, Mlh2 in orange) indicating residues affected by mutations with arrows (purple spheres). Mlh2 amino acid numbers are shown followed by EcMutL homolog residue numbers in parentheses. b Top view of the Mlh1–Mlh2 heterodimer shown in a. c Frameshift mutation rates (lys2-10A reporter) of strains carrying double mlh2 mutations integrated at the chromosomal locus shown in box plots with whiskers. Numbers on top of each box indicate the fold increase in the mutation rate over the WT and black dots represent outliers. d CAN1 mutation spectrum in the indicated strains. Independent canavanine-resistant (CanR) colonies (n ≥ 90 per genotype) were sequenced for CAN1 mutations. Pie graphs show the relative distribution of identified mutations. e Confocal live-cell images of Mlh2-4GFP foci (examples are shown with white arrows) in logarithmically growing cells with the indicated genotype. Bar represents 5 μm. f Quantification of Mlh2-4GFP foci in the indicated strains, shown as box plots. The black and red lines indicate median value and average, respectively. Black dots represent outliers. n indicates the total number of cells counted per genotype. Fold intensity shows the fold increase of the foci intensity of the mlh2 mutants over the WT foci intensity. ***p < 0.001.
Yeast strains carrying double mlh2 mutations showed in average a 12-fold higher mutation rate than strains with the S16P single mutation (Fig. 2c and Supplementary Tables 1 and 2). Analysis of the CAN1 mutation spectra in these strains revealed that more than 60% of the CAN1-inactivating mutations were single nucleotide deletions (and few insertions) mainly at mononucleotide runs (4–6 bases); whereas the WT mutation spectrum was dominated by base substitutions (75%), with a small fraction (16%) of frameshifts (Fig. 2d and Supplementary Table 4). The preponderance of frameshifts over base substitutions in mlh2 double mutants is indicative of a severe MMR defect, similar as described for an msh2Δ strain8,27. In line with the effect on mutation rates, strains carrying double mlh2 mutations (except for S16P-D45N) showed a significantly higher percentage of cells containing Mlh2 foci compared to the mlh2-S16P strain (p < 0.001) (Fig. 2e, f).
Strains expressing double mlh2 mutations showed in average 2–3 times brighter Mlh2 foci than WT (Fig. 2f), which was not caused by an evident increase in Mlh2 (or Pms1) protein levels (Supplementary Fig. 2b). In addition, strains expressing mlh2-S16P-D219G allele showed Pms1 foci that in average persisted 8-times longer and were 2.9-times brighter than WT Pms1 foci (Fig. 3a, b, c).

a Representative time-lapse images following Pms1-4GFP foci in the WT and mlh2-S16P-D219G double mutant strain, over a 20 min period. Images were captured every minute. Arrowheads follow a focus in time; bar represent 5 μm. Brightfield images were taken at the beginning of the time-lapse. b Quantification of Pms1 focus duration in the WT and mlh2-S16P-D219G double mutant. c Intensity of Pms1 foci was calculated in strains with indicated mlh2 mutation. Fold intensity shows the fold increase of the foci intensity (averaged value) of the mlh2 mutants compared with the WT. In b and c, the data are presented as box plots with whiskers, in which the black and red lines indicate the mean and the average, respectively. Black dots represent outliers. d Pms1 and Mlh2 protein expression in the WT and mlh2-S16P-D219G double mutant strains analyzed by Western blot. Lysates were prepared with α-factor synchronized cells that were released at different time points, as indicated. Data are representative of two independent experiments, which gave similar results. Sic1 and Clb2 were used as G1- and G2/M-phase markers, respectively. Pgk1 was used as a loading control. ***p < 0.001; **p < 0.01; n.s indicates not significant compared to WT.
Analysis of Pms1 protein levels throughout the cell cycle revealed a transient S-phase-dependent expression that peaks at 30 min after release from α-factor arrest (Fig. 3d), and is consistent with Pms1 mRNA levels during cell cycle28,29. Similarly, Mlh2 protein expression was highest during S phase. Interestingly, strains expressing the mlh2-S16P-D219G allele showed for both, Pms1 and Mlh2 proteins, a less-tight S-phase-dependent expression pattern (Fig. 3d). Both proteins accumulated at earlier time points and remained detectable until beginning of G2/M (indicated by the expression of the Clb2 cyclin). These results are in agreement with the more abundant and long-lived Pms1/Mlh2 foci observed in strains carrying mlh2 mutant alleles.
The mlh2-S16P mutation results in a stronger Mlh1–Mlh2 interaction by yeast two-hybrid
As Ser16 locates at the NTD–dimer interface (Fig. 1b, c)23,24, we explored the possibility whether this mutation could have an impact on the interaction between Mlh2 and Mlh1. To test this idea, we took advantage of the yeast two-hybrid (Y2H) system, which has been used to detect the interaction between Pms1 and Mlh130,31. Growth on reporter plates (Trp− Leu− His−) (Fig. 4a), revealed a positive interaction between Pms1 and Mlh1, and a very weak interaction between Mlh2 and Mlh1, despite Mlh2’s higher expression level (Supplementary Fig. 3a). Remarkably, the Mlh2-S16P mutant protein interacted with Mlh1 much stronger than WT Mlh2 (Fig. 4a), which is likely caused by the Ser to Pro substitution that will affect the orientation of the first N-terminal 16 residues. This result suggests that the S16P mutation is either promoting the association with Mlh1, or preventing the dissociation between Mlh1- and Mlh2-NTDs. Furthermore, strains carrying double mlh2 mutations caused a similar increase in the Y2H interaction, suggesting that mlh2-S16P enhancer mutations are interfering with MMR using a different mechanism.

a The interaction between Mlh1 (prey) and WT- or mutant Mlh2 (baits) proteins was tested by Y2H. Prey and bait plasmids contain TRP1 and LEU2 auxotrophic markers, respectively. Interaction was scored as growth on Trp− Leu− His− +1 mM 3- AT plates. Cells were spotted in serial dilutions on control (Trp− Leu−) or reporter plates (Trp− Leu− His− +1 mM 3-AT). Mlh1 and Pms1 proteins serve as positive control for the Y2H interaction. b–d Box plots with whiskers showing mutation rates in the indicated strains using the lys2-10A frameshift reporter. The numbers on top correspond to the fold increase in mutation rates over the WT. Black dots indicate outliers.
mlh2-S16P mutation interferes with Exo1-dependent and Exo1-independent MMR pathways
To gain further insight into the mechanism how mlh2 mutations prevent MMR function, we generated double mutant strains carrying the mlh2-S16P (or mlh2-S16P-D219G) allele and a mutation that either inactivates the Exo1-dependent or Exo1-independent MMR pathways16. In agreement with previous reports, inactivation of Exo1 (exo1Δ) resulted in a modest mutator phenotype20,26,32 (Supplementary Table 3a). However, exo1Δ mutation in combination with either mlh2-S16P or mlh2-S16P-D219G mutant allele caused a strong increase in frameshift mutation rates (Fig. 4b and Supplementary Table 3a). These results indicate that the mlh2-S16P mutation is interfering with Mlh1–Pms1 nicking activity, which normally compensates for the lack of Exo1. Moreover, combining the mlh2-S16P allele with mutations that inactivate the Exo1-independent pathway (pol30-K217E or pms1-A99V) by preventing Mlh1–Pms1 nicking activity also caused a synergistic increase in frameshifts (Fig. 4b and Supplementary Table 3a). Together, these results revealed that the mlh2-S16P mutation causes a partial defect in both, Exo1-dependent and Exo1-independent MMR pathways.
