
Abstract
The Acheulean is the longest cultural tradition ever practised by humans, lasting for over 1.5 million years. Yet, its end has never been accurately dated; only broad 300–150 thousand years ago (Kya) estimates exist. Here we use optimal linear estimation modelling to infer the extinction dates of the Acheulean at global and continental levels. In Africa and the Near East the Acheulean is demonstrated to end between 174 and 166 Kya. In Europe it is inferred to end between 141 and 130 Kya. The Acheulean’s extinction in Asia occurs later (57–53 Kya), while global models vary depending on how archaeological sites are selected (107–29 Kya). These models demonstrate the Acheulean to have remained a distinct cultural tradition long after the inception of Middle Palaeolithic technologies in multiple continental regions. The complexity of this scenario mirrors the increasingly dynamic nature of the Middle Pleistocene hominin fossil record, suggesting contemporaneous hominin populations to have practised distinct stone-tool traditions.
Similar content being viewed by others

Introduction
Acheulean stone tools were produced for more than 1.5 million years. Such an extended period of production is well established, with an age bracket of ~1.75 to 0.15 million years ago (Mya) widely cited as ‘the Acheulean period’ (Gowlett, 2015; de la Torre, 2016; Shea, 2017; Galway-Witham et al., 2019). Discoveries at Konso (Ethiopia), Olduvai Gorge (Tanzania), and West Turkana (Kenya) provide convincing evidence of the Acheulean’s origin in east Africa around 1.75 Mya (Lepre et al., 2011; Beyene et al., 2013; Diez-Martín et al., 2015). Multiple other sites support such an early occurrence in this region (de la Torre and Mora, 2014; de la Torre, 2016; Gallotti and Mussi, 2018), and no other countries claim evidence to the contrary (e.g. Dennell, 2018; Moncel and Ashton, 2018). The location and timing of the onset of the Acheulean therefore appears well supported.
In comparison, the end of the Acheulean is a relative unknown. No sites are widely recognised as displaying evidence of the ‘last Acheulean populations’, and no single region (nor continent) is convincingly argued to display the last stronghold of this technology. Instead, the Acheulean is broadly considered to have been replaced across the Old World between 0.3 and 0.15 Mya, but there is considerable debate on precisely when and where these transitions occurred, and how they varied between different regions (McBrearty and Tryon, 2006; Norton et al., 2009; Fontana et al., 2013; Akhilesh et al., 2018; Méndez-Quintas et al., 2020).
Clarity on when and where the Acheulean ended is hampered by a lack of sites younger than 300 thousand years ago (Kya), limited radiometric dating, the publication of minimum-only dates, geographic imbalances in where artefacts are discovered, and debate concerning ‘transitional’ assemblages. The Korean peninsula, for example, has a series of sites displaying handaxe-like implements dating to <100 Kya (Bae, 2017; Lee, 2017). However, because of their recent age (i.e. under 150 Kya) and a lack of understanding concerning the Acheulean of northeast Asia, we do not know whether these occurrences represent technological convergence, a very late, localised continuation of the Acheulean, or part of a broader maintenance of the tradition across east Asia (Bae, 2017; Lee, 2017). Similar arguments can be made concerning other late Acheulean sites in India, the Arabian Peninsula, Western Europe, and sub-saharan Africa (e.g. Michel et al., 2009; Haslam et al., 2011; Scerri et al., 2018; Méndez-Quintas et al., 2019), although temporal and geographic discrepancies with traditional notions of the late Acheulean are often reduced. Brumm and Rainey (2011) highlight such issues well in their description of bifacial core tools from Northern Australia. In any other region of the Old World these tools could easily have been described as Acheulean handaxes “based on typology alone” (Brumm and Rainey, 2011, p. 57), and yet when found in an Australian context with no Acheulean hominin associations, technological convergence is by far the more plausible explanation.
Yet, understanding when and where the Acheulean ended is important. The technologies characterising the Acheulean, handaxes and cleavers, are unavoidably associated with Homo erectus, H. heidelbergensis, and other Middle Pleistocene hominin species (Corvinus, 2004; Dennell, 2009; Lycett, 2009; Haslam et al., 2011; Herries, 2011; de la Torre and Mora, 2014; Bae, 2017; Galway-Witham et al., 2019; Moncel et al., 2020a). In turn, where one is found, the other is often inferred, and an absence of Acheulean artefacts has recurrently (but not always [e.g. Sanchez-Yustos et al., 2018]) been linked to an absence of these species. Moreover, Acheulean tools have been fundamental to debates on the ‘muddle in the middle’ (Isaac, 1972; Gowlett, 1997; Malinsky-Buller, 2016), and are unavoidably linked to our understanding of hominin cognition, sociality, language, anatomy, and behaviour during this period (e.g. Hopkinson, 2007; Stout, 2011; Uomini and Meyer, 2013; Gowlett, 2015; Key and Lycett, 2018; Wynn and Gowlett, 2018; Pappu and Akhilesh, 2019). The replacement of Acheulean tools by the Middle Stone Age (MSA) and Middle Palaeolithic (MP) also represents a significant behavioural shift, marking the arrival of more complex Levallois and blade technologies often associated with early H. sapiens and Neanderthals (H. neanderthalensis) (Foley and Lahr, 1997; Henshilwood and d’Errico, 2005; McBrearty and Tryon, 2006; Villa, 2009; Fontana et al., 2013; Shipton, 2016; Deino et al., 2018; Galway-Witham et al., 2019; Scerri et al., 2019). Levallois and blade production techniques arrive with regionally dependent variation, and yet no matter when and where they first appear, changes to the cognition, anatomy, diet, and behaviour of hominins are inferred (e.g. Villa, 2009; Shipton, 2016; Picin, 2017; Akhilesh et al., 2018; Pappu and Akhilesh, 2019; Mathias et al., 2020; Moncel et al., 2020a; Meignen and Bar-Yosef, 2020).
Lithic culture is malleable and absorptive (Lycett and von Cramon-Taubadel, 2015; O’Brien and Buchanan, 2017), and we are in no doubt that later Acheulean populations likely combined known technologies with newly learnt or discovered ideas in diverse and chronologically variable ways (e.g. Mathias et al., 2020; Meignen and Bar-Yosef, 2020). However, refining our understanding of when hominin populations moved away from a purely ‘Acheulean’ technological way of thinking is important, and has implications for a multitude of cultural and biological factors in human origins research.
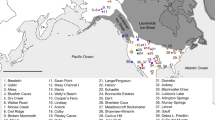
How, then, are we to seek better resolution on the end of the first global technological phenomenon without multiple well-dated sites being discovered across Africa and Eurasia? To start, any attempt to do so should account for imbalances and outliers concerning where late Acheulean sites have been found and dated (Fig. 1). Western Europe, Israel, and South Korea in particular have a relative abundance of sites dating to under <300 Kya compared to other regions and countries. We are not saying there are lots—far from it—but there are more than in other areas, and in turn, our temporal and geographic understanding of the later Acheulean is skewed to reflect this. Second, archaeological sites across 1.5 million years contribute to our view of the Acheulean as a global culture, and yet a paucity of data from the terminal end of this period limits debate to local or regional levels. Thus, there is a clear need to understand the end of the Acheulean as we currently recognise it during its peak; as a global cultural phenomenon displaying elements of unity and diversity, bound by shared functional necessities and the social transmission of tool-design concepts (Lycett and Gowlett, 2008; Wynn and Gowlett, 2018). However, at the moment we do not have the resolution to understand the end of the Acheulean at a localised level with any security, let alone how long it continued as a global or continental tradition.

