
Abstract
Sustainable management of intact tropical peatlands is crucial for climate change mitigation, for biodiversity conservation and to support the livelihoods of local communities. Here, we explore whether sustainable fruit harvesting from Mauritia flexuosa palms could support these linked goals by increasing fruit production and incomes across the 2.8 million hectares of the most carbon-dense ecosystem in Amazonia: the lowland peatlands of northeastern Peru. M. flexuosa is dioecious, and fruits are typically harvested by felling female palms; the proportion of female palms therefore provides a good indicator of the health of a stand. Across 93 widely distributed sites, we found that the proportion of female palms increases with travel time to the urban market, and overall, fruit harvesting has halved the current potential production and income from this resource. However, significantly more female palms are found where fruit are harvested by climbing. We estimate that region-wide uptake of climbing could eventually increase potential fruit production by 51% and increase its gross value to US$62 ± 28.2 million yr–1. These findings demonstrate the high cost of unsustainable resource extraction in Neotropical forests and outline a practical path to conserve and sustainably exploit one of the most carbon-rich landscapes on the planet.
Similar content being viewed by others
Main
Tropical peatlands store approximately 104 billion tonnes of carbon1 (PgC), harbour unique combinations of flora and fauna and provide a wide range of resources for indigenous and local communities2,3,4. Sustainable management of these ecosystems is therefore crucial for addressing three major interlinked challenges: mitigating climate change, conserving biodiversity and supporting livelihoods of forest-dwelling peoples. However, the integrity of these ecosystems is threatened by fragmentation from infrastructure development and agricultural expansion5,6 while logging and unsustainable harvesting of non-timber forest products (NTFPs) are reducing the ability of peatland forests to supply resources that support local communities7,8. To support sustainable management and conserve the carbon stocks and biodiversity of tropical peatlands, it is important to quantify the services that these forests provide for people. In particular, it is crucial to understand how historical resource extraction has altered the abundance of economically important species and whether sustainable harvesting techniques can enhance the health of populations of these taxa.
We address these questions in the context of a key economic resource from Amazonian peatlands: the fruit of the palm Mauritia flexuosa, which is known in Peru as ‘aguaje’. This palm is the most common species of tree in the forests on the extensive lowland tropical peatlands in northeastern Peru that store at least 3.14 PgC, equivalent to >60 years of national fossil fuel emissions1,9. Mauritia flexuosa is characterized by many features that support successful large-scale NTFP use and commercialization10,11: the palm is a native and abundant component of Amazonian peatland ecosystems12,13, there is a long-standing regional market for the product11 and there are emerging, although far from comprehensively applied, legal mechanisms for communities to obtain land tenure along the major rivers5 and rights to extract forest resources via government institutions. The resource is also already a important component of community incomes and activities: where currently harvested, sale of its fruit represents 15–22% of family incomes (US$208–227 yr−1)14,15. However, fruit harvesting is typically performed by felling the fruiting palms, and as Mauritia flexuosa has a dioecious breeding system (separate male and female individuals), fruit harvesting reduces the density of female palms in these stands16. Although the negative impact of this activity has been recognized for more than 30 years17, estimates of the magnitude of this activity are scarce: estimates of the number of stems of Mauritia flexuosa that are felled each year vary from 12,900–24,000 palms in 200718 to approximately 114,000 between 2012 and 20132. There is also strong spatial and temporal variation in the origin of aguaje fruit arriving in the main city of Iquitos2. However, the overall drivers and cumulative impact of decades of tree felling on the spatial patterns of the proportion of female palms are unknown. It is also unclear whether sites where tree climbing has been introduced as an intervention to promote sustainable management have a higher proportion of female palms than sites where female palms continue to be felled. Evidence for a positive impact of tree climbing and an understanding of the spatial distribution of the resource base would enhance the justification for conservation action to expand the typically patchy and ephemeral sustainable management projects in this region.
Establishing a historical baseline of resource availability
Understanding how historical resource extraction has altered the abundance of species used by local communities requires us to address a key challenge in conservation science: how to quantify the impact of resource extraction on populations when we lack data on species abundance before harvesting. Establishing baselines is crucial for assessing the severity of declines in biodiversity and to stimulate conservation action. For example, long-term monitoring data have been used to estimate that 68% of populations of vertebrate species have declined worldwide since the 1970s19. However, data on the long-term trajectory of plant populations, and tropical species in particular, remain scarce. Anecdotal reports, such as the disappearance of Ceiba trees from Amazonia in the 1970s–1980s20, are susceptible to ‘shifting baseline syndrome’, where successive generations assume that the state of resources that existed when they started to interact with the environment represents the baseline condition21,22. By contrast, qualitative assessments of extinction risk23, and recent quantitative estimates of the impact of land-use change on population sizes of Amazonian tree species24, do not provide sufficiently precise, species-level information on the trajectory of tree populations to design focused interventions. In this article, we tackle the lack of baseline data by measuring, mapping and analysing large-scale spatial variation in a direct indicator of the level of unsustainable extraction of the resource: the proportion of female palms in a stand. The sex ratio for unharvested stands and in plantations is 1:125,26; therefore, by comparing variation in the proportion of female palms within stands to a baseline value of 0.5, we can quantify the levels of resource extraction due to destructive harvesting.
We collected an extensive dataset of this novel indicator to explore the drivers of the intensity of historical resource extraction across the peatland complex of northern Peru. Given its regional importance as a food source, we expected levels of unsustainable extraction to be highest nearest the main urban centre of Iquitos, where transport costs are lowest, similar to other NTFPs with large regional markets27. We also explored the impacts of subsistence consumption by communities on the resource by testing whether smaller palm swamps located near larger communities had a lower proportion of female palms independent of their proximity to Iquitos. Finally, we tested whether the sites where climbing techniques are now used have a higher proportion of female palms compared with sites where palms are cut for fruit harvesting. We use insights from these analyses to model levels of unsustainable resource extraction from M. flexuosa populations across the Amazonian peatland complex and to estimate the decline in fruit production that has occurred as a result of felling palms. By demonstrating both the large cost that historical unsustainable resource extraction imposes on communities today and how sustainable management techniques could improve resource levels at large scales, our results open a route to conserve one of the most carbon-rich landscapes in the Neotropics.