The accumulation of Mlh2 foci in strains carrying the mlh2-S16P mutant allele, suggests that the mlh2 mutant complexes are being recruited to mispair sites on DNA. Based on the increased Y2H interaction between Mlh2-S16P and Mlh1, we speculated that the Mlh1–Mlh2 mutant complexes may remain associated with DNA, acting as roadblocks interfering with the MMR reaction. To test if the mutator phenotype caused by mlh2 mutations requires the interaction of Mlh1–Mlh2 mutant complexes with DNA, we introduced the K294E mutation into the mlh2-S16P-D219G allele, that is expected to prevent DNA binding33,34. Remarkably, the K294E mutation completely suppressed the mutator phenotype (Fig. 4c and Supplementary Table 3b) and the accumulation of Mlh2 foci, characteristic of the mlh2-S16P-D219G allele (Supplementary Fig. 3b). A similar result was seen after introducing the E29A mutation that prevents MutL-dependent ATP hydrolysis23 (Fig. 4b, Supplementary Table 3b and Supplementary Fig. 3b). These effects were not caused by a destabilization of the protein, as both triple mutants were expressed at similar levels than mlh2-S16P-D219G protein (Supplementary Fig. 3c). In addition, we found out that the K294E mutation, but not the E29A mutation, completely abolished the Y2H interaction between mlh2-S16P-D219G and Mlh1 (Fig. 4a and Supplementary Fig. 3a). It is interesting that the K294E mutation also suppressed the dominant mutator phenotype (Supplementary Table 3b) and the Mlh2 foci accumulation (Supplementary Fig. 3b) associated with the mlh2-S16P-D219G mutation more efficiently than the E29A mutation. Together, these observations indicate that Mlh1–Mlh2 mutant complexes impart a dominant mutator phenotype, most likely by preventing downstream steps of the MMR reaction.
We also found that overexpression of POL30 or PMS1 (by replacement of their endogenous promoters with a strong constitutive pGPD promoter), largely suppressed the mutator phenotype and the accumulation of Mlh2 foci associated with the mlh2-S16P-D219G allele (Fig. 4c, Supplementary Table 3b and Supplementary Fig. 3b). The suppression caused by Pms1 overexpression (Pms1-OE) is in agreement with the fact that the Mlh2 mutant protein has to compete with Pms1 for Mlh1 binding. On the other hand, the suppression by Pol30 overexpression (Pol30-OE), and the synergistic increase in mutation rates after combining mlh2-S16P with exo1Δ, both indicates that Mlh1–Mlh2 mutant complexes prevent the interaction between Pol30 and Mlh1–Pms1 that is required for the activation of the Mlh1–Pms1 endonuclease.
It is tempting to speculate that this interaction may also trigger a conformational change in Mlh1–Pms1 (and potentially Mlh1–Mlh2) that facilitates the unloading of these complexes from DNA. This idea is consistent with the accumulation of Pms1 foci in yeast strains carrying Pol30 mutant alleles that are defective in the activation of the Mlh1–Pms1 endonuclease26, but also with the suppression of mlh2-S16P-D219G mutator phenotype and the reduction in Mlh2 foci abundance observed upon Pol30-OE.
As the S16P mutation in Mlh2 disturbs a fairly well-conserved residue among MutL homologs, we asked whether homolog mutations in yeast Pms1 or Mlh1 might compromise their function. Mutation rate analysis in strains carrying the Pms1 homolog mutation (pms1-S17P) showed an increased mutator phenotype, whereas strains containing the Mlh1 homolog mutation (mlh1-A18P) were completely MMR defective (Fig. 4d and Supplementary Table 3c). The fact that the homolog mutation is more deleterious in Mlh1 than in Pms1, is probably related to the asymmetry of both NTDs, being more critical the Mlh1-NTD ATPase function31,35. Moreover, these results suggest that the loop 1 plays a conserved and critical function during MMR among yeast MLH proteins.
In summary, these results support a model in which Mlh1–Mlh2 mutant complexes are loaded on DNA, where they remain associated for a longer period than WT complexes. We propose that these mutant protein complexes act as roadblocks on DNA preventing downstream steps of MMR.
The hPMS1-S14P mutation confers a dominant mutator phenotype in mammalian cells
Cumulative evidence indicates that inactivation of human PMS1 (hPMS1), the homolog of yeast Mlh2, is not associated with increased mutagenesis or cancer susceptibility6,7,36. Still, specific mutations in the hPMS1 gene could result in mutant protein complexes that may interfere with human MMR, analogous to our findings in S. cerevisiae. We used CRISPR-Cas9 gene editing to introduce the S14P point mutation (homolog mutation to yeast mlh2-S16P) into hPMS1 in human HAP1 cells (Supplementary Fig. 4). In addition, we generated an hMLH1-knockout (hMLH1-KO) HAP1 cell line that was used as MMR-deficient control, as well as a hPMS1-knockout cell line (hPMS1-KO) that was expected to be MMR-proficient according to previous studies6,37,38. The hPMS1-S14P mutation did not affect the overall stability of the protein (Fig. 5a). In contrast, inactivation of hMLH1 resulted in a strong reduction in hPMS1 levels, in agreement with the fact that MLH proteins are only stable as heterodimers.

a hPMS1 and hMLH1 expression levels in CRISPR/Cas9-edited hPMS1-S14P and hMLH1-KO cell lines, determined by Western blot. Actin was used as a loading control. b Qualitative mutator analysis showing increased resistance to 6-TG in hMLH1-KO and hPMS1-S14P cells compared to mock cells. Cells were treated with 6-TG at the indicated concentration for 7 days and were stained with crystal violet. c Survival curves for hMLH1-KO (n = 4, biologically independent clones) and hPMS1-S14P cells (n = 4, biologically independent clones) based on a colony-formation assay after 12 days of treatment with different concentrations of 6-TG. d Western blot analysis showing hMLH1 expression levels in the indicated cell lines. e Qualitative mutator analysis (as shown in b) for the indicated cell lines. f 6-TG survival curves for HAP1-mock (n = 6, biologically independent clones), hMLH1-KO (n = 4, biologically independent clones), and hMLH1-A21P (n = 3, biologically independent clones), determined as in c. For c and f, error bars represent standard deviation of the mean of the data set.