Some regions have stronger evidence for the presence of Acheulean technology than others. In some instances this may not reflect whether hominins with knowledge of Acheulean technology were present, but instead, reflects political and historical differences between regions and their influence on archaeological discoveries. Alternatively, there may be reduced requirements for Acheulean technologies in some of these regions (e.g. the classic Movius line debate [Lycett and Bae, 2010]). In all instances other than central Saharan regions there is archaeological and/or fossil evidence suggesting hominin presence during portions of the Acheulean period, or strong motivation to infer their presence. Original satellite image credit: NASA Visible Earth Project.
Conservation science provides a route to more reliably define the end of the Acheulean both chronologically and geographically. No other discipline is more concerned with accurately predicting extinction dates based on fragmentary evidence. In the same way that handaxes and cleavers provide sporadic evidence of the presence of Acheulean populations, sightings, spoor and biological samples provide intermittent evidence of the presence of rare species. As species’ populations decrease and go extinct, these traces become more infrequent, geographically disconnected, and can be used as evidence of last-known populations. Optimal linear estimation (OLE) modelling was developed to reliably reconstruct the timing of species extinction dates using such traces (Roberts and Solow, 2003; Solow, 2005; Rivadeneira et al., 2009; Clements et al., 2013). Here, we use OLE to reconstruct the timing of the end of the Acheulean cultural tradition based on the archaeological equivalent of ‘last-seen’ species evidence - dated late Acheulean sites.
Methods
Defining the Acheulean
At its broadest, Acheulean assemblages are characterised by the presence of large bifacially flaked cutting tools (handaxes and cleavers, which do not occur prior to 1.75 Mya) and the absence of MP and MSA technologies (most notably Levallois hierarchical flaking) (Shea, 2017). There can be variation in biface form, alongside some technological differences (McNabb et al., 2004; Lycett and Gowlett, 2008; Sharon et al., 2011; Key, 2019; Shipton, 2020). As noted above, however, regional variation exists in how the Acheulean is defined; most notably through transitional cultural traditions that incorporate other technological elements.
For the present analyses we defined a site as belonging to the Acheulean cultural tradition based on two factors: (1) the presence of large bifacially flaked cutting tools (handaxes and cleavers) and an absence of Levallois technologies, and (2) the original authors describing a site also assigning it to the Acheulean. We are aware that not all individuals will be happy with this definition. Some sites could be excluded or included based on a researcher’s technological and typological predilections concerning the Acheulean, or debate over the accuracy of published dates. The analysis performed here is, however, designed to understand the extinction of the Acheulean cultural tradition at global and continental levels. In turn, it necessitated a broad definition to guarantee that sites feeding into each model were not unduly excluded based on individual pedagogy-based interpretations of the Acheulean (most notably our own). Simultaneously, we wanted to respect the interpretation of those individuals that know the relevant artefacts best, and the peer-review process in determining the accuracy of published dates (although also note the resampling procedures used in the section “OLE modelling methods”). Our definition of the Acheulean is not limited to stone artefacts. As with any discussion of the Acheulean, however, poor preservation rates in organic artefacts impede our ability to include them in our analyses (for example, Acheulean sites defined by bone handaxes/cleavers are included in our definition, but there are no examples late enough for inclusion).
We did not include sites (or assemblages/artefact layers) assigned to transitional cultures; although we do appreciate that the Acheulean ended in a technologically mosaic-like nature in some regions (e.g. Moncel et al., 2020b). Similarly, we do not distinguish between late Acheulean sites containing specific handaxe and cleaver forms (be it due to distinct evolutionary trajectories, or any other process). Application of this sampling procedure allows us to model the end of the Acheulean as a distinct cultural tradition, and thus, as it is recognised for the majority of its existence. An exception was made for the Fauresmith (which include blades and prepared cores alongside bifaces [McBrearty and Tryon, 2006; Porat et al., 2010; Herries, 2011]), where an additional ‘Sub-Saharan African’ model was run that incorporated these sites. Our intention was to demonstrate the impact of including transitional assemblages when inferring Acheulean extinction dates.
Site identification and model classifications
A comprehensive literature review was undertaken to identify ‘late’ or ‘final’ Acheulean sites across Africa and Eurasia. All of those assigned as late Acheulean on typological or morphological grounds alone were not considered (due to their subjective nature and requirements for absolute dates). Sites were then divided into four continental regions; Europe, Asia, Sub-Saharan Africa, and North Africa and the Near East (Table 1; Fig. 2). Only the most recent (youngest) 10 sites in each region were required, as is generally recommended (Roberts and Solow, 2003; Solow, 2005; Rivadeneira et al., 2009). Indeed, OLE reconstructs extinction timings through the chronological spacing of the most recent records and the inclusion of additional older sites (i.e., greater numbers of sites further away from the last known occurrence) would have little impact on the result (or even could detrimentally impact it; see the section “Applying OLE modelling to the archaeological record”). As suggested by Solow (2005), overly large numbers of records used would risk invalidating the assumptions of the model.

As indicated in Fig. 1, not all areas currently have strong or continued evidence of the Acheulean being present (e.g. Congo River Basin). Original satellite image credit: NASA Visible Earth Project.
Geographic biases in research focus mean that some countries have disproportionately greater numbers of late Acheulean sites compared to others. Such weighting results from historical and political biases unrelated to how Acheulean hominins are actually represented geographically and chronologically. Thus, there was potential for continental-level models to be overly influenced by individual countries. To counter this, each country was able to contribute a maximum of three sites to a continental model, meaning that at least four countries were represented in each. All continental models adhered to this except for Asia, as only three countries have dated sites from the terminal phase of the Acheulean (China, India and South Korea, with the latter contributing four sites). In addition, countries could only contribute one date from a localised region (for example, the Middle Son Valley [India]). This ensured that a heavy focus on localised regions could not disproportionately influence models. For these reasons, there will be some late Acheulean sites excluded from these models despite being younger than others that are included.
Two additional continental models were run (Table 1). First, we re-ran the Sub-Saharan African model but included sites assigned to the Fauresmith. This resulted in the four oldest sites in the original model being removed and replaced with younger Fauresmith alternatives. Second, we re-ran the Asian model after the removal of the five youngest sites in this region (i.e. n = 5). The five removed sites fit the criteria for inclusion in our models, however, they provide extremely late occurrences of large bifacial technology and some are critical of their inclusion in the Acheulean (Shipton and Petraglia, 2011; Kuman et al., 2014; Lee, 2017; Bae, 2017; Li et al., 2018). The second Asian model therefore helps us understand the extinction of the Acheulean cultural tradition in Asia irrespective of one’s perspective on these more recent artefacts.
Two models using intercontinental data were run to better understand the end of the Acheulean as a global cultural tradition (Table 1). Fauresmith sites were not included in either. In the first, the 10 youngest sites from those included in the continental models (n = 40), irrespective of their country of origin, were included. This meant that a country could contribute a maximum of three sites, but there were no limits on which continents these countries came from (i.e. individual continents could contribute heavily to the model). The second model included only one site per country, and a maximum of three sites per continental region (the youngest in each instance). This meant that all four continental regions contributed to the modelling process. For both, versions including and excluding the five debated young Asian sites were run.
We recognise that the dates assigned to some sites included in the models are subject to ongoing debate. However, it is not the role of this study to critique specific sites and their geochronology. If we were to remove every site subject to debate by a specific researcher or research group, then we would have very few sites available for inclusion (if any at all). Thus, we must go with the consensus view that appears in the literature and trust in the value of peer-review when assessing the reliability of a site’s published date. This does not discount the views of any individuals; simply, it is necessary for a standardised site sampling procedure to be used. If we were to make exceptions for individual researchers, then we would invalidate the integrity of the site-sampling procedure.