Results
Spatial variation in unsustainable resource extraction
The proportion of female palms of M. flexuosa among stands varied widely among the 93 sites, from 3 sites where less than 6% of adult palms were female to 24 sites with healthy populations where more than 40% of adult individuals were female (Fig. 1). The total travel time to Iquitos from each site was significantly related to the variation in the proportion of female palms (generalized linear mixed model (GLMM); z = 3.9, P < 0.001; Fig. 1 and Supplementary Fig. 1): most of the sites with few female palms are located within four hours travel from Iquitos, whereas four sites located in the communities of Puerto Diaz and Nuevo Milagro, which require >40 hours of travel from Iquitos, have stands with sex ratios approaching 0.5 (Figs. 1 and 2). However, neither the number of inhabitants (GLMM; z = −0.77, not significant (NS)) nor the area of accessible palm swamp for each community (GLMM; z = 0.71, NS) was related to the proportion of female palms.

Sites where trees are cut down to harvest fruits are shown with filled circles, and sites where palms are climbed for harvesting are shown using open circles. The black line indicates the predicted relationship from the mixed model between travel time and the proportion of female palms for sites where trees are cut down to harvest the fruits; the shaded area indicates the 95% confidence limits of this relationship. This relationship was used to predict and map the proportion of female palms across the palm swamps of Loreto.

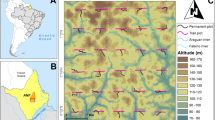
The estimated proportion of female M. flexuosa trees varies more than fourfold across the palm swamps of the region. The solid black line delineates M. flexuosa stands within 5 km of the main rivers and is used for calculating the total area of accessible palm swamps for estimating the maximum potential income from fruit harvesting. Sample sites evaluated for this study are also shown. Inset highlights the location of Iquitos within Loreto, Peru. Country and regional boundaries and rivers downloaded from https://www.geogpsperu.com.
Crucially, sites where palms are climbed, rather than cut, for harvesting have a higher proportion of female palms (GLMM; z = –2.1, P < 0.05; Fig. 1). This effect translates to an increase in the percentage of female palms from 26% to 36% in stands where palms are climbed, compared with cut, to harvest fruits. Variation in the proportion of female palms among sites harvested by climbing may be related to the timing of the transition from cutting to climbing the palms (Fig. 1). For example, in the community of Puerto Alegría, community members initiated tree climbing for harvesting in the 1980s; stands here currently have 52% female adult palms. By contrast, in Pacaya Samiria National Reserve, the transition, overseen by the Peruvian National Parks Authority (SERNANP), was initiated in 2002, and some sites have less than 20% female palms (Supplementary Table 1).
Modelling levels of unsustainable resource extraction of M. flexuosa
The relationship between the proportion of female M. flexuosa palms and the total travel time to Iquitos was used to model variation in this indicator of forest health across the region (Methods). Overall, the predicted level of unsustainable resource extraction declines markedly with distance from Iquitos (Fig. 2), but there are also stands close to the city, such as between the Marañón and Ucayali rivers, that have a high proportion of female palms because they are accessible only by small tributaries rather than via the main rivers (Fig. 3). The Tigre River basin, in the north of the study area, also has M. flexuosa stands with an unexpectedly high proportion of female palms given its proximity to Iquitos (Fig. 2). This finding is consistent with the very low levels of aguaje fruit supply to Iquitos originating from the upper river Tigre in 2012/20132: although there were high levels of fruit supply from close to the mouth of the Tigre from the community of Nueva York, fruit supply diminished much more rapidly with distance along the river, compared with similar distances along the neighbouring river Marañón2,28. The healthy state of the stands alongside the upper river Tigre may be related to the lower frequency of transport compared with other major rivers, as the Tigre River does not connect to other large regional centres or national transport links.

Estimates are calculated for each 1 km2 pixel based on river transport times from Iquitos to the nearest point on a river, plus walking times to the centre of each pixel. River transport is the main method of transportation in Loreto, with boat speed varying according by route and load capacity. Here, we used a speed of 15 km hr−1 for lanchas that ply the main rivers and 4 km hr−1 for peque peques that are used on the tributaries (Methods). Country and regional boundaries and rivers downloaded from https://www.geogpsperu.com.
The total potential production from harvesting fruits of M. flexuosa with the current spatial pattern in the proportion of female palms, estimated for palm swamps located within 5 km of the main rivers, is 298,581 Mg yr−1. This value equates to a gross potential value of US$41 ± 20.1 million yr−1 for the whole region, with per hectare values ranging from US$45 to US$333 ha−1 yr−1 based on current market prices (Fig. 4 and Methods). This large, overall potential value is comparable to the income from tax by oil extraction to the Loreto regional government (Fig. 4). However, historical removal of female palms means that this level of productivity is far lower than might be achieved if the stands of M. flexuosa were in their ‘natural’ state. If 50% of the palms in the stands were female, the estimate of the potential production more than doubles to 829,315 Mg yr−1, with a potential gross value of US$92 ± 38.2 million yr−1 if prices are maintained as the market expands, similar to the gross declared value of timber to the Loreto regional government (Fig. 4). In other words, our estimates suggest that historical human extraction of palm fruits has led to a 55% reduction of the productivity of the resource that is available to communities today.

Left-hand bar shows total potential estimated value for accessible palm swamps <5 km from a main river of (1) M. flexuosa fruit harvesting today (±standard error), (2) M. flexuosa fruit harvesting where sustainable harvesting techniques are employed across the whole area and the proportion of females has increased in absolute terms by 0.1, up to a maximum value of 0.5, consistent with the state of palm swamps today where fruits are harvested by climbing the trees, and (3) under baseline natural conditions assuming a 1:1 sex ratio. Also shown are the average annual value of taxes to the regional government of Loreto for the period 2015–2018 for oil exploitation (centre bar) and the total declared value of timber by forestry companies in Loreto for 2015 (right-hand bar). Timber and oil tax revenue values were adjusted to 2019 values using the World Bank consumer price index67.
More positively, the higher proportion of female palms in sites where tree climbing has been initiated suggests that some of this potential can be recovered over coming decades if this harvesting technique were to be introduced more widely. If the improvement in sex ratio found in the sites where fruit harvesting is carried out by climbing is applied across the region, productivity could rise to 656,384 Mg yr−1, and the potential gross income could rise to US$62 ± 28.2 million yr−1, which is an increase of 51% compared with current values (Fig. 4).
