
Abstract
The squash family (Cucurbitaceae) contains some of the most important crops cultivated worldwide and has played an important ecological, economic, and cultural role for millennia. In the American tropics, squashes were among the first cultivated crop species, but little is known about how their domestication unfolded. Here, we employ direct radiocarbon dating and morphological analyses of desiccated cucurbit seeds, rinds, and stems from El Gigante Rockshelter in Honduras to reconstruct human practices of selection and cultivation of Lagenaria siceraria, Cucurbita pepo, and Cucurbita moschata. Direct radiocarbon dating indicates that humans started using Lagenaria and wild Cucurbita starting ~ 10,950 calendar years before present (cal B.P.), primarily as watertight vessels and possibly as cooking and drinking containers. A rind directly dated to 11,150–10,765 cal B.P. represents the oldest known bottle gourd in the Americas. Domesticated C. moschata subsequently appeared ~ 4035 cal B.P., followed by domesticated C. pepo ~ 2190 cal B.P. associated with increasing evidence for their use as food crops. Multivariate statistical analysis of seed size and shape show that the archaeological C. pepo assemblage exhibits significant variability, representing at least three varieties: one similar to present-day zucchini, another like present-day vegetable marrow, and a native cultivar without modern analogs. Our archaeobotanical data supports the hypothesis that Indigenous cucurbit use started in the Early Holocene, and that agricultural complexity during the Late Holocene involved selective breeding that encouraged crop diversification.
Similar content being viewed by others


Introduction
The squash family (Cucurbitaceae) consists of nearly a thousand species worldwide, all of great ecological, economic and cultural importance1. At least six cucurbit species were domesticated in the Americas and are currently cultivated globally for their edible fruit and seeds1. Lagenaria siceraria (Molina) Standl. (bottle gourd), Cucurbita pepo L. (summer squash, acorn squash, jack-o-lantern pumpkins), Cucurbita argyrosperma K. Koch (cushaw), Cucurbita moschata Duchesne (butternut squash, commercial pie pumpkins), Cucurbita ficifolia Bouché (chilacayote, fig-leaf gourd), and Cucurbita maxima Duchesne (hubbard squashes, pumpkins). These cultivated cucurbits originated from different wild ancestors in different regions and constitute an important part of various cultural traditions across the Americas. Cultivated cucurbits are broadly characterized by large fruit sizes, variability in fruit shape and color, increased sugar and carotenoid content, decreased bitterness, and dominant upright growth2.
Archaeological evidence shows that cucurbits were among the first field crops cultivated in the Americas3,4. L. siceraria is native to Africa, but it arrived in the Americas during the Late Pleistocene and was likely domesticated independently in Eurasia, Africa, and the Americas3,4,5. Bottle gourds were widely used in the Americas during the Early Holocene and archaeological remains have been found in early archaeological contexts ranging from Florida (~ 10,000 cal B.P.), Mexico (~ 8000 cal B.P.), and Panama (~ 8300 cal B.P.) to Ecuador (9300 BP) and Peru (9000–5500 cal B.P.)3,6,7,8.
Cucurbita pepo was separately domesticated in at least two different regions, leading to distinct subspecies: C. pepo subsp. ovifera in eastern North America and C. pepo L. subsp. pepo in northeastern Mexico9,10. The initial domestication of C. pepo might have taken place near Guilá Naquitz and the Tehuacán valley (~ 10,000 cal B.P.) and subsequently diffused to central (7920 cal B.P.) and northern Mexico (6310 cal B.P.)3,11,12,13,14,15. C. argyrosperma was domesticated from C. argyrosperma subsp. sororia and its putative center of origin is in central and southern Mexico16. Phytolith data suggests the presence of a domesticated cucurbit, possibly C. argyrosperma, in the central Balsas River valley of Mexico as early as 8700 cal B.P.17. The earliest unambiguous archaeological specimen of C. argyrosperma is a ~ 5100 cal B.P. peduncle from the Ocampo caves in northeastern Mexico11. Other archaeological and archaeogenomic evidence reveals that following C. argyrosperma’s domestication, it diffused northeast of Mexico into the eastern and central portions of the United States before European contact4,15,18,19. For instance, C. argyrosperma’s seeds have been recovered from Late Mississippian contexts in Arkansas and it has been suggested that this cultivar was established in the region ~ 1310–623 cal B.P.18.
Cucurbita maxima was domesticated in the Amazonian tropical lowlands10,20. C. ficifolia and C. moschata were also likely domesticated in South America, but neither their wild ancestors nor centers of origin are currently well defined10,16,20. C. moschata appears to have arrived in Mexico from farther south as suggested by the identification of distinctive phytoliths dated between 9240 cal B.P. in Panama and desiccated macrobotanical remains dated to 6970 cal B.P. in the northern coast of Peru7,21,22.
Advanced molecular research has resolved the major phylogenetic relationships among squash and gourd species. In addition, archaeobotanical approaches can provide tangible evidence of these crops’ movements across space and changes over time. For example, systematic morphometric analysis of C. pepo seeds has documented diachronic changes in seed traits and size that can be used to differentiate between wild and domesticated subspecies of C. pepo. Because of a lack of similar studies in other cucurbit species, it is unknown whether similar morphological changes associated with a domestication syndrome can be traced. The use of morphometric seed characters (e.g., length, width, and thickness) is further complicated by significant size overlap between wild and domesticated varieties and by seed deformation caused by preservation conditions, cooking, and carbonization23. These issues contribute to a persistent lack of certainty when identifying cucurbit seeds from archaeological contexts. In this paper, we address the question of how human selection shaped domesticated Cucurbita and Lagenaria by conducting a quantitative analysis of desiccated seeds, rinds, and stems from the well-dated chronological sequence of El Gigante, a dry rock shelter located in south-central Honduras. We employ direct radiocarbon dating and a range of multivariate quantitative methods to identify possible changes in size and shape of seeds and rinds to distinguish between wild and domesticated cucurbits over time. The goal of this approach was to fundamentally improve our understanding of the tempo and mode of the domestication of these plant cultivars.