In total, 10 model classifications were examined. All sites and dates input into each model are available in Supplementary Material 1.
Applying OLE modelling to the archaeological record
Here we apply the optimal linear estimation (OLE) method as proposed by Roberts and Solow (2003) for dating extinctions. The OLE method has proved to be robust in the inference of extinction under a variety of scenarios (Rivadeneira et al., 2009; Clements et al., 2013). Although most regularly applied to model the extinction of faunal and floral species, the underlying assumptions of the OLE method are not specific to biological organisms. Instead, the method can readily be applied to diverse phenomena so long as they are characterised by sporadic observations through time prior to their extinction.
OLE uses the last known chronological occurrences, or ‘sightings’, of a given phenomenon to estimate how long it continued after the final (most recent) confirmed occurrence. In straightforward terms, it models the date when a process generating the occurrences had stopped, and the phenomenon will thus no longer be observable. In the case of the Acheulean (or any other material culture), this will be the point at which the knowledge of the cultural tradition is no longer being expressed in the form of archaeologically detectable artefacts.
OLE is based on the result that most recent sightings have the same ‘Weibull form’ regardless of the characteristics of the full record, its density and distribution (Roberts and Solow, 2003; Solow, 2005). Put simply, the model asks given the distribution of the sightings, what is the probability another sighting would not have occurred (from which we can infer ‘extinction’). Thus, the model does not contain any biologically specific parameters and it is applicable to asking questions concerning the extinction of cultural traditions. As such, the applicability of the model beyond the extinction of species has been well recognised, and it has been used in diverse fields, including geological stratigraphy (Marshall, 2010), phenology (Pearse et al., 2017), and phylogenetics (Vinh and von Haesleler, 2004). The method takes into account the intervals between the last known observations of a phenomenon and their distribution, irrespective of whether it is applied to model the extinction of biological species or cultural traditions (for further information see, Solow, 2005; Rivadeneira et al., 2009; Clements et al., 2013; Boakes et al., 2015).
OLE modelling methods
Due to the paucity of the archaeological record, a fast end to the Acheulean (e.g. <1000 years) cannot be automatically assumed after the final occurrences of dated sites. Thus, we did not apply the classical extinction method described by Solow (1993) that assumes instantaneous extinction.
Let T1 > T2 > … > Tk be the k youngest sites, ordered from the youngest (with Tk being the first known site date). Interest centres on using this record of site dates to estimate the extinction time, \(\theta\). In this context, the optimal linear estimation is based on the fact that the joint distribution of the k youngest sites has the same approximate ‘Weibull form’, regardless of the parent distribution of the complete record of dated sites.
The optimal linear estimator of θ has the form of a weighted sum of the site date times (Roberts and Solow, 2003),
The vector of weights is given by
where e is a vector of k 1’s and ∧ is the symmetric k × k matrix with typical element \(\lambda _{ij} = \left( {{\Gamma}\left( {2\widehat v + i} \right){\Gamma}\left( {\widehat v + j} \right)} \right)/\left( {{\Gamma}\left( {\widehat v + i} \right){\Gamma}\left( j \right)} \right),\,j \le i\), and where Γ is the standard gamma function. Also
is an estimate of the shape parameter of the joint Weibull distribution of the k youngest sites date times. Following Solow (2005), an approximate one-sided upper bound of a 1−α confidence interval (CI) for θ is
where \(c\left( \alpha \right) = \left( {\frac{k}{{ - {\mathrm {log}}\alpha }}} \right)^{ - \widehat v}\); note that in Solow (2005) the equation for c(α) was incorrectly inverted.
An advantage of the OLE method is that it takes into consideration the distribution of only the most recent records (in this case the youngest Acheulean sites), which obviated the need for including more ancient Acheulean sites in the analysis. We followed previous studies (Roberts and Solow, 2003; Solow, 2005; Rivadeneira et al., 2009) in using the first 5–10 (k) youngest sites dates for each region. As such we date the extinction of the Acheulean cultural tradition in each region. However, as there is no specific start date for the time series, the 10th youngest site date is used as the beginning of the period. As the end point is non-independent to the time series records, k reduces by 1 (Solow, 1993; Clements et al., 2013). The OLE method produces two types of estimates relevant to understanding the timing of extinctions. The first is TE, which here represents the estimated extinction date for the Acheulean in a given model. TE is presented as years before present (BP). The second, TCI, is the upper bound of each model’s 1−α confidence interval. This is effectively the time beyond which the probability of the tradition still existing is below α. We chose α = 0.05 as the extinction threshold value (Roberts and Solow, 2003). The extinction date for each region was calculated using the R software package sExtinct (Clements, 2013).
Since most dated Acheulean sites provide age ranges, mean values were used for the main analysis. Additionally, to address the uncertainty of some age estimates, a resampling approach was also applied. Dates of each site were randomly drawn from a normal distribution, with the mean value represented by the mean of the age range, and standard deviation equal to the half of the difference between the mean value and range bounds. Such randomly generated datasets were consequently assessed with the OLE method, and the whole procedure was repeated 10,000 times, with results expressed as mean and median values across all iterations.
Results
The end of the Acheulean: continental dates
The four continental regions returned dates spanning >120 Kya, demonstrating the Acheulean to end at different times across the world (Table 2; Fig. 3). Late Acheulean sites from North Africa and the Near East provide an extinction date estimated at 174 Kya. The Sub-Saharan African model returned similar dates, with the end of the Acheulean occurring 170–166 Kya. The Acheulean is demonstrated to end slightly later in Europe, with the techno-complex’s demise happening between 141 and 130 Kya. The Asian OLE model predicted substantially later dates, with the end of the Acheulean occurring 57–53 Kya. Even with the removal of the five debated Asian sites, a similarly recent end to the Acheulean is predicted (57–56 Kya). As expected, the inclusion of the Fauresmith in the Sub-Saharan African model resulted in a later extinction date for this region (73–68 Kya). There was strong consistency in dates between the mean estimate and resampling methods (Table 2).

A Sub-Saharan African model including Fauresmith sites is also displayed, as is the modified Asian (n = 5) model. Data are derived from 10,000 iterations of the random sampling method. Negative outliers in the Asian models extended beyond ‘0’, and are not visible.
TCI values varied between the two methods, and provided diverse 95% confidence intervals between continental regions (Table 2). The sporadic nature of the archaeological record and the tens of thousands of years observed between some of the sites entered into each model explains this diversity. On occasion a negative value was returned, indicating a date in the future (from the present day). This is not meaningful, of course, as the Acheulean no longer exists, and instead reflects the aforementioned nature of the data.
The end of the Acheulean: global dates
Two sampling methods were used to identify sites for the global-level models (Table 1). The first (‘Global 1’) used the 10 youngest Late Acheulean sites from all continental regions, with no limit on where site contributions came from (bar the maximum of three per country). The second (‘Global 2’), which provided stricter limits on how the Acheulean was defined at a global level, required all four continental regions to contribute to the model. Date estimates based on the Global 1 OLE models varied between 52 and 50 Kya, but increased to 68–62 Kya when the five debated Asian sites were excluded (i.e. ‘Global 1b’). The further exclusion of Bhimbetka, India (another site debated by some), increased estimates to 107–105 Kya. A similar pattern exists for the Global 2 models, where Acheulean extinction estimates increase from between 29–32 Kya, to 56–45 Kya (Table 3; Fig. 4), and then 97–95 Kya with the removal of Bhimbetka.