Next, we evaluated the mutator phenotype of HAP1 mock cells (transfected with a plasmid expressing a single guide RNA (sgRNA) targeting the green fluorescent protein (GFP)) and HAP1-mutant cells using the hypoxanthine-guanine phosphoribosyltransferase 1 (HPRT1) inactivation assay, which scores for HPRT1 mutations resulting in 6-TG resistance. Cells expressing a functional HPRT1 gene, convert 6-TG into toxic nucleotides that are incorporated into DNA, triggering G2/M arrest and cell death. In contrast, cells with a strong mutator phenotype, like MMR-deficient cells, frequently inactivate the HPRT1 gene and become resistant to 6-TG. Cells carrying the hPMS1-S14P mutation were more resistant to 6-TG than HAP1-mock cells, but not as resistant as hMLH1-knockout cells (Fig. 5b). In a quantitative colony formation assay at a concentration of 0.8 μM 6-TG, only 15% of mock-transfected cells remained alive, whereas 50% of the hPMS1-S14P mutant cells and 82% of hMLH1-knockout cells survived (Fig. 5c). On the other hand, HAP1 cells lacking the hPMS1 gene showed a similar sensitivity to 6-TG as the HAP1 mock-transfected cells (Supplementary Fig. 6), which is consistent with the minor role of hPMS1 in MMR37,38.
In budding yeast, strains carrying the mlh1-A18P mutation were completely MMR defective (Supplementary Table 3c). Similarly, we found that HAP1 cells carrying the hMLH1-A21P homolog mutation were as strong mutators as hMLH1-KO cells (Fig. 5e, f) and presented unchanged hMLH1 expression levels (Fig. 5d and Supplementary Fig. 5).
In summary, these results demonstrated that specific hPMS1 mutations (and potentially also homolog mutations in hMLH1) can confer a dominant mutator phenotype in human cells.
Discussion
Here we discovered missense mutations affecting the yeast MutL homolog Mlh2 or its human homolog hPMS1 that result in elevated mutation rates. The isolation of these mlh2 mutant alleles, together with the previous observation that deletion of the MLH2 gene in S. cerevisiae does not cause a significant mutator phenotype10,11 indicates that these mlh2 mutant alleles impart a dominant mutator phenotype by interfering with the function of other more relevant MMR proteins. The identified mlh2 mutant alleles did not affect Mlh2 or Pms1 protein levels; therefore, the underlying cause of the mutator phenotype is different from the one reported for Mlh2 or Mlh3 overexpressing strains11. All the identified mutations affected residues located exclusively at Mlh2’s NTD, and can be grouped in two categories: 1) “mlh2-mutators”, those mutations causing a mutator phenotype by itself (e.g., S16P, S18P, and P332L), and 2) “mlh2-S16P mutational enhancers”, which are not mutators per se, but caused a synergistic increase in mutation rates in combination with the mlh2-S16P allele (e.g., D45N, E99K, D178N, and D219G). Our screen revealed some similarities with a study done in E. coli that identified a group of MutL dominant negative mutations, causing a mutator phenotype in the presence of a WT MutL gene39. Interestingly, among the identified EcMutL dominant mutations were A16T, A16V, and P305L, which are affecting the homolog residues to Ser16 and Pro332 in Mlh2, respectively. On the other hand, this previous study39 reported a variety of additional EcMutL dominant mutations that were not found in our screen, despite the fact that our screen was largely saturated as several of the mlh2 mutations were identified more than once. These differences could be related to the fact that in E. coli there is only one homodimeric MutL complex, whereas in eukaryotes there are up to three heterodimeric MutL complexes.
In vivo EcMutL exists as homodimer, promoted by the interaction of EcMutL C-terminal domains, while the NTDs can dimerize in response to ATP binding23. The association of the NTDs has been proposed to result in a central channel that allows EcMutL to enclose the DNA helix3,24. Most likely, this step is a prerequisite for nicking of the newly synthesized strand near the mispair site. Along this line, mutations at the ATPase domain in EcMutL or yeast MutLα cause MMR defects39,40. Moreover, a number of Lynch Syndrome patients harbor mutations at, or nearby the ATPase domain (Supplementary Table 5), suggesting that these mutations compromise the ATPase activity preventing MMR function.
Visualization of MutLα complexes by atomic force microscopy revealed that MutLα go through an ATPase cycle, in which ATP binding promotes dimerization of the NTDs and an overall highly compacted state, whereas ATP hydrolysis favors the dissociation of the NTDs and a more relaxed state41. The ATP-driven conformational change results in the interaction between the N- and C-terminal domains, potentially influencing the nicking reaction at the MutLα C-terminus. These findings are in agreement with recent single-molecule imaging studies of EcMutS and EcMutL proteins that have shown that both EcMutS and EcMutL, in their ATP-bound state, form sliding clamps that diffuse along DNA to direct MMR42,43.
According to the EcMutL crystal structure23,24 (Fig. 1), mlh2 mutations (e.g., S16P, S18P, or P332L) are affecting residues located at two disordered loops at the interface of EcMutL NTDs. These loops that become structured upon ATP binding are predicted to act as conformational switches23,24,34,44. Given that the identified mlh2 mutator alleles (S16P, S18P, P332L) have in common a proline substitution, it is expected that these substitutions will drastically change the orientation of loops L1/L3; potentially affecting the ability to sense ATP hydrolysis at the contiguous NTD, which could prevent the dissociation of the NTDs. This prediction is supported by the observation that the mlh2-S16P mutation results in a stronger interaction with Mlh1 by Y2H (Fig. 4a), similar to mutations that prevent ATP hydrolysis in yeast Mlh1/Pms1 subunits31.
On the other hand, most mlh2-S16P-enhancer mutations (with exception of E99K) are affecting residues located at the surface of the NTD-MutL dimer, suggesting that these residues could participate in transient protein–protein interactions, either with components of the MMR pathway (e.g., Msh2-Msh6 or PCNA) or perhaps, proteins that could be involved in the recycling of MutL subunits. Interestingly, the mlh2-D45N enhancer mutation affects a residue located at the predicted interface according to the EcMutS/EcMutL crystal structure45, suggesting that the D45N mutation could alter the interaction between Mlh2 and MutSα (or MutSβ). However, this possibility is rather unexpected, since the homolog interaction in humans is predominantly mediated by the hMLH1 subunit46,47.
Strains expressing mlh2 mutant alleles show increased abundance of Mlh2- and Pms1 foci, similar to mutations preventing Pms1 endonuclease function or Exo1-dependent excision11,20,25,26. Furthermore, Pms1/Mlh2 foci were brighter and long lasting, suggesting that the mlh2 mutations result in mutant protein complexes that are loaded on DNA, where they remain associated longer than usual. Accordingly, the mutator phenotype and the increased abundance of Pms1/Mlh2 foci caused by the mlh2-S16P-D219G allele were both suppressed by mutations that either prevent DNA binding (K294E) or inhibit ATP hydrolysis (E29A). Both mutations are expected to prevent the loading of Mlh1–Mlh2 mutant complexes via two different mechanisms; K294E mutation is predicted to decrease DNA binding affinity, whereas the E29A mutation results in a constitutive ATP-bound closed conformation with dimerized NTDs.