Discussion
Our extensive dataset of the proportion of female palms in stands of M. flexuosa across the peat swamps of northwest Amazonia reveals multiple insights into the drivers and legacy of resource extraction, the impact of sustainable management practices and the economic potential of this resource. The link between levels of resource extraction of stands and their proximity to the main regional market probably reflects the overall importance of the commercial demand for this product as a food source in Iquitos and beyond, rather than subsistence demand within communities, for driving resource extraction. The influence of the market extends approximately two days travel from Iquitos (Fig. 2), consistent with fruits remaining viable for sale up to five days following harvesting14 and the distances of communities that supplied fruit to Iquitos during 2012/20132. Elsewhere in the tropics, shorter distances to the nearest urban centre have also been linked to variation in levels of biodiversity-based resources, such as lower population sizes of mammal, bird and timber species27,29. Our results, therefore, emphasize the importance of managing the impact of cities in tropical forest countries to reduce the impact of resource extraction on surrounding forests while maintaining the services they provide.
Our finding that the total potential production and value of M. flexuosa fruits has halved due to cutting female palms (Fig. 4) provides an estimate of the often unrecognized but substantial cost that current and historical resource extraction places on communities. This impact is typically noted only anecdotally and at small spatial scales. For example, income from fruit extraction through cutting palms in the community of San Miguel diminished by 46% over five years (1995–2000)10 while the community of Dos de Mayo ultimately was forced to stop extraction as the resource became unavailable within the forests surrounding the community (G. Hidalgo, personal communication). By contrast, this study provides a quantitative estimate of the impact of resource extraction across a Neotropical forest landscape that spans nearly three million hectares. Our estimate of loss is similar to the magnitude of global declines in animal populations due to hunting since the 1970s19 and demonstrates that the fingerprint of human resource extraction can also be strong in plant communities even in apparently intact forest landscapes.
The loss of potential production of M. flexuosa fruits reflects the historical management of this species as a common pool resource in these peatland landscapes30,31. Fortunately, recognition that intensively cutting trees for harvesting fruits is unsustainable is leading to the introduction of communal management systems that incorporate several aspects of successful adaptive governance schemes, particularly related to encouraging dialogue among stakeholders and defining the boundaries of the resource32. Sustainable management initiatives that promote tree climbing for fruit harvesting have been led by the communities themselves (such as in Puerto Alegría), non-governmental organizations (for example, Amazonians for Amazonia) and/or government institutions (for example, the Peruvian Forest Service (SERFOR), the Peruvian Fund for Nature, SERNANP and the Loreto regional government (GOREL)). Key components of all these initiatives have been the designation of land tenure and achieving sole legal access for communities over nearby palm swamps and the establishment of community-led associations where a small number of families plan, extract and market the fruit. For communities inside national protected areas, harvesting activities are ultimately regulated by SERNANP, and there are attempts to regulate harvesting activities in communities outside protected areas as part of projects implemented by SERFOR and GOREL. Such regulation involves issuing permits to implement approved management and harvesting activities. However, in all sites, in practice, monitoring and active regulation of harvesting are at an early stage.
Cutting the palms for harvesting preceded the introduction of climbing in all sites, and the timescales of the start of the interventions and the demography of M. flexuosa suggest that the higher proportion of female palms in sites where palms are climbed today is due to recruitment of new adults since the intervention was introduced: M. flexuosa requires at least 6–10 years to become adult from seed10, and these interventions were introduced 10–30 years ago. Of course, full recovery of these forests would require adequate levels of regeneration, and in some sites, seed limitation may prove a barrier to recruitment of new individuals. However, demographic models of M. flexuosa suggest that recruitment can be maintained, up to harvesting rates of 50–90% of fruit racemes33.
On a per hectare basis, according to the current pattern of the proportion of female palms across the region, our analysis indicates a current potential annual gross income of US$45–333 ha−1 yr−1, which is similar to published values for sustainably harvested stands in the community of Veinte de Enero10,34. These values are, of course, lower than equivalent per hectare values of large-scale plantations of Elaeis guineensis for palm oil, which can rise to several thousand dollars per hectare35. However, sustainable harvesting of M. flexuosa fruits provides a wider and more evenly distributed range of benefits than plantation development15,17: it could provide a foundation for a regional economy that substantially improves incomes to local communities while also protecting the high carbon stocks and unique biodiversity of these ecosystems. It is also more ecologically appropriate within the dynamic Amazonian peatlands where drainage is not possible. This approach of using community-led, sustainable management of existing stands of a native palm species contrasts strongly with that of deforestation and conversion to intensive agriculture of a non-native palm species, which has been the predominant development pathway for tropical peatlands in Southeast Asia in recent decades36. As the debate opens up on the nature of economic development in Amazonia following the COVID-19 pandemic, it is crucial that the full potential of sustainable, climate-friendly development options such as M. flexuosa fruit extraction are explored.
Our estimate of the economic potential of M. flexuosa fruit harvesting is broadly comparable to revenue levels from other important NTFPs in Amazonia such as açaí fruit (Euterpe sp.), which had a gross value of approximately US$200 million in 2019 in Brazil37, and Brazil nut extraction, which was worth US$75 million, US$25 million and US$19 million for Bolivia, Brazil and Peru, respectively, in 200538. Intriguingly, the current potential value of M. flexuosa fruit is equivalent to 39% of the annual declared value of timber in 2015 (US$106.6 million) and almost double the average annual value (US$21.6 million) of tax revenue from oil extraction from 2015 to 201839,40 for the region of Loreto in northeastern Peru (Fig. 4). Overall, even with uncertainty about how prices would change as markets develop, these comparisons indicate that sustainable fruit harvesting has similar potential for generating economic benefits for communities to existing major Amazonian NTFPs and could provide benefits that are comparable to redistributing tax revenues from oil extraction or deriving income from logging.
Realizing the potential of sustainable M. flexuosa fruit harvesting will be challenging and require heeding lessons about the potentially illusory nature of the high value of NTFPs within intact tropical forests41,42,43. M. flexuosa fruit does broadly meet criteria associated with successful expansion of NTFP use8,43; for example, development of this product is closely aligned with national environmental policy (in this case, related to enhancing sustainable management of tropical peatlands as part of the Nationally Determined Contributions of Peru to the Paris Agreement44), increases in production and income can be closely linked to improving livelihoods given the widespread presence of extensive palm swamps close to communities across the region, and demographic models suggest ecological sustainability is possible in stands with fruit harvesting33. However, expansion of national and international markets will be crucial to drive increases in the overall value of this product and maintain prices as production increases42. In this context, support for using fruit of M. flexuosa as the basis for juices and food supplements by national and international companies (for example, RAFSAC, Aje Group and Inkanatura)11 will be vital.