Negative outliers in all models are not displayed.
Dates contributing to the Global 1 models were more tightly clustered around the most recent (i.e. ‘last seen’) records of the Acheulean at a global level, which resulted in a steeper incline of the model’s predictive slope after this occurrence. Effectively, this meant that the end of the cultural tradition was estimated to occur sooner after its final record, explaining why confidence intervals are less varied and remain positive for the Global 1 estimates (Table 3). As Global 2 estimates placed greater geographic limits on where site contributions (dates) came from, there was weaker clustering before the most recent record, which in turn resulted in greater confidence intervals.
Discussion
The Acheulean lasted for over 1.5 million years, spreading widely across Africa and Eurasia. Our understanding of the period’s origin is relatively clear; multiple ~1.75 million-year-old sites support its emergence in East Africa (de la Torre, 2016), and little suggests otherwise. The end of the Acheulean, however, has always been more problematic, being limited to isolated dates from individual sites (e.g. Bates et al., 2014; Scerri et al., 2018) or broad 0.3–0.15 Mya statements drawn from overviews of the literature (McBrearty and Tryon, 2006; Stout, 2011; Fontana et al., 2013; Shea, 2017; Galway-Witham et al., 2019). Here, we use optimal linear estimation modelling and radiometric dates from 44 late Acheulean sites to provide the most accurate estimation yet of when the cultural tradition ended.
Continental-level models reveal broad consensus among estimates for Africa, Europe, and the Near East. North Africa and the Near East provided the oldest dates at 174 Kya, followed closely by Sub-Saharan Africa 170–166 Kya. European estimates are notably later, with its regional extinction estimated to be between 141 and 130 Kya. These dates are later than most statements in the literature concerning the end of the Acheulean in these regions (e.g. Deino et al., 2018; Méndez-Quintas et al., 2020), although not by a substantial margin. Certainly, some Acheulean sites provide dates approaching the model estimates (e.g. Porat et al., 2002; Michel et al., 2009; Herries, 2011). This is to be expected as these are what the OLE models are built on.
What we have done here, however, is estimate how long the Acheulean continues after the youngest sites that we currently know about. For example, the most recent European Acheulean site, Arbo (Spain), dates to 161 Kya (Méndez-Quintas et al., 2019), yet the OLE models predict the tradition to continue for an additional 20,000 years in this region. Saffaqah (Saudi Arabia), however, is the most recent Acheulean site in the Near East and North African model, dating to 188 Kya (Scerri et al., 2018). Our modelling estimates the Acheulean to end here by 174 Kya, putting Saffaqah close to the traditions chronological limit. We do not, and likely never will, know where the final archaeological instances of the Acheulean are; thus, OLE estimates are the most accurate method we have for inferring when the Acheulean ended.
The inclusion of Fauresmith assemblages in the Sub-Saharan model decreased estimated dates considerably. This was not unexpected; Fauresmith artefacts are young compared to other Late Acheulean sites in this region (particularly Abdur Reef [Bruggemann et al., 2004]). The difference between the ‘last seen’ occurrence (i.e. Abdur Reef) and the estimated extinction date is, however, large (~55 Kya). Again, this is not surprising; few sites cluster around Abdur Reef’s late date, meaning that there is a long period without an Acheulean site prior to the last time the tradition is seen. This has the effect of increasing the predicted length of time that the Acheulean could have existed after the artefacts at Abdur Reef. This raises an important point concerning how the OLE method is impacted by the sporadic nature of archaeological finds, and whether an absence of dates immediately prior to a ‘last seen’ occurrence artificially decreases the tradition’s estimated end date. For example, had another site been found in Sub-Saharan Africa dating to 135 Kya (i.e. only 10 Kya older than Abdur Reef), it is likely that the model would have produced an older end date (see the North African and Near East model for an example of when chronologically tighter site clustering predicts a faster end to the tradition). Therefore, we want to stress that OLE estimates are only as accurate as the radiometric dates and definitions available in existing literature. As new archaeological discoveries are made, it is possible that our inferences will not stand the test of time. Later instances of the Acheulean will push the extinction of the tradition even later, while other finds close to the ‘last seen’ occurrences used here could push it back in time (as described above). However, the present extinction inferences are robust in light of current archaeological knowledge.
It is also important to consider the varied definitions used for the Acheulean, and how this could have impacted the models. The Asian models highlight this predicament well. When all Acheulean sites in Asia are taken at face value, the tradition is predicted to end between 57 and 53 Kya. This appears to be due to a series of <100 Kya handaxe sites in South Korea and China. Yet, the inclusion of these assemblages in the Acheulean is controversial, both for their late occurrence and aspects of their form/production (Norton et al., 2006, 2009; Lycett and Gowlett, 2008; Shipton and Petraglia, 2011; Bae, 2017; Lee, 2017; Dennell, 2018; Li et al., 2018). Thus, while these sites adhere to standard technological definitions (see the section “Defining the Acheulean”), some would contest their inclusion. This is why we re-ran the Asian model but excluded the five controversial sites, such that the most recent Acheulean site in Asia was the 106 Kya Bhimbetka rockshelter III (India) (Bednarikm et al., 2005; Petraglia et al., 2012). Even with the exclusion of the controversial younger sites, however, the Asian model predicted the Acheulean to end 57–56 Kya (with Bhimbetka removed as well the model still predicts an end 59–46 Kya). This suggests that, although positioned towards the extreme end of the tradition, handaxe assemblages from Houfang and Danjiangkou in China and Unjeong and Wolso in South Korea (among others) are not outside of the expected chronology for the Acheulean in this region. Future debate on their inclusion within the Acheulean cultural tradition should, therefore, focus on technological and morphological aspects (which includes their marked thickness and reduced elongation [Lee, 2017]).
The Acheulean as a global cultural tradition is demonstrated to end between 107 and 29 Kya. Some of this variation relates to the inclusion or removal of the aforementioned Asian sites. Equally, however, variation is driven by how the Acheulean as a global tradition is defined. If there are no geographic restrictions on the sites contributing to the model, then the Acheulean is predicted to end between 52 and 50 Kya (or 68–62 Kya, excluding the five Asian sites, and 107–105 Kya when also excluding Bhimbetka). When definitions are stricter, and incorporate all four continental regions on a broadly equal basis, the Acheulean is estimated to end between 29 and 32 Kya (or 56–45 Kya and 97–95 Kya, respectively, when excluding the Asian sites). This decrease pushes the end of the Acheulean further towards the end of the Middle Palaeolithic period. An explanation for this can be seen in how sites cluster around the youngest dates in each model. ‘Global 1’ models demonstrate tight clustering, while those in the ‘Global 2’ models extend over a greater period of time, resulting in a less sharp incline after the last seen date.
Both sampling methods have merit, however, we propose that an equal number of site contributions from each region is not necessary to define the Acheulean at a global level. Instead, if the Acheulean is present in two or more continental regions then we would argue that the tradition is still present at a global level. Thus, we favour the ‘Global 1’ estimates for when the Acheulean ended as a global cultural entity (i.e. between 107 and 50 Kya). Either way, the Acheulean is expected to continue on a global level after 100 Kya, most likely being restricted to smaller and more isolated geographic pockets within continental regions as time progresses. This would help to explain why some of the youngest sites exist in the extremes of the tradition’s range.