El Gigante Rockshelter
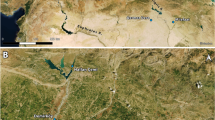
El Gigante (14° 13ʹ N 88° 03ʹ W, 1300 m above sea level) is a large rock shelter located along the western ridge of the Estanzuela River in the highlands of western Honduras (Fig. 1). The site was excavated in 2000 and 200124,25. The occupation at the site spans great time depth with deposits dating from the Terminal Pleistocene through the Postclassic/Colonial periods. Bayesian chronological modeling of radiocarbon dates from the rock shelter suggest 16 phases of occupation26. The initial occupation took place during the Early and Middle Esperanza phases between 10,985–10,705 and 10,705–10,210 cal B.P., respectively, and was characterized by the presence of stemmed bifaces and avocado, squash and maguey remains. After a gap of 190 years, the Late Esperanza (10,020–9,520 cal B.P.) featured wide fluted projectile points as well as ciruela (Spondias), squash, maguey, and avocado remains27. The subsequent phase of occupation, Marcala, was episodic. The Early Archaic (Early Marcala 1: 8945–8520 cal B.P., Early Marcala 2: 8090–7865 cal B.P.) and Middle Archaic (Middle Marcala 1: 7570–7265 cal B.P., Middle Marcala 2: 7140–6960 cal B.P., and Middle Marcala 3: 6665–6365 cal B.P.) habitations are characterized by the abundance of ciruela, avocado, squash, and maguey remains. A long occupation hiatus took place between 6365 and 4440 cal B.P. The Late Marcala phase (4400–4025 cal B.P.) corresponding to the regional Late Archaic is characterized by the appearance of maize (Zea mays), which is found along with tree fruits, including ciruela and avocado as well as a low abundance of squash. Following a second hiatus of 530 years, the Early and Middle Formative occupation (Early Estanzuela phase: 3500–3185 cal B.P.; Middle Estanzuela: 2985–2470 cal B.P.) features the appearance of ceramics. The Late Formative occupation (Late Estanzuela phase 1: 2225–2080 cal B.P.; Late Estanzuela phase 2: 1980–1810 cal B.P.) contains large quantities of maize and the appearance of beans (Phaseolus spp.). During the Classic period (Early Classic: 1690–1475 cal B.P., Late Classic: 1295–865 cal B.P.), maize continues to constitute a main component of the assemblage, and avocados, squash, and wild fruits increase in abundance. The occupation of the rock shelter lasted through the Postclassic/Colonial eras (545–310 cal B.P.), periods when the site was rarely used. During the most recent occupations, acorns were common, but maize and gourds were rare in the assemblage.

(a) Location of El Gigante Rockshelter and archeological sites with Cucurbita measured seeds used in the analysis. (b) Percentages of macrobotanical remains of Cucurbita and Lagenaria and other field crops from El Gigante Rockshelter, Honduras.
Results
Direct radiocarbon dating
Direct accelerated mass spectrometry (AMS) radiocarbon (14C) dating of desiccated cucurbit rinds, stems, and seeds throughout the 11,000-year occupation of El Gigante has revealed a long history of cucurbit use by humans (Table 1). The earliest analyzed Cucurbita rinds date from the Esperanza phase and were directly AMS 14C dated to 11,090–10,720 (2σ; PSUAMS-8055) cal B.P. and 10,655–10,425 (2σ; PSUAMS-8251) cal B.P. Two Early Marcala-phase Cucurbita rinds dated to 8595–8430 (2σ; PSUAMS-8064) cal B.P. and 8020–7880 (2σ; PSUAMS-8056) cal B.P., while two Middle Marcala rinds dated to 6635–6485 (2σ; PSUAMS-8063) cal B.P. and 6495–6395 (2σ; PSUAMS-8062) cal B.P. One Late Marcala Cucurbita moschata stem was dated to 4145–3930 (2σ; PSUAMS-6129) cal B.P. We recovered Cucurbita seeds only from the Estanzuela (Formative) and Classic-period deposits. One C. pepo seed resulted in a date of 2305–2060 (2σ; PSUAMS-6128) cal B.P., and a C. moschata seed yielded a date of 1245–1065 (2σ; PSUAMS-6126) cal B.P.
Lageneria remains were found from the Esperanza to the Colonial occupation phases. The earliest Lagenaria rind, from the Early Esperanza phase, was directly dated to 11,150–10,765 (2σ; PSUAMS-8058) cal B.P. Rinds from Archaic deposits returned results of 8020–7935 (2σ; PSUAMS-8059; Early Marcala 2) cal B.P. and 4155–3985 (2σ; PSUAMS-8061; Late Marcala) cal B.P. Late Estanzuela Lagenaria rinds yielded dates between 2290 and 2000 (2σ; Beta-316169, PSUAMS-8060 and PSUAMS-8454) cal B.P.
Rind fragments
Based on their morphological attributes (14), we identified a total of 448 rind fragments (weighing 16.5 g) of Cucurbita and 387 rind fragments (weighing 56.3 g) of Lagenaria. Cucurbita rind fragments were found in all occupation phases (i.e., Esperanza, Marcala, Estanzuela, Classic, and Postclassic/Colonial). The vast majority of Lagenaria rinds were recovered from contexts associated with the Estanzuela (38%) and Classic (53%) occupation phases, while few rinds were recovered from the Esperanza (6%) and Marcala (2%) phases.