The analysis of Pms1 protein expression throughout the cell cycle in a WT strain, revealed a transient S-phase-dependent expression, which is consistent with Pms1 mRNA levels28,29. Similarly, Mlh2 protein expression was highest during S phase. Interestingly, strains expressing mlh2-S16P-D219G mutant allele showed an altered Pms1 and Mlh2 expression pattern throughout the cell cycle. Although it is possible that the Mlh1–Mlh2 mutant complexes may affect PMS1/MLH2 gene expression, it is more likely that these mutant protein complexes are interfering with the degradation of Mlh2 and Pms1. We speculate that Mlh1–Mlh2 mutant complexes are preventing the unloading of Mlh1–Pms1 from DNA, a process that might be somehow coupled to the degradation of Pms1 and Mlh2 subunits, and perhaps Mlh1 recycling. Intriguingly, several components of the ubiquitin-proteasome pathway were previously found associated with hPMS2 and hPMS1 (the homologs of yeast Pms1 and Mlh2, respectively)48, suggesting that both subunits could be targets of proteasome-mediated degradation, possibly as a mechanism to restrict their availability to the time when the DNA is replicated.
The synergistic increase in the mutation rate observed after combining the mlh2-S16P allele with exo1Δ, but also with mutations that prevent Pms1 endonuclease activity (pol30-K217E and pms1-E99V) (Fig. 4b and Supplementary Table 3a), indicates that the mlh2-S16P mutation partially interferes with both Exo1-dependent and Exo1-independent MMR pathways. On the other hand, the mutator phenotype caused by the mlh2-S16P-D219G allele was suppressed by Pms1 or Pol30 overexpression. Increased Pms1 levels will most likely outcompete mutant Mlh2 for Mlh1 binding, while the suppression by Pol30-OE suggests that Mlh1–Mlh2 mutant complexes interfere with the PCNA-dependent activation of Pms1 endonuclease activity. These genetic interactions are compatible with a model in which Mlh1–Mlh2 mutant complexes act as roadblocks on DNA interfering with PCNA-dependent activation of Pms1 endonuclease and Exo1-dependent excision.
A previous mutational study in S. cerevisiae identified a number of conserved residues located at the surface of Pms1-NTD that when mutated resulted in an elevated mutator phenotype34. Some of these mutations decreased DNA binding, whereas others including R212E/K213E did not. Interestingly, Arg212 and Lys213 are located at the helix αF that connects the ATP binding region from the rest of the NTD that corresponds to the same region identified in the present study as one cluster of mlh2-S16P enhancer mutations (D216G, T217A, and D219G). Certainly, this similarity highlights the importance of this linker region during the ATP-binding dependent conformational change, in which several regions of the NTD become ordered and compacted.
Analysis of human cancer genome databases (International Society for Gastrointestinal Hereditary Tumors (InSIGHT), Clinically relevant variants (ClinVAr), and the Catalog of Somatic Mutations in Cancer (COSMIC)) revealed, for all here-identified yeast MLH2 mutations, homolog substitutions in human MLH genes (hMLH1, hPMS2, and hPMS1) (Supplementary Table 5). Among them, hMLH1-A21V (homolog to Ser16 in S. cerevisiae Mlh2) has been found in Lynch Syndrome patients and is classified as pathogenic, while the hMLH1-E23D mutation (homolog to Ser18 in S. cerevisiae Mlh2) as well as the hMLH1-P309L (homolog to Pro332 in S. cerevisiae Mlh2) have been classified as uncertain. The hMLH1-Q48P mutant (homolog residue to Asp45 in S. cerevisiae Mlh2) has been classified as deleterious, as caused protein instability, reduced interaction with hPMS2, and a dominant negative mutator effect when tested in yeast49. The mutant variants hMLH1-E102D/E102K (homolog to Glu99 in Mlh2) had no effect on protein stability but showed ~50% of WT activity in an in vitro MMR assay using mismatched DNA heteroduplex50.
Interestingly, among the somatic mutations reported in the COSMIC database, we identified one patient with chronic lymphocytic leukemia carrying the hPMS1-P312S mutation, which affects a residue homolog to Pro332 (also identified in our Mlh2 screen). However, the pathogenic potential of this hPMS1 mutation has not yet been investigated. As listed in Supplementary Table 5, additional homolog mutations to the mlh2 mutator/mlh2-S16P enhancer mutations have been identified in hPMS2 and hPMS1 in a variety of cancer types, suggesting that these mutations could result in mutator phenotypes and cancer susceptibility. For some of these human mutant variants there are functional studies indicating an impaired MMR function. However, most of these studies cannot distinguish between loss of function or dominant mutations. Our study points out to specific MLH2 mutations that confer a dominant mutator phenotype, and predicts that homolog mutations in other MutL homologs may have similar consequences. In part, this idea is supported by the increased mutator phenotype in human HAP1 cells carrying the hPMS1-S14P or the hMLH1-A21P mutation. Future studies should evaluate whether a subset of the hMLH1 and hPMS2 missense mutations found in cancer patients are acting dominantly, similar as MLH2 mutations identified in this work.
Clinically, in tumors displaying MSI without identified causative mechanism, i.e., absence of pathogenic mutations in hMLH1, hMSH2, hMSH6, or hPMS2 and absence of hMLH1 promoter methylation, the testing for somatic and germline mutations affecting hPMS1 should be considered, particularly in patients with a clinical history suggestive of Lynch syndrome.
In summary, here we unraveled a novel mechanism that accounts for reduced MMR function in yeast and human cells, caused by dominant missense mutations in Mlh2 and hPMS1, respectively. Our results support a model in which specific Mlh2 mutations prevent the unloading of Mlh1–Mlh2 complexes from DNA, acting as roadblocks that interfere with downstream steps of the MMR reaction. Further studies are necessary to understand, at the molecular level, the biological process(es) affected by the mutations here identified. Such studies may shed light on specific residues that may participate in conformational changes driven by ATP binding/hydrolysis or interactions with DNA, MMR, or DNA replication components.
Methods
Yeast strains and media
Strains used in this study (Supplementary Table 6) are derivatives of the S288c strain RDKY596420, with exception of strain AH109 (Clontech Laboratories) that was used for Y2H analysis. Strains were cultivated at 30 °C in yeast extract-peptone-dextrose media (YPD) or appropriate dextrose-containing synthetic dropout (SD) medium for selection of plasmids markers, lacking lysine (Lys−) or threonine (Thr−) (to select for lys2-10A or hom3-10 frameshift revertants, respectively), or SD medium lacking arginine (Arg−) supplemented with 60 mg/L canavanine, to select canavanine-resistant (CanR) mutants. 5-fluoroorotic acid (5-FOA, US Biological) plates were done in SD medium supplemented with 1 g/L 5-FOA. Antibiotics were used at the following final concentrations: 200 μg/mL geneticin (Santa Cruz Biotechnology), 300 μg/mL hygromycin B (Thermo Fisher Scientific), and 100 μg/mL nourseothricin (clonNAT, Werner BioAgents). Gene deletions and gene tagging were performed using standard PCR-based recombination methods51,52, followed by confirmation by PCR. Tags and junctions were confirmed by PCR and sequencing. Yeast strains carrying mutations in MLH1, MLH2, or PMS1 genes, were generated by pop-in/pop-out strategy using pRS306-based integrative vectors51, and were confirmed by sequencing.
