
Abstract
Pastoralism in Old World savannas is known to emit a significant share of global livestock-sourced greenhouse gases (GHG). Here, we compare calculated emissions from animals in a wildlife-dominated savanna (14.3 Mg km−2), to those in an adjacent land with similar ecological characteristics but under pastoralism (12.8 Mg km−2). The similar estimates for both, wildlife and pastoralism (76.2 vs 76.5 Mg CO2-eq km−2), point out an intrinsic association of emissions with herbivore ecological niches. Considering natural baseline or natural background emissions in grazing systems has important implications in the analysis of global food systems.
Similar content being viewed by others

Introduction
Livestock-mediated greenhouse gas (GHG) emissions are considered a large causative agent of climate change, with up to 3.75 Gt CO2-eq emitted yearly1. A large part of them is attributed to low-input livestock systems in regions with vast savanna landscapes, with Sub-Saharan Africa and South Asia specifically accounting for 0.68 (18% of livestock GHG) and 0.73 (20% of livestock GHG) Gt CO2-eq, respectively1.
Natural rangelands, including savannas, store large amounts of soil carbon and have the capacity to act as carbon sinks, especially when restored from degraded states. However, the current emissions attributed to livestock are considered to exceed the mitigation capacity of rangeland soils2. The bulk of direct GHG emissions attributed to livestock consists of methane (CH4) produced during the enteric fermentation in the rumen and CH4 and nitrous oxide (N2O) released by manure, so that grazing livestock is considered an intense GHG emitter3. Following the identification of CH4 as a good target for quick and effective climate action, more than 100 countries agreed to drastically reduce emissions at the recent UN Climate Change Conference (COP26) in Glasgow4. Such policy guidelines inevitably lead to recommending the abandonment of livestock grazing in vast rangeland areas, which would be subjected to rewilding5, both for the assumed potential of reduction in CH4 emissions and for the potential sequestration of carbon once herbivory is drastically reduced6. These calls are not exclusive to high-income countries but extend to countries with very significant livestock populations, most notably India—the largest livestock CH4 emitter worldwide1—where the abandonment of traditional animal husbandry is proposed7.
Grazing livestock and wild herbivores occupy similar ecological niches, raising the question whether abandonment of pastoralism would necessarily lead to a significant reduction of emissions8. Large herbivores attain biomass densities in wild or rewilded landscapes that are particularly high in Sub-Saharan Africa and, to a lesser extent, South and South-East Asia9. There, unlike in other areas, guilds of large herbivores have been preserved up to the present. Recolonization of abandoned rangelands by large herbivores is thus likely to reach very high densities10, and such wild fauna can potentially attain high levels of GHG emissions8. In order to check for the relevance of this process of herbivore substitution, the hypothesis needs to be tested with current field data comparing analog ecosystems grazed by livestock and wildlife.
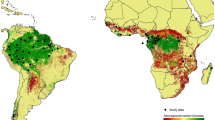
In this study, we aimed to analyze the scenario of a tropical savanna landscape where grazing livestock has dominated for many decades11, but which has the potential to be rapidly recolonized by wild herbivores in the case of abandonment. To examine the contribution to global warming from animals in wildlife-dominated vs. livestock-dominated landscapes, we selected two adjacent areas in Northern Tanzania (Fig. 1) for which enough data are available to establish a comparison. We calculated the GHG emissions per unit area in the Serengeti-Mara ecosystem, a large, 25,000 km2 wilderness area dominated by migratory wildebeest as grazers and elephants as browsers. We compared them with the adjacent Loliondo Game Controlled Area (GCA), an agricultural land essentially belonging to the same savanna ecosystem but being almost exclusively used by Maasai pastoralists.

The Serengeti ecosystem is representative of an African savanna dominated by wild herbivores, while the Loliondo GCA is a similar ecosystem but where pastoralist livestock dominates herbivory processes. Illustrations of the dominant herbivores reproduced, with permission, from Manzano & White8. Note that termites are not considered in the present study and should, in principle, be equally abundant in both systems (see Supplementary Methods).