Thickness of Cucurbita rind fragments increase significantly over time (F12,151 = 11.65, P < 0.0001) (Fig. 2a). The thickness of Lagenaria rinds, however, showed little variation over time (F14,284 = 1.53, P = 0.12) (Fig. 2b). However, given the unequal sample size between occupation phases, results should be interpreted with caution. Evidence of post-harvest modifications in rinds of Cucurbita is notable, particularly thermal alteration during the Esperanza (10,990–9500 year B.P.) and Marcala (8950–4020 year B.P.) phases. Specifically, a large proportion of Cucurbita rind fragments recovered from the Esperanza (50%) and Marcala (66.7%) phases were burned. In contrast, only a few of the rinds from the later Estanzuela (3500–1810 year B.P.; 5%) and Postclassic/Colonial (545–310 year B.P.; 11.1%) phases showed evidence of burning. One small Cucurbita rind fragment recovered from Unit 12, Level 4 was colored red (Fig. 3a). We found that only 3.6% of the rinds of Lagenaria had evidence of abrasion. A few Lagenaria rinds (n = 5) were polished on one side. These rinds yielded AMS dates between 2120 and 320 cal B.P. (2σ; PSUAMS-8450, -8451, -8452, -8453 and -8454). For example, one rind fragment recovered from Unit 11, level 2 was polished on one side with evidence of deliberate holes and included attached to fragments of cordage (Fig. 3b).

Variation of rind thickness of (a) Cucurbita and (b) Lagenaria recovered across occupational phases from El Gigante Rockshelter, Honduras. Different letters indicate significant differences (P < 0.05). The sample size is indicated below each boxplot.

Cucurbit desiccated remains recovered from El Gigante Rockshelter, Honduras. (a) Cucurbita rind stained with red pigment. (b) Polished Lageneria rind with attached rope. (c) Cucurbita pepo peduncle. (d) Cucurbita moschata peduncle (PSUAMS-6129, 4035 cal B.P.). (e) Lagenaria siceraria seed. (f) C. pepo seed. (g) C. moschata seed.
Peduncles
A total of 16 Cucurbita peduncles was recovered, of which seven were identified as C. pepo and seven as C. moschata (following 14, 15, 49, 50). C. pepo stems exhibited seven to nine major ridges that had a mean basal diameter of 19.7 ± 0.5 mm (Fig. 3c). C. moschata peduncles were characterized by the presence of five major ridges, a pentagonal base (Fig. 3d), and a mean basal diameter of 13.9 ± 1.7 mm.
Seeds
A total of 71 ancient cucurbit seeds were recovered from El Gigante. Most of these seeds originate from Late Estanzuela, later, or mixed contexts. Based on their external morphology, we identified these seeds as C. pepo (n = 48), C. cf. pepo (n = 3), C. moschata (n = 5), Cucurbita sp. (n = 6), and Lagenaria (n = 9) (Fig. 3f,g).
A hierarchical cluster analysis comparing previously published paleontological and archaeological C. pepo seeds from sites throughout Central and North America (Fig. 1) generated two main morphological groups, which can be subdivided in five additional subgroups (Fig. 4). Cluster 1 includes the largest seeds. Subcluster 1a includes the longest (17.9 ± 1.4 cm) and widest seeds (11.5 ± 1.3 cm), while subcluster 1b comprises slightly shorter (15.2 ± 1.5 cm) and narrower (9.6 ± 0.2 cm) seeds. In contrast, cluster 2 includes the smallest seeds, from which subcluster 2a (length: 10.7 ± 1.4 cm, width: 6.7 ± 0.8 cm) includes the shortest and narrowest seeds in comparison to subcluster 2b (length: 10.8 ± 1.6 cm, width: 7.1 ± 1.0 cm) and subcluster 2c (length: 13.4 ± 2.6 cm, width: 7.3 ± 1.3 cm). The El Gigante archaeological C. pepo seeds fell either in subcluster 1b (n = 39) or subcluster 2a (n = 9) suggesting the presence of at least two phenotypes (Supplementary Tables S2, S3).

Hierarchical cluster analysis of paleontological and archaeological Cucurbita pepo seeds using Ward’s agglomerative hierarchical classification based on maximum length, maximum width, and length/width ratio. Seeds recovered from El Gigante (EG) are highlighted in red. BRO Brown28, CHM Mount 51 Cahokia—Chmurny’s type 129, CLO Cloudsplitter30, GP Green Point31, HAY Haystack30, HOO Hoonton Island5, HOX Hoxie site32, KIN King Coulee33, NKA Newt Kash Hollow30, OZ-AG Ozarks—Agnew Small, Beaver Pond and Whitney Bluff19, PA-L Page-Ladson, PHS Phillips Spring34,35, RHO Rhoads36, RO Rogers30, ROM Romeros cave12, SAG Saginaw37, SC Salts Cave34, UTZ Utz38, VAL Valenzuelas Cave12.
To better understand the variability in the archaeological assemblage, we compared the archaeological C. pepo seeds from El Gigante to a dataset of modern wild and domesticated C. pepo seeds that we measured (Supplementary Table S1). Principle component analysis (PCA) of morphometric traits allow us to separate modern variants of domesticated C. pepo seeds from those of wild C. pepo (C. pepo var. fraterna, C. pepo subsp. texana and C. pepo var. ozarkana) (Fig. 5a). The first two components collectively explain 78% of the variance in the data (61.5% and 16.8%, sequentially). Principal component 1 discriminated between total seed length and length to width ratio, while principal component 2 separated seeds by maximum width and seed margin width. Measurements of archaeological seeds recovered from El Gigante demonstrate that most cucurbit seeds are comparable in size to modern domesticated C. pepo seeds. Only six of the El Gigante seeds are sufficiently small to fall within the confidence interval ranges of wild C. pepo seeds and specifically, the varieties C. pepo var. ozarkana and C. pepo var. fraterna.

Principal component analysis biplots comparing size of archaeological seeds from El Gigante Rockshelter with modern seeds in (a) Cucurbita pepo; (b) C. pepo varieties39; (c) Cucurbita moschata; and (d) Lagenaria siceraria based on morphometric measurements.
To evaluate if the ancient C. pepo seeds recovered from El Gigante are similar to modern varieties of domesticated and wild C. pepo, we performed a second PCA using the cucurbit cultivar measurement data reported in Ref.40. Principal component 1 (63% of total variance) discriminated among seeds by total length and maximum width whereas principal component 2 (35.2% of total variance) separated seeds according to their length to width ratio (Fig. 5b). The biplot shows high variability within the El Gigante archaeological seeds and the fact that the first two components captured 98% of the variance attests to the strength of the relationships among the cases and variables. Ten archaeological seeds were morphometrically similar to modern acorn, scallop and crookneck varieties of wild C. pepo subsp. texana. Sixteen seeds overlap with modern seeds of cocozelle, vegetable marrow and zucchini varieties of C. pepo subsp. pepo. Finally, 17 of the archaeological seeds are not associated with any variety of modern C. pepo in particular.