A range of steps needs to be taken to support communities that engage in palm fruit harvesting. Currently, uptake of climbing as a sustainable management technique has been limited because of a lack of training, equipment and information about the ease of harvesting by climbing. For example, there is often a perception that cutting down the palm is faster than climbing. However, in practice, both cutting and climbing take similar lengths of time—cutting an adult Mauritia palm takes around 20–25 minutes while climbing takes 17–30 minutes12. A particular challenge is to develop effective community-led monitoring schemes to achieve successful, long-term management32, and long-term support must be offered to communities to ensure that sustainable harvesting techniques are maintained even if demand increases. For example, the Maijunas, an indigenous group in the northern Peruvian Amazon, harvested this resource for their consumption using the mainly sustainable method of collecting ripe fruit from the ground, but as demand increased, the use of destructive techniques became common45. Legal land tenure needs to be expanded to all communities with access to palm swamps, and harvesting areas must be demarcated. More-precise maps of the abundance M. flexuosa palms would also support the expansion of the management of this resource: high-resolution spectral data obtained using unmanned aerial vehicles offers one way to map the distribution of palms accessible to communities as M. flexuosa palms are readily distinguished from the surrounding trees46. Finally, safeguards are required to ensure that communities continue to be the primary beneficiaries of fruit harvesting as the market expands and to avoid the benefits and resource being appropriated solely by private interests47. Overall, careful monitoring of the effects of harvesting on both the social and ecological aspects of these systems is required to ensure that there are not negative impacts37 and support the drive to develop a sustainable and equitable biodiversity-based economy across Amazonia48.
Our map of variation in the proportion of female trees of Mauritia flexuosa across the basin is also important for understanding where greenhouse gas emissions from peatland degradation may occur in the future. Although cutting removes female palms and reduces the abundance of M. flexuosa (Supplementary Fig. 2), the current impact on above- and below-ground carbon stocks and greenhouse gas emissions is likely to be small across the gradient of resource extraction that we studied here. Highly degraded sites, where palm swamps become dominated by pioneer trees, are associated with substantial carbon emissions due to higher respiration rates derived from the remaining organic matter49. However, the sites we studied here still have substantially higher densities of Mauritia flexuosa than the 16 stems per hectare that were found in one highly degraded site where important carbon emissions were reported49 (Supplementary Fig. 4). The sites in our study have, therefore, been modified but currently probably not degraded and damaged to an extent that substantially affects their carbon balance. The challenge for the future is to ensure that these sites are not subsequently converted to secondary forest and other land uses, which is a pathway that may follow the removal of female palms and resulting loss of value of the standing forest. In this context, our map of the proportion of females across this region is an early warning signal of where interventions are required to prevent more substantial modification of the landscape and greenhouse gas emissions. The higher proportion of female palms in sites where trees are climbed for harvesting and increase in the potential economic value of stands where this value approaches 0.5 indicate that introducing climbing techniques is a means to increase the value of standing forest and, as a result, help to conserve the carbon-rich peat deposits below these forests.
Our findings that historical resource extraction has halved the potential production of harvesting M. flexuosa fruit and that this can be reversed by tree climbing provides a powerful incentive to introduce and maintain sustainable management practices. This study, therefore, provides strong support for using sustainable harvesting of M. flexuosa fruits as a focus of Amazonian peatland conservation5,50. For local communities, the demonstrable success of using climbing to harvest fruits for maintaining the health of M. flexuosa populations will help to encourage uptake of this technique across the region. For regional and national government institutions and non-governmental organizations, our findings will help to promote coordinated, region-wide initiatives to support the introduction of sustainable management programmes and support efforts to increase the market for M. flexuosa fruits. For the international conservation community, these results demonstrate the viability of sustainable palm swamp management for preserving Amazonian peatlands and the substantial carbon stocks they hold below ground. Finally, the results demonstrate how apparently intact tropical forests have been affected by resource extraction and how a precise understanding of how ecosystems have been modified can be used to define a pathway for conservation action.
Methods
Study area and species description
The study was carried out across the extensive peatlands of the Pastaza-Marañón foreland basin in the region of Loreto in northern Peruvian Amazonia. This basin is a geological depression formed in reaction to the uplift and eastern movement of the Andes and today acts as a collecting point for water from tributaries of the upper Amazon50. This region is characterized by a warm, humid tropical climate with a mean temperature of 26 °C and a daily range of 22–30 °C51,52. The average annual rainfall of the region is around 3,000 mm, and there is no regular dry season52.
Peatlands cover 35,600 km2 of the basin, and palm swamps, the most extensive vegetation type, cover 2.8 million hectares, or 78%, of this area9. These stands are located along the Marañón, Ucayali and Amazon rivers and their tributaries: the Samiria, Tigrillo, Tigre, Itaya and Nanay rivers (Fig. 3). The palm swamps have an average peat depth of 1.7 m (range 0–5.4 m) and in total contain 2.3 PgC (95% confidence limits: 0.27–6.00)9.
The dominant species of these peatlands is the palm tree Mauritia flexuosa13, which is locally called aguaje (‘burití’ in Brazil, ‘canangucho’ in Amazonian Colombia, ‘moriche’ in Venezuela53). Palm swamps are locally named ‘aguajales’ due to the dominance of M. flexuosa in this type of forest54. M. flexuosa is a large native palm tree of the Neotropics, reaching 35–40 m height and 50 cm diameter12,55. The fruit is an elliptical-globular drupe, 3–7 cm long, with a mean weight of 40 to 85 g (ref. 14). Its phenology varies across the region as the peak of the fruit season is linked to variation in the water level of the river56, and as a result there is a continuous supply of fruit to the market2.
This species is dioecious and thus has separate male and female individuals55,57. The proportion of female palms is therefore a good indicator of levels of unsustainable resource extraction in palm swamp ecosystems as female individuals of M. flexuosa bearing fruit are typically harvested by cutting down the palms16,58 (Supplementary Fig. 6). The sex ratio of M. flexuosa in an undisturbed stand was reported as 1:1 (50% male and 50% female palms) in the Colombian Amazon25 and a similar ratio (1.1:1.0) was found in two experimental plantations of this species in Jenaro Herrera in the Peruvian Amazon12,26. For this study, we assess the level of unsustainable resource extraction as the reduction in the proportion of female palms compared with a baseline of 50%. This knowledge of the baseline gender ratio under natural conditions allows us to estimate the impact of historical resource extraction on the health of M. flexuosa populations.