Overlap between the Acheulean and Middle Palaeolithic/Middle Stone Age
In all continental-level models, dates overlap with the emergence of MP and MSA technologies. The earliest MSA sites in Sub-Saharan Africa occur at Kathu Pan (South Africa), Florisbad (South Africa), and Olorgesailie (Kenya) (Kuman et al., 1999; Porat et al., 2010; Deino et al., 2018), and date to 291, 280, and 320 Kya (respectively). Thus, the Acheulean cultural tradition is demonstrated to overlap with MSA technological behaviours for over 110 Kya. The MP in the Near East displays similarly early dates (e.g. 335–240 Kya [Adler et al., 2014; Zaidner and Weinstein-Evron, 2020]), evidencing regional overlap of 160–70 Kya. France, Italy and the UK have early MP sites dating to between MIS 12 and MIS 8 (Déspriee et al., 2009; Moncel et al., 2020a), suggesting Acheulean and MP technological traditions to co-exist in Europe for over 250 Kya. Middle Palaeolithic assemblages in Asia date to as early as 380 Kya (Norton et al., 2009; Akhilesh et al., 2018), indicating an overlap of up to ~330 Kya years. We are not suggesting that there was constant cultural overlap; it would have been punctuated and dependent on population dynamics, extinctions and colonisation events (Fig. 5).

Distributions of each technology are represented from ~550 to ~50 Kya (top to bottom), respectively. Although hypothetical, the population dynamics outlined here demonstrate how localised extinctions, colonisation waves and admixture likely influenced the distribution of the Acheulean. It is still unknown whether the MP/MSA originated in one location before spreading to alternative populations, or it independently originated in multiple regions. Here, we have illustrated the latter scenario.
Our results reveal that across the globe, the Acheulean overlaps with alternative stone tool cultures for substantial periods. As a result, a shift in how the Acheulean is defined at a chronological level is necessary. Either the Acheulean can no longer be considered exclusive to the Lower Palaeolithic (LP) period or Early Stone Age (ESA), or the LP (and ESA) as a technological entity persisted alongside the Middle Palaeolithic (and MSA). In both scenarios, populations practising Acheulean and Middle Palaeolithic (and MSA) technological behaviours would have coexisted. Definition of the Acheulean as both a LP and MP (or, ESA and MSA) cultural entity arguably provides greater consistency with existing techno-temporal frameworks.
Cultural overlap between the LP and MP is not a new suggestion (Isaac, 1972; Norton et al., 2009; Villa, 2009), and multiple research articles detail instances of the Acheulean continuing into the MP/MSA (McBrearty and Tryon, 2006; James and Petraglia, 2009; Norton et al., 2009; Hublin, 2009; Haslam et al., 2011; Fontana et al., 2013; Akhilesh et al., 2018; Mendez-Quintas et al., 2019, 2020). Santonja et al. (2016) have argued for such a scenario in the Iberian peninsula, even linking different technologies to the presence of different hominin populations. Scerri et al. (2018, p. 6) provide similar evidence at Saffaqah (Arabian Peninsula), where late Acheulean populations “overlap with an emerging Middle Palaeolithic”. What is provided here, however, is evidence of the strength of this overlap, and that irrespective of whether the Acheulean is considered at a continental or global level, it can no longer be restricted to a simple presence and absence dichotomy between the LP and MP (or ESA and MSA).
Implications for Late Acheulean hominin demography
Evidence of overlap between Acheulean and MP (and MSA) technologies indicates the co-existence of hominin populations practicing different stone tool behaviours. The widespread and extended overlap predicted here suggests that the arrival of MP technologies did not quickly replace its technological precursor in all instances. Rather, some populations continued to maintain Acheulean cultural traditions in spite of alternative technologies being practised elsewhere.
Potentially, the duration of technological overlap demonstrated in Africa, Europe and Asia (i.e. in the region of 300–100 Kya) is how long it took for these new technological ideas to permeate through social systems. Certainly, hominin populations were likely highly dispersed during the late Pleistocene (Dennell et al., 2011; Bocquet-Appel and Degioanni, 2013). However, given how fast cultural information can be transmitted in human populations we consider this unlikely. An alternative explanation is the existence of barriers limiting the spread of MP and MSA technologies (and in turn, the demise of the Acheulean) between hominin populations.
Physical and environmental barriers are known to prevent or mediate the spread of cultural information in non-human great apes and modern humans. Moreover, there are suggestions this occurred during the Mid-to-Late Pleistocene (Henshilwood and d’Errico, 2005; James and Petraglia, 2005; Lycett and Norton, 2010; Shea, 2017; Arroyo et al., 2019). Thus, it is plausible that on occasion distance, mountain ranges, seas, and deserts created enough of a barrier to prevent the transmission of new technologies to Acheulean populations (particularly in north east Asia [Lycett and von Cramon-Taubadel, 2008; Lycett and Norton, 2010]). Climate change, mortality, and misfortune could have similarly influenced transmission (Dennell, 2018). However, given the substantial geographic and chronological peripheries discussed, such barriers could not have existed in all instances where overlap exists. Thus, two alternative scenarios can be suggested.
Potentially, the benefits conveyed by MP/MSA technologies were not strong enough to result in the consistent uptake of these new technologies when opportunities arose. Simply, individuals within Acheulean populations did not experience benefits enough to warrant spending time and energy learning new production techniques. Again, we consider this explanation unlikely given that MP/MSA technologies were adopted widely during this period (although we do not discount localised instances of this occurring) and multiple significant benefits have been demonstrated for MP technologies (e.g. Eren and Lycett, 2012; Shimelmitz and Kuhn, 2018).
Alternatively, it is possible that cognitive and anatomical barriers prevented the transmission of these new technologies, either through populations being unable to undertake more demanding tool production and use activities, or differences altering relevant cost benefit ratios. Indeed, hierarchical flaking is cognitively demanding (Stout, 2011), and Levallois tools have been suggested to indicate greater capabilities in MP/MSA species relative to those associated with the Acheulean (Foley and Lahr, 1997; McBrearty and Tryon, 2006; Shipton, 2016; Otte, 2019). We are not suggesting that transitioning between Acheulean and MP/MSA technologies always necessitates a cognitive leap. James and Petraglia (2009) discuss how Acheulean and MP technological overlap can reflect continuity of cognitive capabilities. Rather, cognitive differences between populations (perhaps species) may have prevented the uptake of MP/MSA technologies by some, even when opportunities for the transmission of these technologies occurred. The manual demands of Levallois flaking are not well understood and could also plausibly be greater than that required for Acheulean technologies (even if their use was not [Key et al., 2020]). Certainly, anatomical differences between Late Pleistocene hominin populations could have restricted lithic technological developments (Niewoehner, 2006; Marzke, 2013; Key and Lycett, 2018).
In recent years, fossil and genetic evidence has confirmed the co-existence of multiple hominin species during the later Middle Pleistocene of Africa and Eurasia. Evidence of admixture between some of these species confirms a degree of interaction (Browning et al., 2018; Villanea and Schraiber, 2019; Rogers et al., 2020), while anatomical comparisons highlight manipulative and cognitive differences varying in scale and nature (e.g. Tocheri et al., 2008; Holloway et al., 2018; Détroit et al., 2019; Galway-Witham et al., 2019). Unfortunately, resolution on how these populations relate to the varying lithic technologies present during this time is often lacking. Nonetheless, it is not unreasonable to predict that the anatomical and cognitive differences observed between these species had the potential to result in the maintenance of different cultural traditions. Either through an inability to effectively use/produce some lithic technologies, or changes to relevant cost benefit ratios. Again, Asia provides a suitable (but not the only [e.g. Hawks and Berger, 2020]) example, with H. erectus (Rizal et al., 2020), H. floresiensis (Aiello, 2010), and H. luzonensis (Détroit et al., 2019) all displaying anatomy that could potentially limit their ability to produce and use MP technologies.