Construction of plasmids used in yeast experiments
All plasmids used in this study are listed in Supplementary Table 7. The plasmid pHHB98 encodes the WT-MLH2 gene, including 1 kb of the MLH2 promoter and 300 bp of the terminator, cloned using the SacI and EcoRI sites in pRS31651. The MLH2 sequence was amplified from genomic DNA using primers 5′-CTA CGA GAG CTC ACA AAT GGA TTC ATT AGA TCT ATT AC-3′ and 5′-GAG TAC GAA TTC TAT ATT TAT GTG GAG TGA TCT TTG TC-3′. To generate pHHB157 (an integrative URA3 plasmid containing the WT-MLH2 gene), the MLH2 gene (including promoter and terminator sequences) was cut from pHH98 with SacI and EcoRI, and ligated into pRS306. Integrative plasmids encoding specific mutant mlh2 alleles were generated either by subcloning the mutant alleles identified in the screens or by site-directed mutagenesis. Integrative mlh2 plasmids were linearized with MluI for integration at the MLH2 locus.
The integrative plasmids pHHB270 (encoding the mlh1-A18P mutation) and pHHB240 (encoding the pms1-S17P specific mutation) were generated by site-directed mutagenesis of pRDK133825 and pRDK166725, respectively, followed by subcloning the mutant alleles at the StuI and XhoI sites in pRS306. pHHB270 and pHHB240 were linearized with NheI and MluI, respectively. The integrative plasmid pHHB252, encoding the pol30-K217E specific mutation, was generated by site-directed mutagenesis of the LEU2-integrative plasmid pRDK92526. pHHB252 was linearized with SacI for one-step replacement and Leu+ transformants were confirmed by sequencing.
MLH2 random mutagenesis screen
The plasmid library of randomly mutagenized MLH2 was generated by mutagenic PCR amplification and in vivo gap repair in the lig4Δ strain HHY6620 that is deficient in non-homologous end joining, similar as previously described22. Transformants were plated on SD plates lacking uracil (Ura−) and then replica-plated onto frameshift mutator reporter plates (SD Ura− Lys− and SD Ura− Thr− to test for increased lys2-10A and hom3-10 frameshift reversion mutations, respectively); and onto SD Ura− Arg− + 60 mg/L canavanine to identify CAN1 inactivation mutations8,32. Plasmids resulting in increased frameshift mutations and canavanine resistance were identified, sequenced for mlh2 mutations and retransformed in RDKY5964 for further analysis. To identify mlh2-S16P-dependent enhancer mutations, mlh2-S16P was randomly mutagenized by PCR and a screen was performed as described above.
Determination of mutation rates in S. cerevisiae
Mutation rates using the CAN1 inactivation assay and the frameshift reversion assays (lys2-10A and hom3-10) were determined by fluctuation analysis as previously described8,32. Each mutation rate was determined by using two biological isolates and at least 14 independent cultures. 95% confidence intervals were calculated for all fluctuation tests.
Preparation of yeast and mammalian cell lysates and immunoblotting
Whole-cell protein extracts of S. cerevisiae were generated as previously described20. Mammalian cell lysates were prepared in lysis buffer (10 mM Tris pH 7.5, 50 mM NaCl, 1% Triton X-100, 2 mM PMSF, and protease inhibitor cocktail Complete with EDTA (Roche). Lysates were analyzed on 8% or 10% SDS-PAGE followed by immunoblotting. The following antibodies were used: anti-HA (1:5,000, 3F10, Roche), anti-c-Myc (1:1,000, 4A6, Millipore), anti-Sic1 (1:10,000)53, anti-Pgk1 (1:20,000, 22C5D8, Invitrogen), anti-hMLH1 (1:1,000, BD-551091), anti-hPMS1 (1:1,000, sc-615, Santa Cruz), and anti-actin (1:5,000, A2228, Sigma). Western blots were developed using Immobilon Western Chemiluminscent HRP substrate (Millipore) and imaged using Super RX-N Fuji medical X-ray films (Fujifilm) or using Fusion Solo S (Vilber).
Live-cell imaging of Pms1 and Mlh2 foci in S. cerevisiae
To visualize the localization of Pms1 or Mlh2 proteins in living yeast cells we used strains expressing Pms1 or Mlh2 proteins tagged with a 4xGFP tag (a the C-terminus), strains which according to previous studies retain to large extent MMR proficiency11,20. Exponentially growing cells were processed and imaged as described20 using a Leica SP5 confocal microscope system with an Argon laser, an HCX PL APO 63x/1.4 aperture objective and a high resonance scanner detector at 8 kHz frequency. 10–15 Z stacks spaced 0.4 μm were projected using the maximum intensity in ImageJ for analysis. Three independent biological replicates per genotype were analyzed and a Mann–Whitney rank sum test was used to compare Pms1/Mlh2 foci in different genetic backgrounds. Time-lapse images were performed in a DeltaVision RT (Applied Precision) with an inverted microscope (IX70, Olympus) with a CoolSNAP HQ2 (Photometrics) camera and a plan Apo 100x (1.4 NA) oil immersion objective lens (Olympus). 20 Z stacks spaced 0.3 μm were deconvolved using SoftWoRx software. For this experiment, exponentially growing cells (HHY8072 or HHY8073) grown in complete synthetic medium (CSM) were plated on glass chambers coated with concanavalin A (1 mg/ml, C-2010 Sigma) and imaged at 30 °C for indicated times. Deconvoluted images were processed to obtain the maximal intensity projection with ImageJ, at each shown time point. The Nup49-mCherry was used as nuclear marker. To compare Pms1 foci intensity between different genotypes, a culture of a strain expressing WT Pms1-4GFP (and Sik1-mCherry) (RDKY7600)20 was mixed with culture of a mutant strain, e.g., mlh2-S16P-D219G Pms1-4GFP (HHY5326) in 3:1 ratio. Expression of the nucleolar marker Sik1 (tagged with mCherry) was used to identify WT Pms1-4GFP foci. GFP intensities were measured using ImageJ. Similarly, to measure Mlh2 foci intensity among strains with different genotypes, an MLH2-4GFP NIC96-mCherry strain (RDKY7905) was used mixed in a ratio 5:1 with a mutant strain (e.g., HHY8064 mlh2-S16P-D219G-4GFP).
CAN1 mutation spectra analysis
The CAN1 gene of at least 90 independent canavanine-resistant (CanR) yeast colonies was amplified from genomic DNA by PCR with Phusion high-fidelity DNA polymerase (Thermofisher) and primers 5′- GTT GGA TCC AGT TTT TAA TCT GTC GTC-3′ and 5′- TTC GGT GTA TGA CTT ATG AGG GTG-3′. PCR reactions were purified and sequenced (GATC Biotech). Sequences were analyzed with Lasergene 15.1 (DNASTAR).