Sampling sites and field methods
Data on the proportion of female palms in different palm swamps were collected from sites across the region that encompass the full gradient of human use, including largely undisturbed forests, abandoned areas where harvesting stopped 3–15 years ago and areas that are actively harvested every fruiting season (Supplementary Table 1). Stands were selected on the basis of covering a wide range of distances to Iquitos and prioritizing data gaps. In all sites, estimates of the proportion of female palms were based on evaluating the gender of adult palms according to the presence of floral organs by carefully observing flowers/fruits either on the palm using binoculars or on the ground next to the corresponding palm stem (Supplementary Fig. 5).
Data on the variation in the proportion of females of M. flexuosa in the largely undisturbed forests in the eastern portion of the basin were obtained from 16 0.5 ha plots established by the Instituto de Investigaciones de la Amazonía Peruana and the Amazon Forest Inventory Network between February 2017 and October 201959. These data were augmented by information from six transects located adjacent to the plots that were sampled between June 2008 and July 2009. These older transects represent 6.5% of total sampling sites, and the values were validated by transects established more recently in the surrounding area: 14 transects between February 2017 and October 2019 and 10 transects in January and February 2020. In each of these transects, the gender of 100–200 adult palms was recorded. Data on variation in the proportion of female palms of M. flexuosa in the undisturbed forests in the western portion of the region in Datem del Marañón province were collected using 16 plots varying from 0.36 to 0.72 ha in size (Supplementary Table 1) in August and September 201860.
Finally, 31 transects were established as part of this study in June and July 2019 to understand how the proportion of female palms varies among areas that have been harvested in the past but since abandoned and in areas that are being harvested today. Sampling sites were located in accessible areas along the rivers, which is the most important area for fruit sold in the main regional market. For each of these transects, the gender was recorded for 200 individuals, and the transects were extended if necessary until the sample included at least 20 female palms. This sample size and design were chosen because of how uncertainty in estimates of the proportion of female palms varies with the level of the unsustainable resource use using simulations based on the binomial distribution (Supplementary Fig. 7). Sampling 200 individuals provided reasonable levels of confidence in estimates of the proportion of female palms across three levels of harvesting: the baseline proportion (50%), half the natural female proportion (25%) and where there is a low proportion of female palms (10%). Semi-structured interviews with harvesters during fieldwork were used to obtain information about harvesting techniques, the time and distances to harvesting sites and the sale price of a sack of aguaje fruit in each community.
Explanatory variables
Variation in the proportion of female palms associated with different communities was broadly related to variation in the amount of aguaje fruit that each community supplied to Iquitos in 2012/201328 (Spearman rank correlation, P < 0.001; Supplementary Fig. 3). To identify the ultimate causes of spatial variation in unsustainable resource use of palm swamp forests, we analysed whether a range of variables associated with drivers of the intensity of harvesting activities were associated with variation in the proportion of female palms. We assessed the influence of the urban market by comparing the travel time to the main market (Iquitos) from each sampling site with the proportion of female palms. In Iquitos, the fruits are commercialized as fruits and are transformed into a paste for use in other products. The total travel time (hours) from each sampling site to Iquitos was estimated as the sum of the travel time from Iquitos to the nearest community by river, the travel time from the community by river to the nearest access point for each site and the walking time from the river to the site. The estimate of the travel time from Iquitos to each community was based on a boat speed of 15 km hr−1 (by ‘lancha’)61 for routes via the Marañón, Ucayali and Amazon rivers and 4 km hr−1 (by ‘peque peque’) for boats travelling during the fieldwork on the Nanay, Itaya, Tigre, Tigrillo and Samiria rivers. Each palm swamp was allocated to its nearest community, and the travel time from the community to the nearest access point was estimated on the basis of the distance by river and the travel speed by peque peque. Finally, the walking time from the nearest access point to the sampling site was estimated on the basis of the straight-line distance and a walking speed of 1.5 km hr−1 within the forest recorded during fieldwork in palm swamps in this study; the speeds values were registered by a GPS device. Spatial modelling was performed using the cost distance, Euclidean allocation and Euclidean distance functions in ArcGIS v.10.4.162. The modelled travel times closely matched times to reach Iquitos estimated by community members during fieldwork (Supplementary Fig. 8).
The importance of variation in the intensity of harvesting by local communities for determining levels of unsustainable resource use was assessed by exploring whether variation in the number of inhabitants among communities (population size varies from 10 to 3,500 members63) and the area of palm swamp within 5 km of each community explains variation in the proportion of females among sites. The effect of management technique was assessed by comparing the difference in the proportion of female palms among palm swamps where communities cut the palms to harvest the fruits with those where communities climb the palms without damaging the trees12. Information on the technique that has predominately been used at each site during the previous decade was obtained by a semi-structured interview with harvesters during the fieldwork and verified during the census of the transects by noting the presence/absence of cut trunks.
Data analysis
The proportion of females for each sampling site was obtained by dividing the number of female palms by the total number of adult trees. We used GLMMs implemented using the glmer function within the lme4 package64 in R v.3.5.365 to assess whether the explanatory variables were significantly related to the variation in the proportion of female palms. Our statistical model was:
where y is the proportion of female palms within the stand, β0–β4 represent the model coefficients, Time, Inhabitants, Area and Technique are the travel time to the urban market of Iquitos, the number of inhabitants per community, the area of palm swamp within 5 km of each community and the harvesting technique, respectively. Community and River are nested, random factors that reflect that multiple palm swamps may be accessed by any given community, and multiple communities are found along each river. River was included as a random factor as each watershed has different characteristics in terms of river size and transport frequency that may influence how the other factors relate to the proportion of female trees (Supplementary Fig. 1). All continuous variables were scaled before analysis, and each data point was weighted by number of palms sampled at each site.
The analysis was subjected to k-fold, cross-fold validation to test whether the predictions of the mixed model were robust to omitting subsets of the underlying data (Supplementary Fig. 9). Spatial autocorrelation was explicitly incorporated in the original statistical model through the nested, random factors of community and river basin, so we used a random cross-fold approach to validate the predictions with the same underlying model structure. Predicted values of the proportion of female palms for each site, based on running a mixed model with 80% of the original data, were significantly correlated with the observed values (Supplementary Fig. 9). This result demonstrates that the relationships are not unduly influenced by a small subset of the data and the model can be used to describe variation in the proportion of female palms across the range of the explanatory variables that are found in this region.