A multivariate analysis shows that only one archaeological seed of C. moschata from El Gigante is morphometrically similar to its modern equivalent (Fig. 5c). Principal component 1, which accounts for 43.6% of the variance, separates seeds by total length, maximum width, and mass. Principal component 2, accounting for 30.3% of the variance, separates seeds by surface area and length to width ratio. Three of four archaeological seeds fell outside the confidence intervals of modern C. moschata. Archaeological seeds have low principal component 1 scores, which are reflective of small seeds (small length and small width), but high principal component 2 scores, indicating that they have a broader surface area than most of the modern seeds.
A final PCA comparing ancient and modern L. siceraria seeds shows that the first component (62.8% of the variance) differentiates seed surface area, total length, and mass. The second component (26.8% of the variance) is related to maximum width and length to width ratio (Fig. 5d). Most of the El Gigante Lagenaria seeds are distributed along the left side of the first component, showing their smaller size in relation to modern seeds.
To assess shape variation, we calculated elliptical Fourier descriptors (EFDs) (Fig. 6). The results largely show that Cucurbita species are clearly discriminated from Lagenaria siceraria on the LDA1-LDA2 biplot (94% of the total variation). Furthermore, the LDA discriminates the seeds of domesticated C. pepo from the wild C. pepo species (C. pepo var. fraterna, C. pepo var. ozarkana, and C. pepo subsp. texana). It is notable, however, that there is significant overlap in seed shape between C. pepo and C. moschata and between C. pepo var. ozarkana and C. moschata.

Linear discriminant analysis (LDA) biplot of axes 1 and 2 (77.1% and 13.7% of the total variance, respectively) showing the morphological gradient of modern Cucurbita seeds in relation to Lagenaria siceraria according to the elliptic Fourier transform (EFT) method. Seed outlines represent approximate seed shape.
The geometric morphometric analysis indicates that the shape of the archaeological gourd and squash seeds from El Gigante vary considerably. Of the 21 archaeological seeds morphologically identified as C. pepo, the elliptical Fourier transform (EFT) method correctly classified only three specimens as C. pepo. The remaining archaeological seeds were allocated to the wild C. pepo group (seven as C. pepo var. ozarkana, eight as C. pepo var. texana, and three as C. pepo var. fraterna). Only one of the archaeological C. moschata seeds was correctly classified as C. moschata, while the other two seeds were attributed to the C. pepo group. Finally, most of the archaeological Lagenaria seeds (n = 5) were correctly classified as L. siceraria, but three were assigned as C. pepo and one as C. pepo var. ozarkana. In light of the variability inherent in cucurbit seeds, this method has limited power to add to morphological classification in this case.
Discussion
The chronology, size, and geometric morphometric analyses of Cucurbita and Lagenaria macrobotanical remains from El Gigante Rockshelter reveal intriguing patterns related to human reliance, use, and cultivation of cucurbits for food and technology during the last ~ 11,000 years. Key changes in plant selection and use correspond with broader subsistence and settlement changes spanning the Esperanza and Marcala phases, a time span that represents the long transition from plant foraging to low-level cultivation.
Ancient people inhabiting the area around El Gigante consistently used L. siceraria and Cucurbita through the entire sequence of occupation of the site. Initially, Cucurbita rinds likely gathered from the wild were significantly thinner than rinds recovered from subsequent periods when cultivated varieties were probably utilized. Research has shown that an increase in rind thickness is associated with the presence of domesticated squash in contrast to the brittle rinds of wild squash forms12. El Gigante Cucurbita rinds recovered from the earliest occupation levels were < 2 mm thick, suggesting that humans used wild Cucurbita species during this time. In contrast, Cucurbita rind thickness increases during the Late Archaic (2.5 mm, range 0.5–5.1), indicating the introduction of domesticated Cucurbita to the region ~ 4000 years ago. A similar change in rind thickness over time has been documented in the Guilá Naquitz cave, where Cucurbita rinds showed an increase in thickness from 0.84 mm (range 0.5–1.6 mm) to 1.15 mm (range 0.5–2.0 mm) between ca. 10,500 and 8500 years ago12. In contrast, rinds of C. maxima recovered from the Pampa Grande archaeological site in Argentina show high variation in thickness through time (mean 4.09 mm, range 2.08–6.64 mm), suggesting that different varieties were cultivated and managed for different purposes (e.g., some as food and others as containers)41,42.
Given that a large percentage of Cucurbita rinds from these earlier occupations were burned, it seems likely that mobile hunter-gatherers used some cucurbits as cooking containers. We also suggest that some of these early, thinner cucurbits were roasted directly in proximity to cooking fires for consumption, potentially focusing on the squashes that had a lower concentration of bitter cucurbitacin than the rest of the fruits. In contrast, likely cultivated Cucurbita with more durable and thicker rinds could have been used to store food or water. The recovery of a Cucurbita rind stained with red pigment suggests that squash containers also may have been decorated vessels42,43.
The recovery of Lagenaria rinds and seeds spanning the occupation of El Gigante Rockshelter provides further evidence that gourds were used consistently during the early Holocene in Mesoamerica. AMS dating of a Lagenaria rind recovered from Unit 18, level 39 corresponding to the early Esperanza phase produced a date of 10,935 cal B.P. (11,150–10,765 cal B.P.). The date is considerably older than the most ancient specimens of bottle gourd previously reported, which were from Guilá Naquitz, Oaxaca (10,021 ± 105 cal B.P.; 10,198–9781 cal year BP) and Little Salt Spring, Florida (10,015 cal B.P.)8,44. Lagenaria was likely one of the first field crops in El Gigante and the light-weight fruits were used as containers even after ceramic technology was introduced to the region during the Early Formative period (~ 3600 cal B.P.).