Currently, there are still considerable limitations to estimate GHGs from animals in free-range conditions, especially with regard to their enteric CH4 output. In an attempt to capture the variability associated with the calculation method, we applied five different approaches to estimate GHGs from herbivore animals (Supplementary Table 1), based mainly on the recent IPCC guidelines12, refs. 13 and 14.
On average from the five different calculation approaches (Fig. 2), the wildlife area (Serengeti-Mara ecosystem) showed similar GHG emissions (76.2 ± 14.7 Mg CO2-eq km−2 year−1) compared to pastoralism (76.5 ± 15.3 Mg CO2-eq km−2 year−1). Animal sourced GHG emissions from both landscapes were also in a very similar range when compared under the same estimation method (Fig. 3), and consistently dominated by enteric CH4 from ruminant digestion, which implies between 84 and 91% of their GHG emissions. Differences among the two studied areas were particularly low (<2%), when estimated through the allometric methods, which highlights the importance of the calculation approach in the results. Indeed, the greatest discrepancies among the two studied areas (5–6%) were obtained when applying the three methods based on IPCC guidelines. These guidelines are specifically designed to help countries reporting GHG emissions from domestic animals, and inherently imply limitations for calculating enteric CH4 production from wild herbivores. Based on the animal’s energy requirements and feed conditions, IPCC Tier 2 is considered the most accurate method for computing CH4 output from livestock ruminants12. However, when applied to wild herbivores, calculation involves important assumptions and simplifications related to feed quality and digestive system (e.g.,: Tier 2 method is especially designed for ruminants, no hindgut or foregut non-ruminant fermenters13) possibly leading to deviations of GHG emissions from Serengeti-Mara ecosystem, with a higher presence of hindgut fermenters (Supplementary Fig. 1).

Points and error bars indicate the average and standard deviation of GHG emissions computed by applying five different estimation methods13,14 (IPCC Tiers 1, 2a, and 2b, as well as allometric 1 and 2). GHG calculations include CH4 from enteric fermentation, and CH4 as well as N2O from manure deposition in pasture. Stacked bars indicate the biomass density of large herbivores present in the area. The orange box indicates the estimated range (first and third quartiles) of herbivore biomass density in Africa across medium-productivity ecosystems9.

Greenhouse gas emissions are expressed in kg CO2eq km–2 year–1. The Serengeti ecosystem is dominated by wild herbivores, while the Loliondo Game Controlled Area is dominated by pastoralism. The figure shows the results of five different calculation methods13,14 applied (IPCC Tiers 1, 2a and 2b, as well as allometric 1 and 2).
The GHG figures are coherent with the similar herbivore biomass densities observed in both systems (Fig. 2 and Supplementary Tables 2 and 3), although these data are also subject to uncertainty from census estimates and body mass assumptions. Grazing animals in the Loliondo GCA are mostly ruminants (12.1 Mg km−2) composed by herds of cattle (Bos taurus×indicus), sheep (Ovis aries) and goats (Capra hircus). The Serengeti-Mara ecosystem, which shows a greater diversity of grazer animals than adjacent pastoral land, is also dominated by ruminants (11.8 Mg km−2), in particular blue wildebeest (Connochaetes taurinus) and buffalo (Syncerus caffer), but a considerable population of hindgut herbivores (e.g., zebras: Equus quagga) is also found (2.4 Mg km−2). A greater diversity of wild grazing animals enables a higher carrying capacity and therefore a greater biomass density of herbivores, which nevertheless emits GHGs in a similar magnitude than equivalent areas under pastoralism due to the presence of hindgut grazers. No differences in land degradation or drastic changes in plant communities are to be expected in the long run, given the long history of pastoralism in the area11, and the ecological similarities of wild migratory systems and mobile pastoralism that also yield high, yet sustainable, herbivore densities10.
Our results show that, in the case of the Serengeti ecosystem, strategies aimed at intensifying and even industrializing livestock production, abandoning grazing lands as a consequence, would not achieve a significant reduction in the total amount of direct GHG emissions from animals. This raises concerns about any related measures in sub-Saharan Africa or in many Asian countries, where a recolonization of abandoned grazing land with rich herbivore guilds and high herbivore densities is more likely9 (orange box in Fig. 2), but also for other continents like Europe, where remarkably high herbivore densities (7.8–12.8 Mg km−2) have been observed in rewilding sites9.