Parsimony suggests the common ancestor of Neanderthals and anatomically modern humans to be capable of producing MSA/MP technologies. In turn, it is logical to link the last known occurrences of the Acheulean with species that share a common ancestor with modern humans prior to our split with Neanderthals. Currently, this includes the aforementioned Asian Late Pleistocene hominin species, along with H. naledi, and H. heidelbergensis (s.l.) populations separate to those that evolved into Neanderthals and modern humans. However, we again stress the dynamic nature of cultural and biological evolutionary pathways (Lycett and von Cramon-Taubadel, 2015; Scerri et al., 2019), and there is no specific reason that some Neanderthal, Denisovan and modern human populations could not also have reverted to or continued the Acheulean cultural tradition.
It is not our intention to discuss individual regions or species in detail. Nor is it within the scope of the paper to discuss precisely why regions transitioned away from the Acheulean at different times (although cultural, biological/species-related and ecological factors could be involved). Rather, we wish to stress that evidence of cultural overlap and increasing complexity in the distribution of late Middle Pleistocene lithic technologies is to be expected given increasing diversity in the fossil record (Wood and Boyle, 2016; Galway-Witham et al., 2019). Indeed, there is evidence of geographic and temporal overlap between multiple hominin species between 300 and 50 Kya (e.g. Dirks et al., 2017; Jacobs et al., 2019). The cultural scenario outlined here therefore mirrors the dynamic nature of the Middle Pleistocene hominin fossil record. Moreover, they are likely linked, with contemporaneous hominin species engaging in distinct stone-tool cultural practises in multiple regions around the world.
Conclusion
Presented here are the most accurate estimates yet for when the Acheulean cultural tradition ended. We do so using optimal linear estimation, a modelling technique often used to estimate the extinction of faunal species, but novel to archaeological research. In Africa and the Near East the Acheulean is predicted to end 174–166 Kya. In Europe, the Acheulean is predicted to end 141–130 Kya. These dates are only slightly younger than current understanding on the end of the tradition in these regions. Asian estimates, however, range between 57 and 53 Kya. Thus, the Acheulean continues in this region long after it has ended elsewhere, and for the majority of the MP. The Acheulean stopped being a global tradition between 107 and 29 Kya, although we favour an age bracket of between 107 and 50 Kya. These estimates suggest the Acheulean to have remained a distinct cultural tradition long after the inception of MP technologies in multiple continental regions. Persistence of the tradition in Europe and Asia may be linked to each region’s geographic isolation, relative to the rest of the Acheulean world. In line with the increasingly dynamic nature of the Middle Pleistocene hominin fossil record, contemporaneous hominin populations are demonstrated to have been practicing distinct stone-tool behaviours, potentially due to cognitive and anatomical differences.
Data availability
All data are available in the relevant Supplementary Information.
Change history
28 October 2021
A Correction to this paper has been published: https://doi.org/10.1057/s41599-021-00932-5
References
Adler DS, Wilkinson KN, Blockley S et al. (2014) Early Levallois technology and the Lower to Middle Paleolithic transition in the Southern Caucasus. Science 345(6204):1609–1613
Aiello LC (2010) Five years of Homo floresiensis. Am J Phys Anthropol 142(2):167–179
Akhilesh K, Pappu S, Rajapara HM et al. (2018) Early Middle Palaeolithic culture in India around 385-172 ka reframes Out of Africa models. Nature 554:97–101
Arroyo A, Proffitt T, Key A (2019) Morphometric and technological analysis of Acheulean large cutting tools from Porzuna (Ciudad Real, Spain) and questions of African affinities. J Archaeol Sci: Rep 27:101992
Bae K (2017) Paleolithic archaeology in Korea. In: Habu J, Lape P, Olsen J (eds) Handbook of East and Southeast Asian archaeology. Springer, New York, pp. 219–239
Bates MR, Wenban-Smith FF, Bello SM et al. (2014) Late persistence of the Acheulian in southern Britain in an MIS 8 interstadial: evidence from Harnham, Wiltshire. Quat Sci Rev 101:159–176
Bednarikm RG, Kumar G, Watchman A et al. (2005) Preliminary results of the EIP project. Rock Art Res 22(2):147–197
Beyene Y, Katoh S, WoldeGabriel G et al. (2013) The characteristics and chronology of the earliest Acheulean at Konso, Ethiopia. Proc Natl Acad Sci USA 110(5):1584–1591
Boakes EH, Rout TM, Collen B (2015) Inferring species extinction: the use of sighting records. Methods Ecol Evol 6(6):678–687
Bocquet-Appel JP, Degioanni A (2013) Neanderthal demographic estimates. Curr Anthropol 54(S8):S202–S213
Browning SR, Browning BL, Zhou Y et al. (2018) Analysis of human sequence data reveals two pulses of archaic Denisovan admixture. Cell 173(1):53–61
Bruggemann JH, Buffler RT, Guillaume MMM et al. (2004) Stratigraphy, palaeoenvironments and model for the deposition of the Abdur Reef Limestone: context for an important archaeological site from the last interglacial on the Red Sea coast of Eritrea. Palaeogeogr Palaeoclimatol Palaeoecol 203(3–4):179–206
Brumm A, Rainey A (2011) The Acheulean downunder: modern human ‘handaxes’ from the Barkly Tableland of northern Australia. Lithics 32:50–61
Clements C (2013) sExtinct R Package. https://cran.rproject.org/src/contrib/Archive/sExtinct/ Accessed 16 Jul 2020
Clements CF, Worsfold NT, Warren PH et al. (2013) Experimentally testing the accuracy of an extinction estimator: Solow’s optimal linear estimation model. J Animal Ecol 82:345–54
Corvinus G (2004) Homo erectus in East and Southeast Asia, and the questions of the age of the species and its association with stone artifacts, with special attention to handaxe-like tools. Quat Int 177(1):141–151
Deino AL, Behrensmeyer AK, Brooks AS et al. (2018) Chronology of the Acheulean to Middle Stone Age transition in eastern Africa. Science 360(6384):95–98
Dennell R (2009) The Palaeolithic settlement of Asia. Cambridge University Press, Cambridge
Dennell R (2018) The Acheulean assemblages of Asia: a review. In: Gallotti R, Mussi M (eds) The emergence of the Acheulean in East Africa and beyond: contributions in Honor of Jean Chavaillon. Springer, Cham, pp. 195–214
Dennell RW, Martinon-Torres M, Bermudez de Castro J (2011) Hominin variability, climatic instability and population demography in Middle Pleistocene Europe. Quat Sci Rev 30(11–12):1511–1524
Déspriee J, Voinchet P, Gageonnet R et al. (2009) Les vagues de peuplements humains au Pléistocène inférieur et moyen dans le bassin de la Loire moyenne, région Centre, France. Apports de l’étude des formations fluviatiles. L’Anthropologie 133(1):125–167
Détroit F, Mijares AS, Corney J et al. (2019) A new species of Homo from the Late Pleistocene of the Philippines. Nature 568:181–186