Lagenaria rinds show little change in thickness over time. It is notable that El Gigante Lagenaria rinds exhibit a thickness (1.9 ± 1 mm) that overlaps with rinds of wild African L. siceraria (< 2.0 mm)3. El Gigante rinds are also consistently thinner than those recovered from other archaeological contexts in the Americas, including the Ocampo (3.2 mm), the Guilá Naquitz (4.6 mm), and the Coxcatlán (3.8 mm) caves in Mexico, and Ancón (4.8 mm) in Peru3. The domestication status of the recovered Lagenaria remains to be assessed in detail due to the absence of morphological characterizations in wild and domesticated Lagenaria species. These results highlight the importance of conducting further comparative studies of modern and archaeological plant remains alongside paleogenomic analyses to assess the domestication syndrome in Lagenaria and its possible center of domestication in south-central Mesoamerica.
Morphometric studies of variation in seed size improve our understanding of how domestication shaped plant diversity45. Refined methods of morphometric analysis are critical if we are to fully understand the processes involved in the domestication and spread of these cultigens. Geometric morphometric analysis is a complementary approach that can be used to assess changes in seed shape that may be associated with domestication. This method has been useful in the analysis of several crop species, including olives, grapes, barley, and date palms46,47,48,49. For instance, seed morphology has been used to distinguish between pips of wild and domesticated grape (Vitis vinifera) varieties in Italy50. Our results indicate that domesticated C. pepo show discernable differentiation in seed size and shape from wild species (C. pepo var. fraterna, C. pepo subsp. texana, and C. pepo var. ozarkana). Furthermore, distinct groups of C. pepo and C. pepo visualized with PCA and EFDs confirm existing species distinctions based on morphological data. We argue that the high level of phenotypic variability in cultivated C. pepo is a consequence of varietal diversification and reflects the long-term history of cultivation associated with the selection of specific morphological traits, such as fruit size and shape. These morphological differences may be explained by divergent selection pressures. For instance, wild C. pepo species were subject to environmental constraints that seemed to have minimized seed size, while domesticated C. pepo were subject to ongoing human selection related to cultivation practices that shaped particular phenotypes.
In contrast to fruit size, it is likely that seed shape was not a direct target of human selection in C. pepo. The evolutionary role of seed shape in cucurbits is not well understood, but shape changes could be related to a pleiotropic effect that determines seed persistence in the soil, seed germination and dispersal51. Some empirical studies have shown that long-term persistent species tend to have spherical seeds, while transient species produce flattened and/or elongated seeds52,53. Genetic drift associated with the restricted gene pool of breeding can also have essentially random implications for non-selected traits such as seed shape.
The shape analysis was not able to differentiate seeds of C. moschata from subspecies of C. pepo. Modern seeds of C. moschata showed a high overlap with both the domesticated C. pepo and the wild C. pepo var. ozarkana. The results expose the limitations of using shape analysis for identifying C. moschata seeds and highlights the importance of including other qualitative characters for identification.
Multivariate morphometric analyses show that most of the C. pepo seeds from El Gigante fall within the size parameters defined for modern types of domesticated C. pepo seeds. Paris and Nerson39 have also shown a relationship between seed size and fruit size in C. pepo. Our analysis suggests that some of the archaeological seeds are similar to domesticated present-day varieties, including the cocozelle and zucchini varieties, which are characterized by long and uniformly cylindrical fruits, and the vegetable marrow variety, which is distinguished by short round fruits with a slightly broad apex39.
The seeds of C. pepo that are similar in size to wild C. pepo subspecies could have been the product of genetic exchange between crops and wild cucurbit species in the area. Gene flow and crop-to-wild hybridization have been reported to be common in C. pepo due to pollen transfer between plants by bees40,54, and hybrid plants have been shown to produce highly variable fruit sizes54.
In contrast to the morphometric results, shape analysis via linear discriminant analysis (LDA) allocated most of the El Gigante C. pepo seeds to wild forms. These results could suggest the presence of an unknown autochthonous variety or a separate subspecies that has since become extinct, for instance the as-yet-unknown wild progenitor of C. pepo ssp. pepo cultivated types. They also suggest that the region in which El Gigante is located may have been an important setting of squash diversification. A molecular analysis could help to better address squash diversity via aDNA analysis. However, the difficulty of differentiating wild from domesticated seeds in archaeological contexts could be associated with the preservation of the seed remains. Even though this analysis only included complete and well-preserved seeds, it is possible that the desiccation process might have altered the shape of some seeds.
Conclusions
The archaeobotanical record from El Gigante Rockshelter suggests early indigenous foragers utilized wild cucurbits for generations before the introduction of cultivated varieties. Identifications of maize pollen and microcharcoal particles recovered from wetlands in Lago Yojoa ~ 5400 cal B.P. and in the Copan Valley ~ 4300 cal B.P. suggests that the adoption of farming in Mesoamerica was a relatively early and protracted process55,56. In correspondence with these studies, domesticated butternut squash seems to have been initially introduced in El Gigante during the Late Marcala phase, between 4400 and 4025 cal B.P., yet the utilization of wild cucurbits and other tropical plants persisted over time. In fact, as indigenous farmers began to substantially modify their surrounding tropical ecosystems, they were able to successfully incorporate domesticated crops such as maize and squash while still relying on and likely managing other resources such as tree crops (e.g., Persea americana, Pouteria spp., and Spondias spp.) for their subsistence26.
The significance of El Gigante lies in both the length of its chronology and its extensive well-preserved plant assemblage. The rock shelter thus has great potential to both broaden and reshape our understanding of long-term human adaptations to domestication and early agriculture. The analytical techniques we used (digital morphometrics, multivariate statistics, and outline analysis based on EFDs) proved to be powerful tools for assessing variation in rind thickness and seed size and shape. Indeed, geometric morphometric approaches combined with traditional biometric methods of length and width measurements have allowed us to distinguish among cucurbit species, specifically domesticated C. pepo, wild C. pepo, and Lagenaria. We note, however, that the classification accuracy of C. moschata from C. pepo based on EFDs is more limited compared to qualitative morphological attributes.