Modeling the climatic consequences of grazing abandonment strategies that promote ecological restoration through rewilding, accounting even for carbon services from wild animals, as recently proposed15, is a difficult challenge. There is insufficient information to come up with reliable natural baselines of wild herbivore populations, and their associated GHG emissions. In light of our results, we believe it is important to increase the efforts to estimate more accurately such ecological baselines, so decisions are well informed enough to be effective against climate change. In short, an effort to promote wildlife conservation needs other arguments than GHG emissions.
The Serengeti ecosystem is possibly the best-studied among the migratory herbivore systems that have been preserved up to the present and its biomass densities are supported by observations in other African sites of similar characteristics9. Rangeland ecosystems in other continents reflect more complex scenarios. Oceania displays extreme examples where rewilded ecosystems should be dominated by mammal non-ruminant foregut fermenters such as kangaroos, who display low emission intensities16, or even by ratite birds17 that would also cause low emissions typical of avian monogastrics. In these cases, however, abandonment scenarios are likely to host introduced ruminating foregut fermenters—in fact, Australia already hosts the world’s largest population of feral camels18. Europe, Central and North Asia, or the Americas, have native ruminants but integrated into impoverished herbivore guilds with lower population densities9. Whether they would experience great increases following livestock abandonment is difficult to predict. Fragmented landscapes are a hindrance to the re-establishment or preservation of large animal migrations19. The ability of wildlife to coexist with rural human populations or with recreational activities is also limited20. In any case, we would expect numbers of wild herbivores in fragmented or relatively humanized settings to be sizeable enough for their GHG emissions to be considered.
Out of practical reasons, at the current guidelines of the IPCC for national GHG inventories, animal GHG emissions on managed lands (e.g., N2O) are ‘assumed to be equal to emissions on unmanaged land. These latter emissions are very low. Therefore, nearly all emissions on managed land are considered anthropogenic’21. However, measured N2O emissions from unmanaged land have been found to be similar to those emissions from crop residues in managed land22. Such N2O emissions represent one of the largest sources of inventoried N2O emissions (e.g., 24% of all soil N2O in 2021 in the USA23).
Overlooking bott CH4 and N2O natural emissions in national GHG accounting is, therefore, expected to lead to an overestimation of the net anthropogenic emissions caused by managed agriculture. Our research points to a significant overlap of managed and natural rangelands, especially when conservation or climatic policies are proposed that suggest massive conversion of a given land from one category into the other6. We can help nuancing current calls to shift into plant-based diets to increase the sustainability of the food system24, because the resulting abandoned land will clearly not be a zero-emission scenario8. Instead, livestock systems that mimic natural migratory herbivore systems as much as possible, such as mobile pastoralism, should be promoted to mitigate climate change at the expense of industrialized high-input production systems10,25. In addition, decisions to promote wildlife conservation or pastoralism cannot rest by putative advantages to GHG emissions but must focus on other reasons or effects. The maximum potential of CH4 and N2O emissions reductions may therefore be overstated26. The inclusion of ecosystem-specific natural baselines in GHG accounting methods could help clarifying the climatic impacts of such changes.
Methods
Livestock and wild herbivores population in studied areas
We used the information provided in Supplementary Tables 2 and 3 describing livestock and wildlife populations in the studied areas. For livestock population in the Loliondo Game Controlled Area (GCA), we used the information provided in Slootweg27 originating from an average year28, assuming an area of 4000 km2 and negligible wildlife presence29 (see Supplementary Methods).
Body mass values for the different animal categories of livestock were taken from Tables 10A.5 of the IPCC 2019 guidelines12. For animal populations in the Serengeti, we used estimates from ref. 30, which considers an area of 25,000 km2 for the Serengeti-Mara Ecosystem. Body mass estimates of wild herbivores were taken from ref. 31.