Diez-Martín F, Yustos PS, Uribelarrea D et al. (2015) The origin of the Acheulean: the 1.7 million-year-old site of FLK West, Olduvai Gorge (Tanzania). Sci Rep 5:17839
Dirks PHGM, Roberts ER, Hilbert-Wolf H et al. (2017) The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife 6:e24231
Eren MI, Lycett SJ (2012) Why Levallois? A morphometric comparison of experimental ‘preferential’ Levallois flakes versus debitage flakes. PLoS ONE 7(1):e29273
Foley R, Lahr MM (1997) Mode 3 technologies and the evolution of modern humans. Camb Archeol J 7(1):3–36
Fontana F, Moncel M-H, Nenzioni G et al. (2013) Widespread diffusion of technical innovations around 300,000 years ago in Europe as a reflection of anthropological and social transformations? New comparative data from the western Mediterranean sites of Orgnac (France) and Cave dall’Olio (Italy). J Anthropol Archeol 32(4):478–498
Gallotti R, Mussi M (2018) The emergence of the Acheulean in East Africa and beyond: contributions in Honor of Jean Chavaillon. Springer, Cham
Galway-Witham J, Cole J, Stringer C (2019) Aspects of human physical and behavioural evolution during the last 1 million years. J Quat Sci 34(6):355–378
Gowlett JAJ (1997) Why the muddle in the middle matters: the language of comparative and direct in human evolution. Arch Pap Am Anthropol Assess 7(1):49–65
Gowlett JAJ (2015) Variability in an early hominin percussive tradition: the Acheulean versus cultural variation in modern chimpanzee artefacts. Philos Trans R Soc B 370(1682):20140358
Haslam M, Roberts RG, Shipton C et al. (2011) Late Acheulean hominins at the Marine Isotope Stage 6/5e transition in north-central India. Quat Res 75:670–682
Hawks J, Berger L (2020) On Homo naledi and its significance in evolutionary anthropology. In: Deane-Drummond C, Fuentes A (eds) Theology and evolutionary anthropology. Routledge, Abingdon
Henshilwood C, d’Errico F (2005) Being modern in the Middle Stone Age: individuals and innovation. In: Gamble C, Poor M (eds) The Hominid individual in context. Routledge, Abingdon, pp. 244–264
Herries AIR (2011) A chronological perspective on the Acheulian and its transition to the Middle Stone Age in southern Africa: the question of the Fauresmith. Int J Evol Biol 2011:961491
Holloway RL, Hurst SD, Garvin HM et al. (2018) Endocast morphology of Homo naledi from the Dinaledi Chamber, South Africa. Proc Natl Acad Sci USA 115(22):5738–5743
Hopkinson T (2007) The transition from the Lower to the Middle Palaeolithic in Europe and the incorporation of difference. Antiquity 81(312):294–307
Hublin JJ (2009) The origin of Neanderthals. Proc Natl Acad Sci USA 106(38):16022–16027
Isaac G (1972) Chronology and the tempo of cultural change during the Pleistocene. In: Bishop W, Miller J (eds) Calibration of Hominid evolution. Scottish Academic Press, Edinburgh, pp. 381–430
Jacobs Z, Li B, Shunkov MV et al. (2019) Timing of archaic hominin occupation of Denisova Cave in southern Siberia. Nature 565:594–599
James HVA, Petraglia MD (2005) Modern human origins and the evolution of behaviour in the later Pleistocene record of South Asia. Current Anth 46:S3–S27
James H, Petraglia MD (2009) The Lower to Middle Palaeolithic transition in South Asia and its implications for hominin cognition and dispersals. In: Camps M, Chauhan PR (eds) Sourcebook of Palaeolithic transitions. Springer, New York, pp. 255–264
Key AJM (2019) Handaxe shape variation in a relative context. C R Palevol 18(5):555–567
Key AJM, Lycett SJ (2018) Investigating interrelationships between Lower Palaeolithic stone tool effectiveness and tool user biometric variation: implications for technological and evolutionary changes. Archaeol Anthropol Sci 10:989–1006
Key AJM, Farr I, Hunter R et al. (2020) Muscle recruitment and stone tool use ergonomics across three million years of Palaeolithic technological transitions. J Hum Evol 144:102796
Kuman K, Inbar M, Clarke RJ (1999) Palaeoenvironments and cultural sequence of the Florisbad Middle Stone Age hominid site, South Africa. J Archeol Sci 26(12):1409–1425
Kuman K, Li C, Li H (2014) Large cutting tools in the Danjiangkou Reservoir Region, central China. J Hum Evol 76:129–153
Lee HW (2017) Late-occurring handaxes in Korea: their homogeneity and variability. Archaeologies 13:489–534
Lepre CJ, Roche H, Kent DV et al. (2011) An earlier origin for the Acheulian. Nature 477:82–85
Li Y, Zhou Y, Sun X et al. (2018) New evidence of a lithic assemblage containing in situ Late Pleistocene bifaces from the Houfang site in the Hanshui River Valley, Central China. C R Palevol 17(1–2):131–142
Lycett SJ (2009) Understanding ancient hominin dispersals using artefact data: a phylogeographic analysis of Acheulean handaxes. PLOS ONE 4(1):e7404
Lycett SJ, Gowlett JAJ (2008) On questions surrounding the Acheulean ‘tradition’. World Archit 40(3):295–315
Lycett SJ, von Cramon-Taubadel N (2008) Acheulean variability and hominin dispersals: a model-bound approach. J Archaeol Sci 35(3):553–562
Lycett SJ, Bae CJ (2010) The Movius Line controversy: the state of the debate. World Archit 42(4):521–544
Lycett SJ, Norton CJ (2010) A demographic model for Palaeolithic technological evolution: the case of East Asia and the Movius Line. Quat Int 211:55–65
Lycett SJ, von Cramon-Taubadel N (2015) Toward a “quantitative genetic” approach to lithic variation. J Archaeol Method Theory 22:646–675
Malinsky-Buller A (2016) The muddle in the Middle Pleistocene: the Lower–Middle Palaeolithic transition from the Levantine perspective. J World Prehist 29:1–78
Marshall CR (2010) Using confidence intervals to quantify the uncertainty in the end-points of stratigraphic ranges. Paleontol Soc Pap 16:291–316
Marzke M (2013) Tool making, hand morphology and fossil hominins. Philos Trans R Soc B 368(1630):20120414
Mathias C, Bourguignon L, Brenet M et al. (2020) Between new and inherited technical behaviours: a case study from the Early Middle Palaeolithic of Southern France. Archaeol Anthropol Sci 12:146
McBrearty S, Tryon C (2006) From Acheulean to Middle Stone Age in the Kapthurin Formation, Kenya. In: Hovers E, Kuhn SL (eds) Transitions before the transition: evolution and stability in the Middle Paleolithic and Middle Stone Age. Springer, New York, pp. 257–278
McNabb J, Binyon F, Hazelwood L (2004) The large cutting tools from the South African Acheulean and the question of social traditions. Current Anthropol 45(5):653–677
Meignen L, Bar-Yosef O (2020) Acheulo-Yabrudian and Early Middle Paleolithic at Hayonim Cave (Western Galilee, Israel): continuity or break? J Hum Evol 139:102733
Méndez-Quintas E, Demuro M, Arnold LJ et al. (2019) Insights into the late stages of the Acheulean technocomplex of Western Iberia from the Arbo site (Galicia, Spain). J Archaeol Sci: Rep 27:101934
Méndez-Quintas E, Santonja M, Arnold LJ et al. (2020) The Acheulean technocomplex of the Iberian Atlantic margin as an example of technology continuity through the Middle Pleistocene. J Paleolit Archaeol https://doi.org/10.1007/s41982-020-00057-2