Modelling variation in the proportion of female palms of M. flexuosa
The finding that travel time explains a substantial amount of variation in the proportion of female palms of M. flexuosa among sites was used to model this parameter across the region of Loreto (Fig. 2). Travel time to Iquitos was estimated across Loreto at a 1 km2 pixel size (Fig. 3), following the preceding methods. All pixels representing M. flexuosa stands were then extracted using a map of the distribution of palm swamps across the Pastaza-Marañón basin9. The coefficients from the mixed model (Fig. 1 and Supplementary Fig. 1) were used to estimate the proportion of female palms of M. flexuosa across the palm swamps of Loreto on the basis of the travel time to Iquitos from each pixel. The proportion of female palms was estimated independently for palm swamps in each river basin as the proportion of female palms varies among watersheds (Supplementary Fig. 1).
Estimating the potential value of M. flexuosa to local communities
The potential area of fruit harvesting was defined as all palm swamps within 5 km from a main river on the basis of information from harvesters obtained during fieldwork. This area was delimited according to the information recorded about the distance they walk to the extraction zones and the distance of the sampling sites from the communities. Palm swamps within this area cover 331,900 ha (Fig. 2; area indicated by the black solid line).
The proportion of females across this harvesting area was used to estimate the gross potential income of fruit sale. First, the density of female palms was estimated on the basis of the estimated proportion of females of each pixel. Our field data demonstrate that the density of males remains constant as the proportion of females declines in the sampled stands; within our dataset, there is no evidence of substantial felling of males even where the females have been almost entirely harvested (Supplementary Fig. 2), even though in some communities, a few male palms are occasionally felled for making walkways or cultivating the palm weevil (Rhynchophorus palmarum3,4). We therefore estimated the density of females as:
where Dfem,i is the estimated number of adult female palms of M. flexuosa per hectare within each pixel, i, in the harvesting area, Fi is the estimated proportion of females of each pixel and Dmale is the average density of male palms across all sites (86 individuals ha–1). This model of how female harvesting affects the density of populations of M. flexuosa fits the observed data closely (Supplementary Fig. 2).
The total potential current income was then estimated as:
where Dfem,i is the density of female palms in each pixel, i, in the harvesting area, and A is the area in hectares per pixel (100 ha). R is the average number of racemes harvested per palm (1.3 ± 0.03 racemes); this value was calculated from data on the number of racemes of 3,567 female palms recorded during fieldwork across all sites and integrates across the temporal and spatial variation in fruit production among individual palms. S is the factor (0.6 ± 0.1 sack/raceme) used to convert the number of racemes into the number of sacks of fruit of aguaje; this value was calculated from measurements of fruit production from 16 M. flexuosa palms in Jenaro Herrera in 201812. P is the average price of US$5 ± 0.32 (17 PEN) per sack recorded in the fieldwork from 13 communities during the interviews with harvesters. The overall projected income for the total harvesting area, Incomet, was calculated as the sum of the value across all pixels and represents a potential, maximum gross value of this resource in this region.
The potential income from harvesting area was also estimated for natural forests, assuming a uniform proportion of 50% female palms across all pixels. Finally, to estimate the potential income if the fruits were harvested by climbing and the forests began to recover, the estimates of the proportion of female palms across all pixels from the current scenario were modified by adding the estimated impact of climbing from the mixed model (an absolute increase of 0.1 in the proportion of female palms up to a maximum value of 0.5). This scenario represents the potential value of the stand after approximately one to two decades if cutting trees down for harvesting ceases today.
The density of palms, number of racemes per palm, the sack/racemes factor and price of sacks all vary due to natural variation among stands and variation in market conditions. To estimate the standard error of the overall potential income, the standard errors of each component were propagated conservatively as the sum of the fractional errors of each term66 as:
Data availability
The datasets used in the analysis in the current study are available within the article and Supplementary Information.
References
Dargie, G. C. et al. Age, extent and carbon storage of the central Congo Basin peatland complex. Nature 542, 86–90 (2017).
Horn, C. M., Vargas Paredes, V. H., Gilmore, M. P. & Endress, B. A. Spatio-temporal patterns of Mauritia flexuosa fruit extraction in the Peruvian Amazon: implications for conservation and sustainability. Appl. Geogr. 97, 98–108 (2018).
Virapongse, A., Endress, B. A., Gilmore, M. P., Horn, C. & Romulo, C. Ecology, livelihoods, and management of the Mauritia flexuosa palm in South America. Glob. Ecol. Conserv. 10, 70–92 (2017).
van der Hoek, Y., Solas, S. Á. & Peñuela, M. C. The palm Mauritia flexuosa, a keystone plant resource on multiple fronts. Biodivers. Conserv. 28, 539–551 (2019).
Roucoux, K. H. et al. Threats to intact tropical peatlands and opportunities for their conservation. Conserv. Biol. 31, 1283–1292 (2017).
Dargie, G. C. et al. Congo Basin peatlands: threats and conservation priorities. Mitig. Adapt. Strateg. Glob. Change 24, 669–686 (2019).
Pandey, A. K., Tripathi, Y. C. & Kumar, A. Non timber forest products (NTFPs) for sustained livelihood: challenges and strategies. Res. J. For. 10, 1–7 (2016).
Kor, L., Homewood, K., Dawson, T. P. & Diazgranados, M. Sustainability of wild plant use in the Andean Community of South America. Ambio 50, 1681–1697 (2021).
Draper, F. C. et al. The distribution and amount of carbon in the largest peatland complex in Amazonia. Environ. Res. Lett. 9, 124017 (2014).
Freitas, L. Impacto del aprovechamiento en la estructura, producción y valor de uso del aguaje en la Amazonía peruana. Recur. Naturales y Ambient. 67, 35–45 (2012).
Aprovechamiento de los Residuos de Mauritia flexuosa (ITP-CITE, 2018).
Falen, L. Y. & Honorio Coronado, E. N. Assessment of the techniques use to harvest buriti fruits (Mauritia flexuosa L.f.) in the district of Jenaro Herrera, Loreto, Peru. Folia Amazónica 27, 131–150 (2018).
Draper, F. C. et al. Peatland forests are the least diverse tree communities documented in Amazonia, but contribute to high regional beta-diversity. Ecography 41, 1256–1269 (2018).
Bejarano, P. & Piana, R. Plan de Manejo de los Aguajales Aledaños al Caño Parinari (WWF-AIF/DK - Reserva Nacional Pacaya Samiria, 2002).
Manzi, M. & Coomes, O. T. Managing Amazonian palms for community use: a case of aguaje palm (Mauritia flexuosa) in Peru. For. Ecol. Manage. 257, 510–517 (2009).
Baker, T. R. et al. How can ecologists help realise the potential of payments for carbon in tropical forest countries? J. Appl. Ecol. 47, 1159–1165 (2010).