Greenhouse gas emissions from livestock and wild herbivores
There are still considerable limitations to estimate GHGs from animals in free-range conditions, especially with regards to its enteric CH4 output. In an attempt to capture the variability associated with the calculation method, we applied five different approaches to estimate GHG emissions from herbivore animals (Supplementary Table 1). Hence, GHG emissions estimates were calculated according to the most updated version of IPCC guidelines12 computed with the Tier 1 and two variants of the Tier 2 approach as applied by Smith et al.13. In addition, two allometric methods were used for calculating enteric emissions based on refs. 13 and 14.
Statistical analysis
The statistical analysis was performed using the R software32. Descriptive statistics were used to analyze the main features (mean, standard deviation) of the results from the five estimation methods applied to each scenario (wildlife and pastoralism).
Data availability
The data generated during the current study are available at Figshare (https://doi.org/10.6084/m9.figshare.22141448.v1).
References
Food and Agriculture Organization of the United Nations. FAOSTAT Database https://www.fao.org/faostat/en/#data/GT (2022).
Chang, J. et al. Climate warming from managed grasslands cancels the cooling effect of carbon sinks in sparsely grazed and natural grasslands. Nat. Commun. 12, 118 (2021).
Gerber, P. J. et al. Tackling Climate Change Through Livestock: A Global Assessment of Emissions and Mitigation Opportunities (FAO, Rome, 2013).
United Nations Environment Programme and Climate and Clean Air Coalition. Global Methane Assessment: Benefits and Costs of Mitigating Methane Emissions (UNEP, Nairobi, 2021).
Sun, Z. et al. Dietary change in high-income nations alone can lead to substantial double climate dividend. Nat. Food 3, 29–37 (2022).
Hayek, M. N., Harwatt, H., Ripple, W. J. & Mueller, N. D. The carbon opportunity cost of animal-sourced food production on land. Nat. Sustain. 4, 21–24 (2021).
George, L. S. & Panda, S. Climate Change: A Threat to Global Health Security. Policy Brief for India (The 2021 report of the Lancet Countdown on Health and Climate Change, 2021).
Manzano, P. & White, S. R. Intensifying pastoralism may not reduce greenhouse gas emissions: wildlife-dominated landscape scenarios as a baseline in life-cycle analysis. Clim. Res. 77, 91–97 (2019).
Fløjgaard, C., Pedersen, P. B. M., Sandom, C. J., Svenning, J. C. & Ejrnæs, R. Exploring a natural baseline for large‐herbivore biomass in ecological restoration. J. Appl. Ecol. 59, 18–24 (2022).
Manzano, P., Pardo, G., Itani, M. A. & del Prado, A. Underrated past herbivore densities could lead to misoriented sustainability policies. NPJ Biodivers. 2, 2 (2023).
Estes, A. B., Kuemmerle, T., Kushnir, H. Radeloff, V. C. & Shugart, H. H. Serengeti IV. Sustaining Biodiversity in a Coupled Human-Natural System, Ch. 18 (University of Chicago Press, 2015).
Gavrilova, O. et al. 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories, Ch. 4.10 (IPCC, Geneva, 2019).
Smith, F. A., Lyons, S. K., Wagner, P. J. & Elliott, S. M. The importance of considering animal body mass in IPCC greenhouse inventories and the underappreciated role of wild herbivores. Glob. Chang. Biol. 21, 3880–3888 (2015).
Clauss, M. et al. Comparative methane production in mammalian herbivores. Animal 14, s113–s123 (2020).
Berzaghi, F. et al. Value wild animals’ carbon services to fill the biodiversity financing gap. Nat. Clim. Change 12, 598–601 (2022).
Wilson, G. R. & Edwards, M. J. Native wildlife on rangelands to minimize methane and produce lower‐emission meat: kangaroos versus livestock. Conserv. Lett. 1, 119–128 (2008).
Bond, W. J., Lee, W. G. & Craine, J. M. Plant structural defences against browsing birds: a legacy of New Zealand’s extinct moas. Oikos 104, 500–508 (2004).
Saalfeld, W. K. & Edwards, G. P. Distribution and abundance of the feral camel (Camelus dromedarius) in Australia. Rangel. J. 32, 1–9 (2010).
García-Fernández, A. et al. Herbivore corridors sustain genetic footprint in plant populations: a case for Spanish drove roads. PeerJ 7, e7311 (2019).