The study of the plant remains recovered from El Gigante Rockshelter substantially contributes to improving our understanding of Indigenous human-plant coevolution and the dispersal of key cultigens57,58. As part of broader research, this paper presents an important piece of the tropical Americas domestication puzzle.
Materials and methods
Recovery of macrobotanical remains
El Gigante is one of six dry rock shelters in Mesoamerica that contain well-preserved macrobotanical assemblages that include both desiccated and carbonized remains57. The plant remains analyzed in this study were either hand-collected in situ during excavations or recovered through sieving (mesh size 4.5 mm and 500 μm) sediment from Unit 18. Well-preserved cucurbit (Cucurbita and Lagenaria) remains in the assemblage include rinds, peduncles, and seeds. Cucurbita rinds (n = 448) were distinguished from Lagenaria rinds (n = 387) based on diagnostic cross-section cell structure (Supplementary Fig. S1). Cucurbita rinds exhibit isodiametric cells with a regular configuration, a feature that enabled us to distinguish them from Lagenaria rinds12. Cucurbita peduncles were identified at the species level based on the shape of the peduncle. Wild Cucurbita presents relatively small peduncles (< 13 mm in diameter) with circular outline; domesticated C. pepo peduncles are relatively bigger (> 14 mm in diameter) and are characterized by the presence of alternating ten major bridges with an angular or pentagonal outline at the fruit attachment, while C. moschata peduncle presents five ridges, with regular furrows and flaring at the base12,13,59,60.
Direct radiocarbon dating
To further verify the age of the cucurbit assemblage, we conducted direct AMS 14C dating of 28 samples, including some of the earliest specimens in the assemblage. Samples were pretreated using conventional acid–base–acid methods and subsequently combusted, graphitized, and measured at the accelerated mass spectrometer of the Pennsylvania State University Radiocarbon Laboratory (PSUAMS). In addition, direct 14C dating of 310 additional (non-cucurbit) plant specimens allowed us to confidently assign the majority of cucurbit specimens to clarify chronological phases26.
Modern seed material
To assess differences in seed shape and size, we relied on modern seeds from the USDA Germplasm Resources Information Network (GRIN). Samples included accessions from wild Cucurbita species (C. pepo subsp. texana [n = 25], C. pepo var. ozarkana [n = 25], and C. pepo var. fraterna [n = 35]) and domesticated species (C. pepo [n = 100], C. moschata [n = 48], and L. siceraria [n = 33]) originating from Mexico, Guatemala, and the United States (Supplementary Table S4). We also relied on previously published measurements of additional varieties of domesticated subspecies and cultivar groups of C. pepo39.
Seed size and shape analysis
Digital images of the modern and archaeological seed samples were acquired using a digital camera attached to an Olympus SZX16 microscope with a 300-dpi resolution. Modern and archaeological seeds were photographed dorsally. Seeds were not photographed laterally because most of the archaeological seed coats were empty, resulting in lateral shape distortion. Digital images of the seeds were processed and measured using ImageJ v. 1.4261. The allometric characteristics of maximum length, maximum width, border thickness, and area were recorded in millimeters. Seeds were also weighed using a digital scale with a measuring error of 0.01 g. The full measurement datasets are included in Supplementary Tables S1 and S2.
Individual digital pictures were converted to binary images (i.e., seed image on a black background) in ImageJ. Each binary image was converted into chain code along the perimeter of each seed to create a harmonic series using SHAPE v. 1.362. Seed shape was quantified using outline analysis based on EFDs to assess for differences in overall seed shape within species. Chain code contours were posteriorly converted to normalized EFDs for Fourier analysis.
Data analysis
To assess differences in rind thickness between phases of occupation, we performed a one-way ANOVA for Lagenaria and Cucurbita, respectively. Differences in seed size between modern and archaeological seeds of C. pepo, C. pepo, C. moschata, and L. siceraria were assessed using a Principal Components Analysis (PCA). A second PCA was performed to compare similarities in seed size between seeds of different varieties of C. pepo from Paris and Nerson39 and archaeological seeds from El Gigante. A Ward’s agglomerative hierarchical classification using Euclidean distance was used to assess the morphological relationship of C. pepo seeds from El Gigante to other seeds recovered from paleontological and archaeological sites from North America (Supplementary Table S3). The cluster analysis was conducted using maximum length, maximum width, and length/width ratio. A Linear Discriminant Analysis (LDA) was performed to explore differences in seed shape between modern seeds of C. pepo, C. pepo, C. moschata, and L. siceraria. LDA is a statistical classification method used to evaluate shape variation of seeds by maximizing differences between predefined groups (i.e., species, subspecies and varieties) compared to intragroup variation.
Data availability
The datasets generated during the current study are provided in the Supplementary Information. The archeological remains studied in this research are curated in the Department of Anthropology (Pennsylvania State University).
References
Schaffer, A. A. & Paris, H. S. Melons, squashes, and gourds. In Encyclopedia of Food Sciences and Nutrition (ed. Caballero, B.) 3817–3826 (Academic Press, 2003).
Chomicki, G., Schaefer, H. & Renner, S. S. Origin and domestication of Cucurbitaceae crops: Insights from phylogenies, genomics and archaeology. N. Phytol. 226, 1240–1255 (2020).
Erickson, D. L., Smith, B. D., Clarke, A. C., Sandweiss, D. H. & Tuross, N. An Asian origin for a 10,000-year-old domesticated plant in the Americas. Proc. Natl. Acad. Sci. 102, 18315–18320 (2005).
Kistler, L. et al. Gourds and squashes (Cucurbita spp.) adapted to megafaunal extinction and ecological anachronism through domestication. Proc. Natl. Acad. Sci. 112, 15107–15112 (2015).
Decker, D. S. & Newsom, L. A. Numerical analysis of archaeological Cucurbita pepo seeds from Hontoon Island, Florida. J. Ethnobiol. 8, 35–44 (1988).
Piperno, D. R. Identifying crop plants with phytoliths (and starch grains) in Central and South America: A review and an update of the evidence. Quat. Int. 193, 146–159 (2009).