Yeast two-hybrid analysis
Protein–protein interactions were tested using the Y2H system, using the strain AH109 (Clontech laboratories) (Supplementary Table 6). Briefly, AH109 strain was transformed with bait and prey plasmids, derivatives of pGBKT7 and pGADT7, respectively (Supplementary Table 7). Cells were grown overnight in SD media lacking tryptophan (Trp−) and leucine (Leu−) and were spotted in 10-fold serial dilutions on control plates (SD Trp− Leu−) and reporter plates (SD Trp− Leu− His− + 1 mM 3-amino-1,2,4-triazole (3-AT) (Sigma)). Plates were incubated at 30 °C for 3–4 days and imaged using GelDoc system (Bio-Rad). The expression of bait and prey proteins was confirmed by SDS-PAGE and immunoblotting using anti-HA and anti-Myc antibodies.
Generation of plasmids used for CRISPR-Cas9 editing in mammalian cells
Plasmids used in mammalian tissue culture experiments are listed in Supplementary Table 7. sgRNAs used for inactivation of the hMLH1 and hPMS1 genes or the introduction of hPMS1-S14P or hMLH1-A21P point mutations were designed with the Optimized CRISPR Design tool (http://crispr.mit.edu) and were cloned into pLentiCRISPR-puro (a gift from Feng Zhang, Addgene plasmid # 49535). This plasmid expresses a gene-specific sgRNA, the human codon-optimized Streptococcus pyogenes Cas9 protein and the puromycin N-acetyl transferase gene. The plasmid pHHB586 that contains the sgRNA-hMLH1 (5′-TGA TAG CAT TAG CTG GCC GC-3′) was used to target the hMLH1 gene. Plasmids pHHB484 that contains the sgRNA-hPMS1 (5′-TTC TCA GAT CAT CAC TTC GG-3′) and pHHB487 that contains the sgRNA-hPMS1 (5′-CAC AAG CGT AGA TGT TAA AC-3′) were used to target hPMS1. An sgRNA that targets GFP (5′-GGG CGA GGA GCT GTT CAC CG-3′) was similarly cloned into pLentiCRISPR-puro resulting in the plasmid pHHB761, which was used in control experiments (mock).
The donor plasmid pHHB730 that was used to introduce the hPMS1-S14P mutation contains a 5′ and 3′ homology arms of ~900 and ~300 bp long, respectively (relative to Ser14 coding sequence) (Supplementary Fig. 4). To construct pHHB730 a 1.3 kb DNA fragment of the hPMS1 gene (including part of 5′ UTR, exon 1, and part of a downstream intron) was PCR-amplified from genomic DNA isolated from HAP1 cells using primers (F) 5′ CTG ACT GGT ACC GTG CTT GTG GCA GAA TAT TGT GGA-3′ and (R) 5′ GGT GAC CTC GAG CCA TAT CTC TAT GTG TTA GCA-3′, and was cloned into pcDNA5 FRT/TO (Invitrogen) using KpnI and XhoI sites (underlined). This construct was then subjected to site-directed mutagenesis to introduce the S14P point mutation, a silent mutation (TGG to TCG) at the Protospacer adjacent motif (PAM) and a third silent mutation (TTACAA to TTATAA) that creates a PsiI cutting site ~100 bp upstream of the ATG start codon, used later for verification purposes. Finally, a hygromycin resistance cassette flanked by loxP sites was cloned at the intronic SwaI site located ~250 bp downstream of the Ser14 coding sequence in the donor plasmid. This hygromycin resistance cassette was obtained by PCR-amplification using pHHB689 as template and primers (F) 5′ CTGACT ATTTAAAT ATAACTTCGTATAGCATA CAT TAT ACG AAG TTA TAT ACG CGT TGA CAT TGA TTA TTG AC-3′ and (R) 5′- GGTGAC ATTTAAAT ATAACTTCGTATA ATG TAT GCT ATA CGA AGT TAT CAG AAG CCA TAG AGC CCA CC-3′ (SwaI sites underlined). Plasmid pHHB689 contains the hygromycin resistance gene cloned using the KpnI and XhoI sites in pcDNA5 FRT/TO (Invitrogen).
The hMLH1-A21P mutation was introduced into HAP1 cells using the donor plasmid pHHB690, which contains a 5′ and 3′ homology arms of ~1.2 kb and ~800 bp long, respectively (relative to Ala21 coding sequence) (Supplementary Fig. 5). The plasmid pHHB690 was constructed using a similar strategy as described for pHHB730. A ~2 kb fragment of the human hMLH1 gene (including part of the 5′ UTR, exon 1, and a downstream intron) was PCR-amplified from genomic DNA of HAP1 cells with primers (F) 5′-CTG ACT GGT ACC GCG TAG ATT CCT GTC AAT GCT CAG G-3′ and (R) 5′-GGT GAC CTC GAG CTC TCA GTC CCA TTG CCC TGA ATA G-3′ and was cloned using the BamHI and XhoI sites in pcDNA5 FRT/TO. Next, the A21P mutation and a silent mutation at the PAM (TGG to TGA, that creates an XmnI site used for verification purposes) were introduced by site-directed mutagenesis. Finally, a hygromycin resistance cassette flanked by loxP sites was introduced into an intronic SmaI site (136 bp downstream of the A21 coding sequence) in the donor plasmid. This loxP-flanked hygromycin gene resistance cassette was obtained by PCR amplification using the plasmid pHHB689 and the primers (F) 5′ CTG ACT CCCGGG ATAACTTCGTATA GCA TAC ATT ATA CGA AGT TAT ATA CGC GTT GAC ATT GAT TAT TGA C-3′ and (R) 5′ GGT GAC CCCGGG ATAACTTCGTATA ATG TAT GCT ATA CGA AGT TAT CAG AAG CCA TAG AGC CCA CC-3′, (SmaI sites are underlined).
Generation of hMLH1-KO, hPMS1-KO, hPMS1-S14P, and hMLH1-A21P human cell lines
hMLH1-KO cell lines were generated with the plasmid pHHB586. hPMS1-KO cell lines were generated with the plasmid pHHB484 and pHHB487. An sgRNA-GFP encoded in the pHHB761 plasmid was used in control (Mock) experiments. Plasmids containing the sgRNAs were transfected into HAP1 cells by nucleofection using the Neon Transfection System (Life Technologies). Briefly, 106 HAP1 cells were transfected with 2.5 μg of the corresponding sgRNA-carrying plasmid with 3 pulses at 1575 V for 10 ms. After transfection, cells were grown under puromycin selection (1 μg/mL) for 48 h, and then in the absence of the antibiotic for 10–12 days. Single clones were expanded and tested by Western blot for MLH1 expression and confirmed by sequencing analysis.
To generate HAP1 cells carrying the MLH1-A21P point mutation, cells were co-transfected (nucleofection) with pHHB586 and pHHB690 plasmids. HAP1 cells carrying the PMS1-S14P mutation were obtained after co-transfection with pHHB484 and pHHB730 plasmids. After 48 h post transfection, cells were selected with puromycin (1 μg/mL) for 48 h, recovered for 24 h, and finally selected with hygromycin (700 μg/mL, Sigma) for 10–12 days. The loxP-flanked hygromycin resistance cassette was excised by CRE-mediated site-specific recombination. Briefly, 106 cells were transfected (nucleofection) with 2.5 μg CRE mRNA. Cells were grown without antibiotics for 10–12 days. Single clones were expanded and tested for loss of the hygromycin resistance. Finally, positive clones were identified by PCR and confirmed by sequencing analysis. All cells used in this study were negative for mycoplasma infection according to tests performed regularly (GATC, Eurofins Genomics).