Padoch, C. Marketing of non-timber forest products in Western Amazonia: general observations and research priorities. Adv. Econ. Bot. 9, 43–50 (1192).
Delgado, C., Couturierb, G. & Mejía, K. Mauritia flexuosa (Arecaceae: Calamoideae), an Amazonian palm with cultivation purposes in Peru. Fruits 62, 157–169 (2007).
Living Planet Index 2020—Bending the Curve of Biodiversity Loss (WWF, 2020).
Gentry, A. H. & Vasquez, R. Where have all the ceibas gone? A case history of mismanagement of a tropical forest resource. For. Ecol. Manage. 23, 73–76 (1988).
Pauly, D. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 10, 430 (1995).
Soga, M. & Gaston, K. J. Shifting baseline syndrome: causes, consequences, and implications. Front. Ecol. 16, 222–230 (2018).
Nic Lughadha, E. et al. Extinction risk and threats to plants and fungi. Plants People Planet 2, 389–408 (2020).
Ter Steege, H. et al. Estimating the global conservation status of more than 15,000 Amazonian tree species. Sci. Adv. 1, e1500936 (2015).
Khan, F. & de Granville, J. J. Palms in Forest Ecosystems of Amazonia (Springer-Verlag, 1992).
Freitas, L., Zárate, Z., Bardales, R. & Del Castillo, D. Efecto de la densidad de siembra en el desarrollo vegetativo del aguaje (Mauritia flexuosa L.f.) en plantaciones forestales. Rev. Peru. de. Biol. 26, 227–234 (2019).
Benítez-López, A. et al. The impact of hunting on tropical mammal and bird populations. Science 356, 180–183 (2017).
Endress, B. A., Gilmore, M. P., Vargas, V. H. & Horn, C. Data on spatio-temporal patterns of wild fruit harvest from the economically important palm Mauritia flexuosa in the Peruvian Amazon. Data Brief 20, 132–139 (2018).
Ahrends, A. et al. Predictable waves of sequential forest degradation and biodiversity loss spreading from an African city. Proc. Natl Acad. Sci. USA 107, 14556–14561 (2010).
Hardin, G. The tragedy of the commons. Science 162, 1243–1248 (1968).
Ostrom, E. in The New Palgrave Dictionary of Economics Online (eds Durlauf, N.S. & Blume, L.E.) (Palgrave Macmillan, 2008); https://hdl.handle.net/10535/5887
Dietz, T., Ostrom, E. & Stern, P. C. The struggle to govern the commons. Science 302, 1907–1912 (2003).
Isaza, C., Bernal, R., Galeano, G. & Martorell, C. Demography of Euterpe precatoria and Mauritia flexuosa in the Amazon: application of integral projection models for their harvest. Biotropica 49, 653–664 (2017).
Chuquinbalqui, C. M. et al. Diagnóstico socioeconómico de la población organizada para el manejo de recursos naturales en las cuencas Yanayacu Pucate y Pacaya en la Reserva Nacional Pacaya Samiria (Reserva Nacional Pacaya Samiria – SERNANP, 2014).
Koh, L. & Wilcove, D. Cashing in palm oil for conservation. Nature 448, 993–994 (2007).
Murdiyarso, D., Suryadiputra, I. N. & Wahyunto. Tropical peatlands management and climate change: a case study in Sumatra, Indonesia. In Proc. 12th International Peat Congress on Wise Use of Peatlands Vol. 1 (ed. Paivanen, J.) 698–706 (International Peat Society, 2004).
Freitas, M. A. B. et al. Intensification of açaí palm management largely impoverishes tree assemblages in the Amazon estuarine forest. Biol. Conserv. 261, 109251 (2021).
Plan Operativo de Castaña Región Madre de Dios (MINCETUR, 2007).
La Industria de la Madera en el Perú. Identificación de las Barreras y Oportunidades para el Comercio Interno de Productos Responsables de Madera, Provenientes de Fuentes Sostenibles y Legales en las MIPYMES del Perú (FAO, 2018).
Transferencias por Tipo de Canon, Regalías, y Otros (Congreso Perú, 2019).
Peters, C. M., Gentry, A. H. & Mendelsohn, R. O. Valuation of an Amazonian rainforest. Nature 339, 655–656 (1989).
Sheil, D. & Wunder, S. The value of tropical forest to local communities: complications, caveats, and cautions. Conserv. Ecol. 6, 9 (2002).
Belcher, B. & Schreckenberg, K. Commercialisation of non-timber forest products: a reality check. Dev. Policy Rev. 25, 355–377 (2007).
López, M. et al. What Do We Know about Peruvian Peatlands? (CIFOR, 2020).
Gilmore, M. P., Endress, B. A. & Horn, C. M. The socio-cultural importance of Mauritia flexuosa palm swamps (aguajales) and implications for multi-use management in two Maijuna communities of the Peruvian Amazon. J. Ethnobiol. Ethnomed. 9, 29 (2013).
Tagle Casapia, X. et al. Identifying and quantifying the abundance of economically important palms in tropical moist forest using UAV imagery. Remote Sens 12, 9 (2020).
Bruenig, E. F. Conservation and Management of Tropical Rainforests: An integrated Approach to Sustainability 2nd edn (CABI, 2016).
de Mello, N. G., Gulinckb, H., Van den Broeckc, P. & Parra, P. Social-ecological sustainability of non-timber forest products: a review and theoretical considerations for future research. For. Policy Econ. 112, 102109 (2020).
van Lent, J. Land-Use Change and Greenhouse Gas Emissions in the Tropics: Forest Degradation on Peat Soils. PhD thesis, Wageningen Univ. Res. (2020).
Baker, T. R. et al. in Peru: Deforestation in Times of Climate Change (ed. Chirif, A.) 155–174 (IWGIA, Servindi, ONAMIAP & COHARYIMA, 2019).
Bhomia, R. K. et al. Impacts of Mauritia flexuosa degradation on the carbon stocks of freshwater peatlands in the Pastaza-Marañón river basin of the Peruvian Amazon. Mitig. Adapt Strateg. Glob. Change 24, 645–668 (2019).
Marengo, J. in Geoecología y Desarrollo Amazónico: Estudio Integrado en la Zona de Iquitos Biológica – Geographica – Geológica (eds Kalliola, R. & Flores, S.) 35–57 (Univ. Turku Press, 1998).
Koolen, H. H. F., Da Silva, F. M. A., Da Silva, V. S. V., Paz, W. H. P. & Bataglion, G. A. in Exotic Fruits (eds Rodrigues, S. et al.) 61–67 (Elsevier, 2018).