Dillehay, T. D., Rossen, J., Andres, T. C. & Williams, D. E. Preceramic adoption of peanut, squash, and cotton in northern Peru. Science 316, 1890–1893 (2007).
Kistler, L. et al. Transoceanic drift and the domestication of African bottle gourds in the Americas. Proc. Natl. Acad. Sci. 111, 1937–2941 (2014).
Paris, H. S. History of the cultivar-groups of Cucurbita pepo. Horticult. Rev. 25, 71–170 (2001).
Sanjur, O. I., Piperno, D. R., Andres, T. C. & Wessel-Beaver, L. Phylogenetic relationships among domesticated and wild species of Cucurbita (Cucurbitaceae) inferred from a mitochondrial gene: Implications for crop plant evolution and areas of origin. PNAS 99, 535–540 (2002).
Smith, B. D. Reconsidering the Ocampo caves and the era of incipient cultivation in Mesoamerica. Latin Am. Antiq. 8, 342–383 (1997).
Smith, B. D. The initial domestication of Cucurbita pepo in the Americas 10,000 years ago. Science 276, 932–934 (1997).
Smith, B. D. Seed size increase as a marker of domestication in squash. In Documenting Domestication (ed. Smith, B. D.) (University of California Press, 2006).
Smith, B. D. Eastern North America as an independent center of plant domestication. Proc. Natl. Acad. Sci. 103, 12223–12228 (2006).
Smith, B. D. Reassessing Coxcatlan cave and the early history of domesticated plants in Mesoamerica. Proc. Natl. Acad. Sci. 102, 9438–9445 (2005).
Kates, H. R., Soltis, P. S. & Soltis, D. E. Evolutionary and domestication history of Cucurbita (pumpkin and squash) species inferred from 44 nuclear loci. Mol. Phylogenet. Evol. 111, 98–109 (2017).
Piperno, D. R., Ranere, A. J., Holst, I., Iriarte, J. & Dickau, R. Starch grain and phytolith evidence for early ninth millennium B.P. maize from the Central Balsas River Valley, Mexico. Proc. Natl. Acad. Sci. 106, 5019–5024 (2009).
Fritz, G. J. Precolumbian Cucurbita argyrosperma spp. argyrosperma (Cucurbitaceae) in the eastern woodlands of North America. Econ. Bot. 48, 280–292 (1994).
Fritz, G. J. Prehistoric Ozark Agriculture: The University of Arkansas Rockshelter Collections (The University of North Carolina, 1986).
Lombardo, U. et al. Early Holocene crop cultivation and landscape modification in Amazonia. Nature 581, 190–193 (2020).
Piperno, D. R. & Dillehay, T. D. Starch grains on human teeth reveal early broad crop diet in northern Peru. PNAS 105, 19622–19627 (2008).
Piperno, D. R. The origins of plant cultivation and domestication in the new world tropics: Patterns, process, and new developments. Curr. Anthropol. 52, S453–S470 (2011).
Ucchesu, M. et al. Predictive method for correct identification of archaeological charred grape seeds: Support for advances in knowledge of grape domestication process. PLoS ONE 11, e0149814 (2016).
Scheffler, T. E., Hirth, K. G. & Hasemann, G. The El Gigante rockshelter: Preliminary observations on an early to late Holocene occupation in southern Honduras. Latin Am. Antiq. 23, 597–610 (2012).
Scheffler, T. E. The El Gigante Rock Shelter, Honduras (The Pennsylvania State University, 2008).
Kennett, D. J. et al. Trans-Holocene Bayesian chronology for tree and field crop use from El Gigante Rockshelter, Honduras. PLoS ONE 18, e0287195 (2023).
Iceland, H. B. & Hirth, K. G. The Paleoindian to archaic transition in central America: Esperanza phase projectile points recovered at the El Gigante rockshelter site, Honduras. In Preceramic Mesoamerica (eds Lohse, J. C. et al.) 259–277 (Routledge, 2021).
Blake, L. W. Cultivated plant remains from historic Indian and Osage Indian sites. In Plants from the Past (eds Blake, L. W. & Cutler, H. C.) 46–53 (University of Alabama Press, 2001).
Chmurny, W. W. The Ecology of the Middle-Mississippian Occupation of the American Bottom (University of Illinois at Urbana-Champaign, 1973).
Cowan, C. W. Evolutionary changes associated with the domestication of Cucurbita pepo: Evidence from eastern Kentucky. In People, Places, and Landscapes: Studies in Paleoethnobotany (ed. Gremillion, K. J.) 63–85 (University of Alabama Press, 1997).
Lovis, W. A. & Monaghan, G. W. Chronology and evolution of the green point flood plain and associated Cucurbita pepo. In Current Northeast Paleoethnobotany II Vol. 512 (eds Lovis, W. A. & Monaghan, G. W.) 141–150 (New York State Museum Albany, 2008).
Simon, M. L. Evidence for variability among squash seeds from the Hoxie site (11CK4), Illinois. J. Archaeol. Sci. 38, 2079–2093 (2011).
Perkl, B. E. Cucurbita pepo from King Coulee, Southeastern Minnesota. Am. Antiq. 63, 279–288 (1998).
King, F. B. Early cultivated cucurbits in eastern North America. In Prehistoric Food Production in North America Vol. 75 (ed. Ford, R. I.) 73–98 (Museum of Anthropology, University of Michigan, 1985).
Kay, M., King, F. B. & Robinson, C. K. Cucurbits from Phillips Spring: New evidence and interpretations. Am. Antiq. 45, 806–822 (1980).
Blake, L. W. & Cutler, C. Plant remains from the Rhoads site (11LO8), Illinois. In Plants from the Past (eds Blake, L. W. & Cutler, C.) 72–92 (University of Alabama Press, 1974).
Ozker, D. An Early Woodland Community at the Schultz Site 20SA2 in the Saginaw Valley and the Nature of the Early Woodland Adaptation in the Great Lakes Region Vol. 70 (University of Michigan Museum Anthro Archaeology, 1982).