Baynham-Herd, Z., Redpath, S., Bunnefeld, N., Molony, T. & Keane, A. Conservation conflicts: behavioural threats, frames, and intervention recommendations. Biol. Conserv. 222, 180–188 (2018).
Eggleston, H. S., Buendia, L., Miwa, K., Ngara, T. & Tanabe K. (eds.) IPCC Guidelines for National Greenhouse Gas Inventories (IGES, Hayama, Japan, 2006).
Kim, D.-G., Giltrap, D. & Hernandez-Ramirez, G. Background nitrous oxide emissions in agricultural and natural lands: a meta-analysis. Plant Soil 373, 17–30 (2013).
Inventory of US Greenhouse Gas emissions and sinks 1990-2019. National Inventory Submissions 2021. United States of America. Common Reporting Format (CRF) Table; https://unfccc.int/sites/default/files/resource/usa-2021-crf-15apr21.zip (2021).
Ripple, W. J. et al. World scientists’ warning of a climate emergency 2021. Bioscience 71, 894–898 (2021).
Pardo, G., Casas, R., del Prado, A., & Manzano, P. Carbon footprint of transhumant sheep farms: accounting for natural baseline emissions in Mediterranean systems. Int. J. Life Cycle Assess. https://doi.org/10.1007/s11367-023-02135-3 (2023).
Staniaszek, Z. et al. The role of future anthropogenic methane emissions in air quality and climate. NPJ Clim. Atmos. Sci. 5, 1–8 (2022).
Slootweg, S. Handbook of Climate Change Resilience, Ch. 82 (Springer, Cham, 2018).
Manzano, P. & Yamat, L. Livestock sector in the Ngorongoro District: Analysis, Shortcomings and Options for Improvement (Ngorongoro District Council and GIZ, 2018).
Hopcraft, J. G. C. et al. Serengeti IV. Sustaining Biodiversity in a Coupled Human-Natural System, Ch. 6 (University of Chicago Press, 2015).
Sinclair, A. R., Packer, C., Mduma, S. A. & Fryxell, J. M. Serengeti III: Human Impacts on Ecosystem Dynamics (University of Chicago Press, 2009).
Sinclair, A. R. E., Mduma, S. & Brashares, J. S. Patterns of predation in a diverse predator–prey system. Nature 425, 288–290 (2003).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria https://www.R-project.org/ (2020).
Acknowledgements
Financial support was provided by the Spanish Government through María de Maeztu excellence accreditation 2023-2026 (Ref. CEX2021-001201-M, funded by MCIN/AEI/10.13039/501100011033); by the Basque Government through the BERC 2022-2024 program; by the CircAgric-GHG project funded by the 2nd 2021 call “Programación conjunta internacional 2021” (MCIN/AEI/10.13039/501100011033) and the European Union NextGenerationEU/PRTR (ref. num: PCI2021-122048-2A); and by the IUBS project “Global Integrative Pastoralism Program”. AdP is funded by the Ramon y Cajal program from the Spanish Ministry of Economy and Competitiveness (RYC-2017-22143). AdP and PM are funded by Ikerbasque—Basque Science Foundation. P.M. was also funded by a post-doctoral grant of the Helsinki Institute of Sustainability Science.
Author information
Authors and Affiliations
Contributions
P.M. contributed conceptualization, methodology, investigation, and writing—original draft. A.d.P. contributed methodology, investigation, funding acquisition, project administration, supervision, and writing—review & editing. G.P. contributed methodology, investigation, visualization, and writing—original draft.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Manzano, P., del Prado, A. & Pardo, G. Comparable GHG emissions from animals in wildlife and livestock-dominated savannas. npj Clim Atmos Sci 6, 27 (2023). https://doi.org/10.1038/s41612-023-00349-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41612-023-00349-8
This article is cited by
-
Wildlife harvests can advance food security and the food systems agenda
Nature Food (2024)
-
Risk to rely on soil carbon sequestration to offset global ruminant emissions
Nature Communications (2023)
-
Herbivory baseline estimates in Spanish protected areas, and environmental implications
Landscape Ecology (2023)