Malleux, O. J. Inventarios Forestales en Bosques Tropicales (Universidad Nacional Agraria La Molina, 1982).
Del Castillo, D., Otárola, E. & Freitas, L. Aguaje, La Maravillosa Palmera de la Vida (Instituto de Investigaciones de la Amazonía Peruana, 2006).
Khorsand Rosa, M., Barbosa, R. & Koptur, S. Which factors explain reproductive output of Mauritia flexuosa (Arecaceae) in forest and savanna habitats of northern Amazonia? Int. J. Plant Sci. 175, 307–318 (2014).
Quinteros, Y., Roca, F. & Quinteros, V. in XIV. Morichales y cananguchales y otros palmares inundables de Suramérica. Parte II: Colombia, Venezuela, Brasil, Perú, Bolivia, Paraguay, Uruguay y Argentina Vol. XIV Serie recursos hidrobiológicos y pesqueros continentales de Colombia (eds Lasso, C. A. et al.) 265–282 (Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, 2016).
Hergoualc’h, K., Gutiérrez-Vélez, V. H., Menton, M. & Verchot, L. V. Characterizing degradation of palm swamp peatlands from space and on the ground: an exploratory study in the Peruvian Amazon. For. Ecol. Manage. 393, 63–73 (2017).
Honorio Coronado, E. N. et al. Intensive field sampling increases the known extent of carbon-rich Amazonian peatland pole forests. Environ. Res. Lett. 16, 074048 (2021).
de Jong, J. The Impact of Indigenous and Local Communities in the Peruvian Amazon: Integrating Forest Inventory and Remote Sensing. MSc thesis, Wageningen Univ. Res. (2019).
Alvarado, L. Estudio del Potencial de las Embarcaciones Solares en la Amazonía. Caso de Estudio Río Napo. MA thesis, Universidad Politécnica Madrid (2017).
ArcGIS Desktop v.10.4 (ESRI, 2015).
Directorio Nacional de Centrol Poblados - Censos Nacionales 2017- XII de Poblacion, VII de vivienda y III de Comunidades indigenas (Instituto Nacional de Estadítica e Informática, 2018).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. https://doi.org/10.18637/jss.v067.i01 (2015).
R Core Team. R: A Language and Environment for Statistical Computing. R version 3.5.3 (R Foundation for Statistical Computing, 2019).
Taylor, J. R. An Introduction to Error Analysis: The Study of Uncertainties in Physical Measurements 2nd edn (University Science Books, 1997).
Consumer Price Index (Peru) (World Bank Group, 2020); https://data.worldbank.org/indicator/FP.CPI.TOTL?locations=PE
Acknowledgements
We thank the Gordon and Betty Moore Foundation (grant number 5349; ‘MonANPeru: Monitoring Protected Areas in Peru to Increase Forest Resilience to Climate Change’ to T.R.B. and O.L.P.) for funding a Masters by Research scholarship to C.G.H.P. at the University of Leeds and subsequent time for manuscript preparation, fieldwork by C.G.H.P. for this study in 2019 and establishment and re-measurement of forest plots in the palm swamps of Loreto during 2017–2019. Data collection was also funded by the BOSQUES research group of IIAP (led by D.d.C.T.), the projects ‘Protecting biodiversity and livelihoods in the wetlands of Peruvian Amazonia’ (agreement no. 220-2018-FONDECYT, to J.d.A.-P., E.N.H.C. and K.H.R.), which was funded through the Newton-Paulet Fund by the Department for Business, Energy and Industrial Strategy (UK) and CONCYTEC (Peru) and delivered by the British Council, and ‘ARBOLES: A trait-based understanding of LATAM forest biodiversity and resilience’ (agreement no. 001-2019-FONDECYT; to E.N.H.C.), which was supported by CONCYTEC, the Newton Fund, the Embajada Británica Lima and NERC. Data analysis and writing was also supported by the project ‘Novel approaches to understand the state of biodiversity and support livelihoods: the distribution and degradation levels of Mauritia flexuosa stands in Amazonia’ (grant agreement 41469429; to T.R.B. and D.d.C.T.), which was funded through the Newton-Paulet Fund by the Department for Business, Energy and Industrial Strategy (UK) and CONCYTEC (Peru) and delivered by the British Council. E.N.H.C. also acknowledges support from a NERC Knowledge Exchange Fellowship (NE/V018760/1). We thank Amazónicos por la Amazonía (AMPA), Pacaya Samiria and Allpahuayo Mishana National Reserves managed by the Peruvian Protected Areas Authority (SERNANP) and the many communities that hosted fieldwork as part of this study for their assistance and permission to conduct this study. We thank D. Williams and T. Milleron for discussions about earlier drafts of this manuscript.
Author information
Authors and Affiliations
Contributions
C.G.H.P., E.N.H.C. and T.R.B. conceived the study; C.G.H.P., T.R.B., O.L.P. and E.N.H.C. designed the study. C.G.H.P. and T.R.B. analysed the data; C.G.H.P. created the maps and wrote the paper. J.d.A.-P., J.D.J., J.M.R.H., C.J.C.O., G.G.M., G.F.L., E.R.P., J.I.P., L.G.S. and M.M.B. led or took part in the field data collection, and T.R.B., O.L.P., D.d.C.T., E.N.H.C., K.H.R. and J.d.A.-P. contributed funding. S.J.C., F.C.D., S.d.B., M.P.-C., M.v.d.Z., G.M. and J.L. provided materials for data analysis and mapping. All co-authors commented on and approved the manuscript.
Corresponding author
Ethics declarations
Consent for publication
The authors affirm that human research participants provided informed consent for publication of the images in Supplementary Fig. 6.
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Sustainability thanks Verina Ingram and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Table 1 and Figs. 1–9.
Rights and permissions
About this article

Cite this article
Hidalgo Pizango, C.G., Honorio Coronado, E.N., del Águila-Pasquel, J. et al. Sustainable palm fruit harvesting as a pathway to conserve Amazon peatland forests. Nat Sustain 5, 479–487 (2022). https://doi.org/10.1038/s41893-022-00858-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41893-022-00858-z
This article is cited by
-
Anthropogenic threats and habitat degradation challenge the conservation of palm genetic resources—an appraisal of current status, threats and look-ahead strategies
Biodiversity and Conservation (2023)
-
Major carbon losses from degradation of Mauritia flexuosa peat swamp forests in western Amazonia
Biogeochemistry (2023)