Blake, L. W. & Cutler, H. Plant remains from the King Hill Site (23BN1) and comparisons with those from the Utz site (23SA2). Missouri Archaeol. 43, 86–110 (1982).
Paris, H. S. & Nerson, H. Seed dimensions in the subspecies and cultivar-groups of Cucurbita pepo. Genet. Resour. Crop Evol. 50, 615–625 (2003).
Pope, N. S. et al. The expansion of agriculture has shaped the recent evolutionary history of a specialized squash pollinator. Proc. Natl. Acad. Sci. 120, e2208116120 (2023).
Martínez, A. et al. Multidisciplinary studies in Cucurbita maxima (squash) domestication. Veg. Hist. Archaeobot. 27, 207–217 (2018).
Lema, V. S. Non-domesticated cultivation in the Andes: Plant management and nurturing in the Argentine northwest. Veg. Hist. Archaeobot. 24, 143–150 (2015).
Duncan, N. A., Pearsall, D. M. & Benfer, R. A. Gourd and squash artifacts yield starch grains of feasting foods from preceramic Peru. Proc. Natl. Acad. Sci. 106, 13202–13206 (2009).
Smith, B. D. Guilá Naquitz revisited: Agricultural origins in Oaxaca, Mexico. In Cultural Evolution: Contemporary Viewpoints (eds Feinman, G. M. & Mansanilla, L.) 15–60 (Kluwer Academic, 2000).
Spengler, R. N. & Mueller, N. G. Grazing animals drove domestication of grain crops. Nat. Plants 5, 656–662 (2019).
Terral, J.-F. et al. Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J. Biogeogr. 31, 63–77 (2004).
Terral, J.-F. et al. Insights into the historical biogeography of the date palm (Phoenix dactylifera L.) using geometric morphometry of modern and ancient seeds. J. Biogeogr. 39, 929–941 (2012).
Orrù, M., Grillo, O., Lovicu, G., Venora, G. & Bacchetta, G. Morphological characterization of Vitis vinifera L. seeds by image analysis and comparison with archaeological remains. Veg. Hist. Archaeobot. 22, 231–242 (2013).
Ros, J., Evin, A., Bouby, L. & Ruas, M.-P. Geometric morphometric analysis of grain shape and the identification of two-rowed barley (Hordeum vulgare subsp. distichum L.) in southern France. J. Archaeol. Sci. 41, 568–575 (2014).
Bonhomme, V. et al. Seed morphology uncovers 1500 years of vine agrobiodiversity before the advent of the Champagne wine. Sci. Rep. 11, 2305 (2021).
Thompson, K., Band, S. R. & Hodgson, J. G. Seed size and shape predict persistence in soil. Funct. Ecol. 1, 236–241 (1993).
Schwienbacher, E., Marcante, S. & Erschbamer, B. Alpine species seed longevity in the soil in relation to seed size and shape—A 5-year burial experiment in the Central Alps. Flora Morphol. Distrib. Funct. Ecol. Plants 205, 19–25 (2010).
Zhao, L.-P., Wu, G.-L. & Cheng, J.-M. Seed mass and shape are related to persistence in a sandy soil in northern China. Seed Sci. Res. 21, 47–53 (2011).
Spencer, L. J. & Snow, A. A. Fecundity of transgenic wild-crop hybrids of Cucurbita pepo (Cucurbitaceae): Implications for crop-to-wild gene flow. Heredity 86, 694–702 (2001).
Rue, D. J. Archaic Middle American agriculture and settlement: Recent pollen data from Honduras. J. Field Archaeol. 16, 177–184 (1989).
Rue, D., Webster, D. & Traverse, A. Late Holocene fire and agriculture in the Copan Valley, Honduras. Ancient Mesoam. 13, 267–272 (2002).
Kennett, D. J. et al. High-precision chronology for Central American maize diversification from El Gigante rockshelter, Honduras. PNAS 114, 9026–9031 (2017).
Kistler, L. et al. Archaeological Central American maize genomes suggest ancient gene flow from South America. PNAS. https://doi.org/10.1073/pnas.2015560117 (2020).
Lim, T. K. & Lim, T. K. Cucurbita moschata. In Edible Medicinal and Non-medicinal Plants: Volume 2, Fruits (ed. Lim, T. K.) 266–280 (Springer, 2012).
Vestal, P. A. Cucurbita moschata found in pre-Columbian mounds in Guatemala. Bot. Mus. Leafl. Harvard Univ. 6, 65–69 (1938).
Abràmoff, M. D., Magalhães, P. J. & Ram, S. J. Image processing with ImageJ. Biophoton. Int. 11, 36–42 (2004).
Iwata, H. & Ukai, Y. SHAPE: A computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors. J. Hered. 93, 384–385 (2002).
Acknowledgements
The authors acknowledge the Instituto Hondureño de Antropología e Historia for permission to conduct this research. They thank Brendan Culleton and Laurie Eccles for laboratory assistance and Sarah Chamberlain and the Pennsylvania State University Herbarium for assistance with seed accessions. Methods were carried out in accordance with relevant guidelines and regulations. Financial support for this research came from the National Science Foundation Archaeology Program Grants BCS-100343 to K.H., BCS-1757375 to D.J.K., BCS-1757383 to A.M.V., BCS-1757374 to H.B.T., and The Pennsylvania State University.
Author information
Authors and Affiliations
Contributions
A.I.D., J.M.C., T.M.S., K.H., and D.J.K. designed the research. T.M.S. organized and managed fieldwork and conducted the excavations. A.I.D. identified the plant remains and analyzed the samples. A.D. wrote a draft of the manuscript. A.V., H.B.T., and J.M.C. assisted with the analysis. All authors commented, provided input to the manuscript, and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Domic, A.I., VanDerwarker, A.M., Thakar, H.B. et al. Archaeobotanical evidence supports indigenous cucurbit long-term use in the Mesoamerican Neotropics. Sci Rep 14, 10885 (2024). https://doi.org/10.1038/s41598-024-60723-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-60723-1
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