Michel V, Shen G, Valensi P et al. (2009) ESR dating of dental enamel from Middle Palaeolithic levels at Lazaret Cave, France. Quat Geochronol 4(3):233–240
Moncel MH, Ashton N (2018) From 800 to 500 Ka in Western Europe. The Oldest evidence of Acheulean in their technological, chronological, and geographic framework. In: Gallotti R, Mussi M (eds) The emergence of the Acheulean in East Africa and beyond: contributions in Honor of Jean Chavaillon. Springer, Cham, pp. 215–235
Moncel MH, Ashton N, Arzarello M et al. (2020a) Early Levallois core technology between Marine Isotope Stage 12 and 9 in Western Europe. J Hum Evol 139:102735
Moncel MH, Biddittu I, Manzi G et al. (2020b) Emergence of regional cultural traditions during the Lower Palaeolithic: the case of Frosinone-Ceptano basin (Central Italy) at the MIS 11-10 transition. Archaeol Anthropol Sci 12:185
Niewoehner WA (2006) Neanderthal hands in their proper perspective. In: Hublin J-J, Harvati K, Harrison T (eds) Neanderthals revisited: new approaches and perspectives. Springer, Dordrecht, pp. 157–190
Norton CJ, Bae K, Harris JWK et al. (2006) Middle Pleistocene handaxes from the Korean Peninsula. J Hum Evol 51(5):527–536
Norton CJ, Gao X, Feng X (2009) The East Asian Middle Paleolithic reexamined. In: Camps M, Chauhan PR (eds) Sourcebook of Palaeolithic transitions. Springer, New York, pp. 245–254
O’Brien MJ, Buchanan B (2017) Cultural learning and the Clovis colonization of North America. Evol Anthropol 26(6):270–284
Otte M (2019) Cognitive capabilities of the Neanderthals. In: Nishiaki Y, Joris O (eds) Learning among Neanderthals and Palaeolithic modern humans. Springer, New York, pp. 35–55
Pappu S, Akhilesh K (2019) Tools, trails and time: debating Acheulian group size at Attirampakkam, India. J Hum Evol 130:109–125
Pearse WD, Davis CC, Inouye DW et al. (2017) A statistical estimator for determining the limits of contemporary and historic phenology. Nat Ecol Evol 1:1876–1882
Petraglia MD, Ditchfield P, Jones S et al. (2012) The Toba volcanic super-eruption, environmental change, and hominin occupation history in India over the last 140,000 years. Quat Int 258:119–134
Picin A (2017) Technological adaption and the emergence of Levallois in Central Europe: new insights from the Markkleeberg and Zwochau open-air sites in Germany. J Quat Sci 33(3):300–312
Porat N, Chazan M, Schwarcz H et al. (2002) Timing of the Lower to Middle Palaeolithic boundary: new dates from the Levant. J Hum Evol 43(1):107–122
Porat N, Chazan M, Grun R et al. (2010) New radiometric ages for the Fauresmith industry from Kathu Pan, southern Africa: implications for the Earlier to Middle Stone Age transition. J Archeol Sci 37(2):269–283
Rivadeneira MM, Hunt G, Roy K (2009) The use of sighting records to infer species extinctions: an evaluation of different methods. Ecology 90:1291–300
Rizal Y, Westaway KE, Zaim Y et al. (2020) Last appearance of Homo erectus at Ngandong, Java, 117,000-108,000 years ago. Nature 577:381–385
Roberts DL, Solow AR (2003) When did the dodo become extinct? Nature 426:245
Rogers AR, Harris NS, Achenbach AA (2020) Neanderthal-Denisovan ancestors interbred with a distantly related hominin. Science 6(8):eaay5483
Sanchez-Yustos P, Diez-Martin F, Dominguez-Rodrigo M et al. (2018) Acheulean without handaxes? Assemblage variability at FLK West (Lowermost Bed II, Olduvai, Tanzania). J Anthropol Sci 96:1–22
Santonja M, Perez-Gonzalez A, Panera J et al. (2016) The coexistence of Acheulean and ancient Middle Palaeolithic techno-complexes in the Middle Pleistocene of the Iberian Peninsula. Quat Int 411(Part B):367–377
Scerri EML, Shipton C, Clark-Balzan L et al. (2018) The expansion of later Acheulean hominins into the Arabian Peninsula. Sci Rep 8:17165
Scerri EML, Chikhi L, Thomas MG (2019) Beyond multiregional and simple out-of-Africa models of human evolution. Nat Ecol Evol 3:1370–1372
Sharon G, Alperson-Afil N, Goren-Inbar N (2011) Cultural conservatism and variability in the Acheulian sequence of Gesher Benot Ya’aqov. J Hum Evol 60(4):387–397
Shea JJ (2017) Stone tools in human evolution. Cambridge University Press, New York
Shimelmitz R, Kuhn SL (2018) The toolkit in the core: there is more to Levallois production than predetermination. Quat Int 464(Part A):81–91
Shipton C (2016) Hierarchical organization in the Acheulean to Middle Palaeolithic transition at Bhimbetka, India. Camb Archeol J 26(4):601–618
Shipton C (2020) The unity of Acheulean culture. In: Groucutt HS (ed) Culture history and convergent evolution. Springer, Cham, pp. 13–27
Shipton C, Petraglia MD (2011) Inter-continental variation in Acheulean bifaces. In: Norton CJ, Braun DR (eds) Asian paleoanthropology. Springer, Cham, pp. 49–55
Solow AR (1993) Inferring extinction from sighting data. Ecology 74:962–64
Solow AR (2005) Inferring extinction from a sighting record. Math Biosci 195:47–55
Stout D (2011) Stone toolmaking and the evolution of human culture and cognition. Philos Trans R Soc B 366:1050–1059
Tocheri MW, Orr CM, Jacofsky MC et al. (2008) The evolutionary history of the hominin hand since the last common ancestor of Pan and Homo. J Anat 212(4):544–562
de la Torre I, Mora R (2014) The transition to the Acheulean in East Africa: an assessment of paradigms and evidence from Olduvai Gorge. J Archaeol Method Theory 21:781–823
de la Torre I (2016) The origins of the Acheulean: past and present perspectives on a major transition in human evolution. Philos Trans R Soc B 371(1698):20150245
Uomini NT, Meyer GF (2013) Shared brain lateralization patterns in Language and Acheulean stone tool production: a functional transcranial doppler ultrasound study. PLoS ONE 8(8):e72693
Villa P (2009) The Lower to Middle Paleolithic transition. In: Camps M, Chauhan PR (eds) Sourcebook of Palaeolithic transitions. Springer, New York, pp. 265–270
Villanea FA, Schraiber JG (2019) Multiple episodes of interbreeding between Neanderthal and modern humans. Nat Ecol Evo 3:39–44
Vinh LS, von Haeseler A (2004) IQPNNI: moving fast through tree space and stopping in time. Mol Biol Evol 21:1565–1571
Wood B, Boyle EK (2016) Hominin taxic diversity: fact or fantasy? Yearb Phys Anthropol 159:S37–S78
Wynn T, Gowlett J (2018) The handaxe reconsidered. Evol Anthropol 27(1):21–29
Zaidner Y, Weinstein-Evron M (2020) The emergence of the Levallois technology in the Levant: a view from the Early Middle Paleolithic site of Misliya Cave, Israel. J Hum Evol 144:102785
Acknowledgements
I.J. was supported by J.E. Purkyně Fellowship of the Czech Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Key, A.J.M., Jarić, I. & Roberts, D.L. Modelling the end of the Acheulean at global and continental levels suggests widespread persistence into the Middle Palaeolithic. Humanit Soc Sci Commun 8, 55 (2021). https://doi.org/10.1057/s41599-021-00735-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1057/s41599-021-00735-8
This article is cited by
-
Ecosystem productivity affected the spatiotemporal disappearance of Neanderthals in Iberia
Nature Ecology & Evolution (2022)