Hypoxanthine phosphoribosyl transferase 1 (HPRT1) inactivation assay
HAP1 cells were usually grown in IMDM media supplemented with 10% FBS, in 5% CO2 at 37 °C. However, for mutator phenotype analysis using the HPRT1 inactivation assay, cells were grown for 7–10 days (passaged every 48 h) in hypoxanthine-aminopterin-thymidine (HAT)-supplemented medium (Thermo Fisher Scientific) to purge pre-existing 6-TG resistant cells. After HAT treatment, cells were recovered for 2–3 passages. For the qualitative HPRT1 inactivation assay, 104 cells of each clone were seeded in a 48-well plate in IMDM medium supplemented with 10% FBS. After 24 h, 6-TG was added and cells were grown for 7 days (medium was changed every 2–3 days). Finally, cells were fixed with ice-cold 20% methanol and were stained with 0.02% crystal violet (Sigma) in 20% methanol. A quantitative analysis of the 6-TG survival was determined with a colony formation assay by counting the number of 6-TG resistant colonies using four independent clones per genotype. Cells were plated at a density of 400 cells in 10 cm plates (in triplicate), in medium lacking 6-TG. After 24 h, 6-TG was added and cells were cultured for 12 days (medium was changed every 2–3 days). Plating efficiency was determined by plating 200 cells for each clone in 10 cm plates in media lacking 6-TG (in triplicate). At the end of the experiment, plates were stained, scanned and colonies were counted with ImageJ. The percentage of survival was calculated after counting the number of 6-TG resistant colonies (at each used concentration) and correcting for plating efficiency.
Statistics and reproducibility
To compare the percentage of nuclear Pms1/Mlh2 foci in different genetic backgrounds, three independent biological isolates per genotype were analyzed and a Mann–Whitney rank sum test was used for statistical analysis. P-values are indicated on the graphs and represent statistical significance of the difference between the two data groups. Mutation rates analysis in S. cerevisiae were determined using two independent biological isolates and a total of at least 14 independent cultures. The mutation rate data correspond to median rates for the indicated mutational reporters with 95% confidence intervals.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
References
Jiricny, J. Postreplicative mismatch repair. Cold Spring Harb. Perspect. Biol. 5, a012633 (2013).
Reyes, G. X., Schmidt, T. T., Kolodner, R. D. & Hombauer, H. New insights into the mechanism of DNA mismatch repair. Chromosoma 124, 443–462 (2015).
Kunkel, T. A. & Erie, D. A. Eukaryotic mismatch repair in relation to DNA replication. Annu. Rev. Genet. 49, 291–313 (2015).
Fishel, R. Mismatch repair. J. Biol. Chem. 290, 26395–26403 (2015).
Peltomaki, P. Role of DNA mismatch repair defects in the pathogenesis of human cancer. J. Clin. Oncol. 21, 1174–1179 (2003).
Boland, C. R. & Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 138, 2073–2087 e2073 (2010).
Boland, C. R. & Lynch, H. T. The history of Lynch syndrome. Fam. Cancer 12, 145–157 (2013).
Marsischky, G. T., Filosi, N., Kane, M. F. & Kolodner, R. Redundancy of Saccharomyces cerevisiae MSH3 and MSH6 in MSH2-dependent mismatch repair. Genes Dev. 10, 407–420 (1996).
Flores-Rozas, H. & Kolodner, R. D. The Saccharomyces cerevisiae MLH3 gene functions in MSH3-dependent suppression of frameshift mutations. Proc. Natl Acad. Sci. USA 95, 12404–12409 (1998).
Harfe, B. D., Minesinger, B. K. & Jinks-Robertson, S. Discrete in vivo roles for the MutL homologs Mlh2p and Mlh3p in the removal of frameshift intermediates in budding yeast. Curr. Biol. 10, 145–148 (2000).
Campbell, C. S. et al. Mlh2 is an accessory factor for DNA mismatch repair in Saccharomyces cerevisiae. PLoS Genet. 10, e1004327 (2014).
Zakharyevich, K., Tang, S., Ma, Y. & Hunter, N. Delineation of joint molecule resolution pathways in meiosis identifies a crossover-specific resolvase. Cell 149, 334–347 (2012).
Rogacheva, M. V. et al. Mlh1-Mlh3, a meiotic crossover and DNA mismatch repair factor, is a Msh2-Msh3-stimulated endonuclease. J. Biol. Chem. 289, 5664–5673 (2014).
Kadyrov, F. A., Dzantiev, L., Constantin, N. & Modrich, P. Endonucleolytic function of MutLalpha in human mismatch repair. Cell 126, 297–308 (2006).
Pluciennik, A. et al. PCNA function in the activation and strand direction of MutLalpha endonuclease in mismatch repair. Proc. Natl Acad. Sci. USA 107, 16066–16071 (2010).
Goellner, E. M., Putnam, C. D. & Kolodner, R. D. Exonuclease 1-dependent and independent mismatch repair. DNA Repair 32, 24–32 (2015).
Zhang, Y. et al. Reconstitution of 5′-directed human mismatch repair in a purified system. Cell 122, 693–705 (2005).
Li, F. et al. The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSalpha. Cell 153, 590–600 (2013).
Mendillo, M. L., Mazur, D. J. & Kolodner, R. D. Analysis of the interaction between the Saccharomyces cerevisiae MSH2-MSH6 and MLH1-PMS1 complexes with DNA using a reversible DNA end-blocking system. J. Biol. Chem. 280, 22245–22257 (2005).
Hombauer, H., Campbell, C. S., Smith, C. E., Desai, A. & Kolodner, R. D. Visualization of eukaryotic DNA mismatch repair reveals distinct recognition and repair intermediates. Cell 147, 1040–1053 (2011).
Hombauer, H., Srivatsan, A., Putnam, C. D. & Kolodner, R. D. Mismatch repair, but not heteroduplex rejection, is temporally coupled to DNA replication. Science 334, 1713–1716 (2011).
Schmidt, T. T. et al. A genetic screen pinpoints ribonucleotide reductase residues that sustain dNTP homeostasis and specifies a highly mutagenic type of dNTP imbalance. Nucleic Acids Res. 47, 237–252 (2019).
Ban, C. & Yang, W. Crystal structure and ATPase activity of MutL: implications for DNA repair and mutagenesis. Cell 95, 541–552 (1998).
Ban, C., Junop, M. & Yang, W. Transformation of MutL by ATP binding and hydrolysis: a switch in DNA mismatch repair. Cell 97, 85–97 (1999).
Smith, C. E. et al. Dominant mutations in S. cerevisiae PMS1 identify the Mlh1-Pms1 endonuclease active site and an exonuclease 1-independent mismatch repair pathway. PLoS Genet. 9, e1003869 (2013).
Goellner, E. M. et al. PCNA and Msh2-Msh6 activate an Mlh1-Pms1 endonuclease pathway required for Exo1-independent mismatch repair. Mol. Cell 55, 291–304 (2014).
Tishkoff, D. X., Filosi, N., Gaida, G. M. & Kolodner, R. D. A novel mutation avoidance mechanism dependent on S. cerevisiae RAD27 is distinct from DNA mismatch repair. Cell 88, 253–263 (1997).
Morrison, A., Johnson, A. L., Johnston, L. H. & Sugino, A. Pathway correcting DNA replication errors in Saccharomyces cerevisiae. EMBO J. 12, 1467–1473 (1993).
Kramer, W., Fartmann, B. & Ringbeck, E. C. Transcription of mutS and mutL-homologous genes in Saccharomyces cerevisiae during the cell cycle. Mol. Gen. Genet. 252, 275–283 (1996).
Prolla, T. A., Pang, Q., Alani, E., Kolodner, R. D. & Liskay, R. M. MLH1, PMS1, and MSH2 interactions during the initiation of DNA mismatch repair in yeast. Science 265, 1091–1093 (1994).
Tran, P. T. & Liskay, R. M. Functional studies on the candidate ATPase domains of Saccharomyces cerevisiae MutLalpha. Mol. Cell Biol. 20, 6390–6398 (2000).
Amin, N. S., Nguyen, M. N., Oh, S. & Kolodner, R. D. exo1-dependent mutator mutations: model system for studying functional interactions in mismatch repair. Mol. Cell Biol. 21, 5142–5155 (2001).
Hall, M. C. et al. DNA binding by yeast Mlh1 and Pms1: implications for DNA mismatch repair. Nucleic Acids Res. 31, 2025–2034 (2003).
Arana, M. E. et al. Functional residues on the surface of the N-terminal domain of yeast Pms1. DNA Repair 9, 448–457 (2010).
Hall, M. C., Shcherbakova, P. V. & Kunkel, T. A. Differential ATP binding and intrinsic ATP hydrolysis by amino-terminal domains of the yeast Mlh1 and Pms1 proteins. J. Biol. Chem. 277, 3673–3679 (2002).
Peltomaki, P. Epigenetic mechanisms in the pathogenesis of Lynch syndrome. Clin. Genet. 85, 403–412 (2014).
Raschle, M., Marra, G., Nystrom-Lahti, M., Schar, P. & Jiricny, J. Identification of hMutLbeta, a heterodimer of hMLH1 and hPMS1. J. Biol. Chem. 274, 32368–32375 (1999).
Peltomaki, P. & Vasen, H. Mutations associated with HNPCC predisposition – update of ICG-HNPCC/INSiGHT mutation database. Dis. Markers 20, 269–276 (2004).
Aronshtam, A. & Marinus, M. G. Dominant negative mutator mutations in the mutL gene of Escherichia coli. Nucleic Acids Res. 24, 2498–2504 (1996).
Tran, P. T., Simon, J. A. & Liskay, R. M. Interactions of Exo1p with components of MutLalpha in Saccharomyces cerevisiae. Proc. Natl Acad. Sci. USA 98, 9760–9765 (2001).
Sacho, E. J., Kadyrov, F. A., Modrich, P., Kunkel, T. A. & Erie, D. A. Direct visualization of asymmetric adenine-nucleotide-induced conformational changes in MutL alpha. Mol. Cell 29, 112–121 (2008).
Liu, J. et al. Cascading MutS and MutL sliding clamps control DNA diffusion to activate mismatch repair. Nature 539, 583–587 (2016).
Liu, J., Lee, J. B. & Fishel, R. Stochastic processes and component plasticity governing DNA mismatch repair. J. Mol. Biol. 430, 4456–4468 (2018).
Guarne, A., Junop, M. S. & Yang, W. Structure and function of the N-terminal 40 kDa fragment of human PMS2: a monomeric GHL ATPase. EMBO J. 20, 5521–5531 (2001).
Groothuizen, F. S. et al. MutS/MutL crystal structure reveals that the MutS sliding clamp loads MutL onto DNA. Elife 4, e06744 (2015).
Plotz, G., Raedle, J., Brieger, A., Trojan, J. & Zeuzem, S. N-terminus of hMLH1 confers interaction of hMutLalpha and hMutLbeta with hMutSalpha. Nucleic Acids Res. 31, 3217–3226 (2003).
Plotz, G. et al. Mutations in the MutSalpha interaction interface of MLH1 can abolish DNA mismatch repair. Nucleic Acids Res. 34, 6574–6586 (2006).
Cannavo, E., Gerrits, B., Marra, G., Schlapbach, R. & Jiricny, J. Characterization of the interactome of the human MutL homologues MLH1, PMS1, and PMS2. J. Biol. Chem. 282, 2976–2986 (2007).
Hardt, K. et al. Missense variants in hMLH1 identified in patients from the German HNPCC consortium and functional studies. Fam. Cancer 10, 273–284 (2011).
Takahashi, M. et al. Functional analysis of human MLH1 variants using yeast and in vitro mismatch repair assays. Cancer Res. 67, 4595–4604 (2007).
Sikorski, R. S. & Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 122, 19–27 (1989).
Janke, C. et al. A versatile toolbox for PCR-based tagging of yeast genes: new fluorescent proteins, more markers and promoter substitution cassettes. Yeast 21, 947–962 (2004).
Schmidt, T. T. et al. Inactivation of folylpolyglutamate synthetase Met7 results in genome instability driven by an increased dUTP/dTTP ratio. Nucleic Acids Res. 48, 264–277 (2020).
Acknowledgements
We thank to Dr. Michael Knop for generous support and to Dr. Richard Kolodner for sharing strains, plasmids, reagents, and advice. We thank Dr. Christopher Putnam and Dr. Thomas Kunkel for helpful discussions. We thank Dr. Haikun Liu, Dr. Elmar Schiebel, Dr. Marius Lemberg, and Dr. Thomas Hofmann for sharing reagents and protocols, and the DKFZ Imaging Core Facility and Daniel Kirrmaier (Knop lab) for their technical support. We thank Umran Ceylan for initial Y2H interaction analysis. This work was supported by the Harald zur Hausen fellowship from the Deutsches Krebsforschungszentrum and the Deutsche Forschungsgemeinschaft DFG grant HO-5501/1-1 (both granted to H.H.).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
H.H. conceived in large the overall experimental design. G.X.R., T.T.S., K.G., B.Z., and H.H. performed yeast strain and plasmid construction, and quantitative mutation rate measurements. G.X.R. performed microscopy analysis of Pms1 and Mlh2 foci. B.Z. generated HAP1 mutant cells and performed mutator phenotype analysis. M.K. supervised MSI analysis in human cells. G.X.R., B.Z., and H.H. wrote the paper; and all of the authors revised and modified the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Reyes, G.X., Zhao, B., Schmidt, T.T. et al. Identification of MLH2/hPMS1 dominant mutations that prevent DNA mismatch repair function. Commun Biol 3, 751 (2020). https://doi.org/10.1038/s42003-020-01481-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-020-01481-4
This article is cited by
-
MutS and MutL sliding clamps in DNA mismatch repair
Genome Instability & Disease (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
